
Real and Deepfake Face Recognition: An EEG Study on Cognitive and Emotive Implications

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
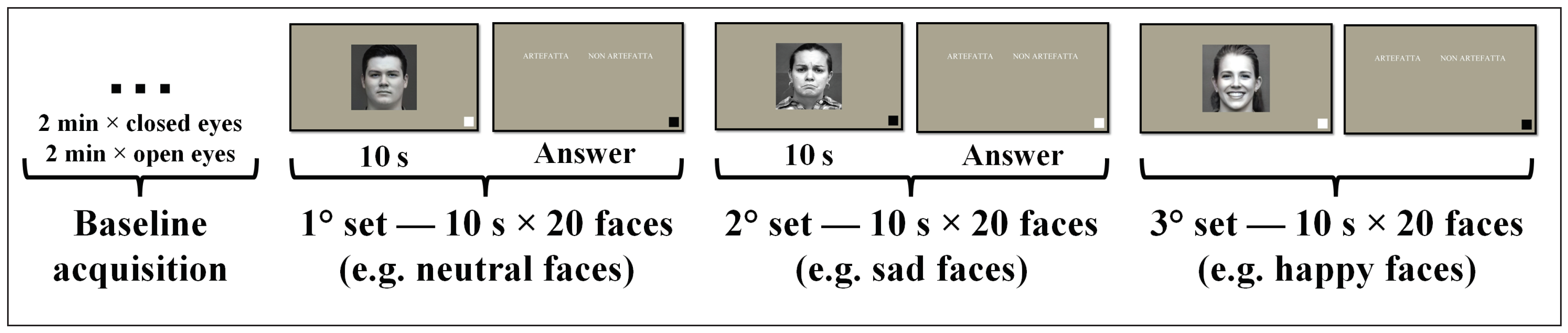
2.2. Experimental Protocol
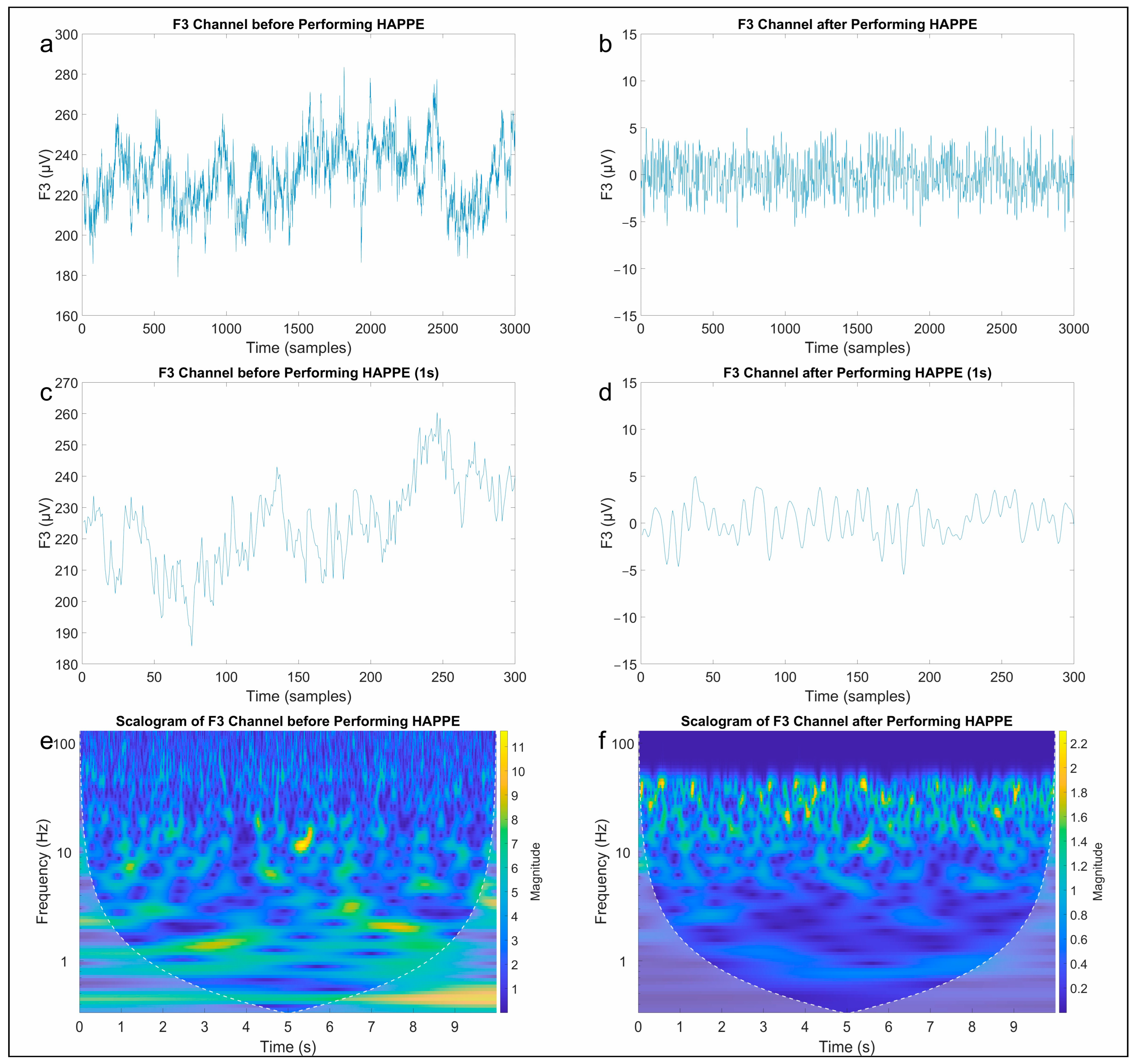
2.3. Signal Processing Chain
- Band-pass filtering (1–45 Hz);
- Channel selection;
- 50 Hz electrical noise removal through Cleanline EEGLAB plugin;
- Crude bad channel detection using spectrum criteria and 3 standard deviations as channel outlier threshold;
- Independent component analysis (ICA) for clustering the data;
- Wavelet-enhanced independent component analysis (W-ICA) for thresholding with a level 5 coiflet wavelet and threshold multiplier 0.75;
- Multiple artifact rejection algorithm (MARA) for independent component rejection if artifact probability is greater than 0.5;
- Segmentation in epochs of 10 s each;
- Interpolation of bad data within segments from good channels only;
- Rejection of bad segments using amplitude-based and joint probability artifact detection;
- Channel interpolation with the spherical method;
- Average re-referencing of channels.
2.4. Statistical Analysis
- Happy—in these epochs, the presented face expressed a “happy” emotion;
- Neutral—in these epochs, the presented face expressed a neutral emotion;
- Sad—in these epochs, the presented face expressed a “sad” emotion;
- tt (i.e., true–true)—in these epochs, the presented face was real, and the subject’s answer was “real”;
- ff (i.e., false–false)—in these epochs, the presented face was synthetic, and the subject’s answer was “synthetic”;
- tf (i.e., true–false)—in these epochs, the presented face was real, and the subject’s answer was “synthetic”;
- ft (i.e., false–true)—in these epochs, the presented face was synthetic, and the subjects’ answer was “real”.
- Imfalse vs. Imtrue: comparison between synthetic and real stimulation.
- -
- HappyF vs. HappyT: comparison between synthetic happy and real happy stimulation.
- -
- NeutralF vs. NeutralT: comparison between synthetic neutral and real neutral stimulation.
- -
- SadF vs. SadT: comparison between synthetic sad and real sad stimulation.
- Happy vs. Neutral: comparison between happy vs. neutral emotions
- -
- Imfalse_H vs. Imfalse_N: comparison between synthetic happy vs. synthetic neutral emotions.
- -
- Imtrue_H vs. Imtrue_N: comparison between real happy vs. real neutral emotions
- Happy vs. Sad: comparison between happy vs. sad emotions
- -
- Imfalse_H vs. Imfalse_S: comparison between synthetic happy vs. synthetic sad emotions
- -
- Imtrue_H vs. Imtrue_S: comparison between real happy vs. real sad emotions
- Neutral vs. Sad: comparison between neutral vs. sad emotions
- -
- Imfalse_N vs. Imfalse_S: comparison between synthetic neutral vs. synthetic sad emotions
- -
- Imtrue_N vs. Imtrue_S: comparison between real neutral vs. real sad emotions
3. Results
3.1. Power Spectrum Analysis
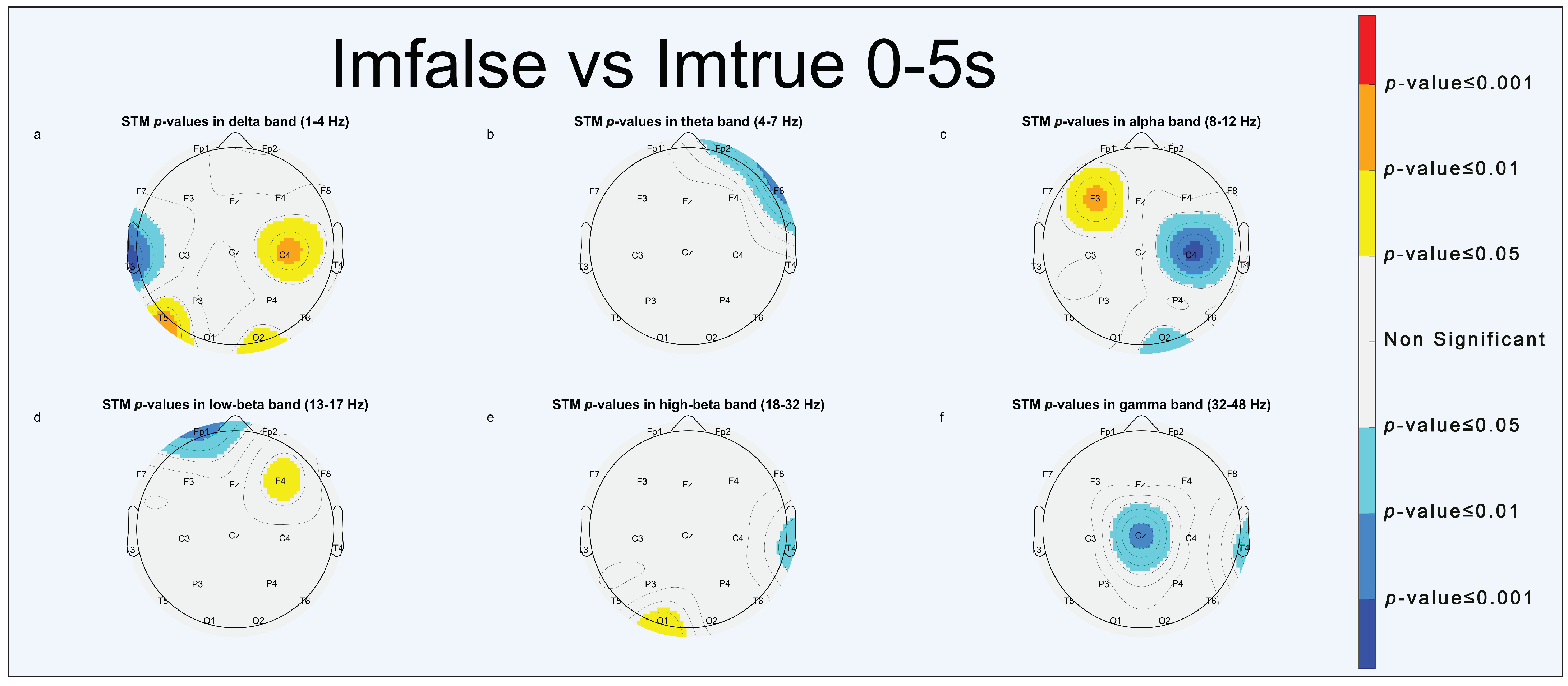
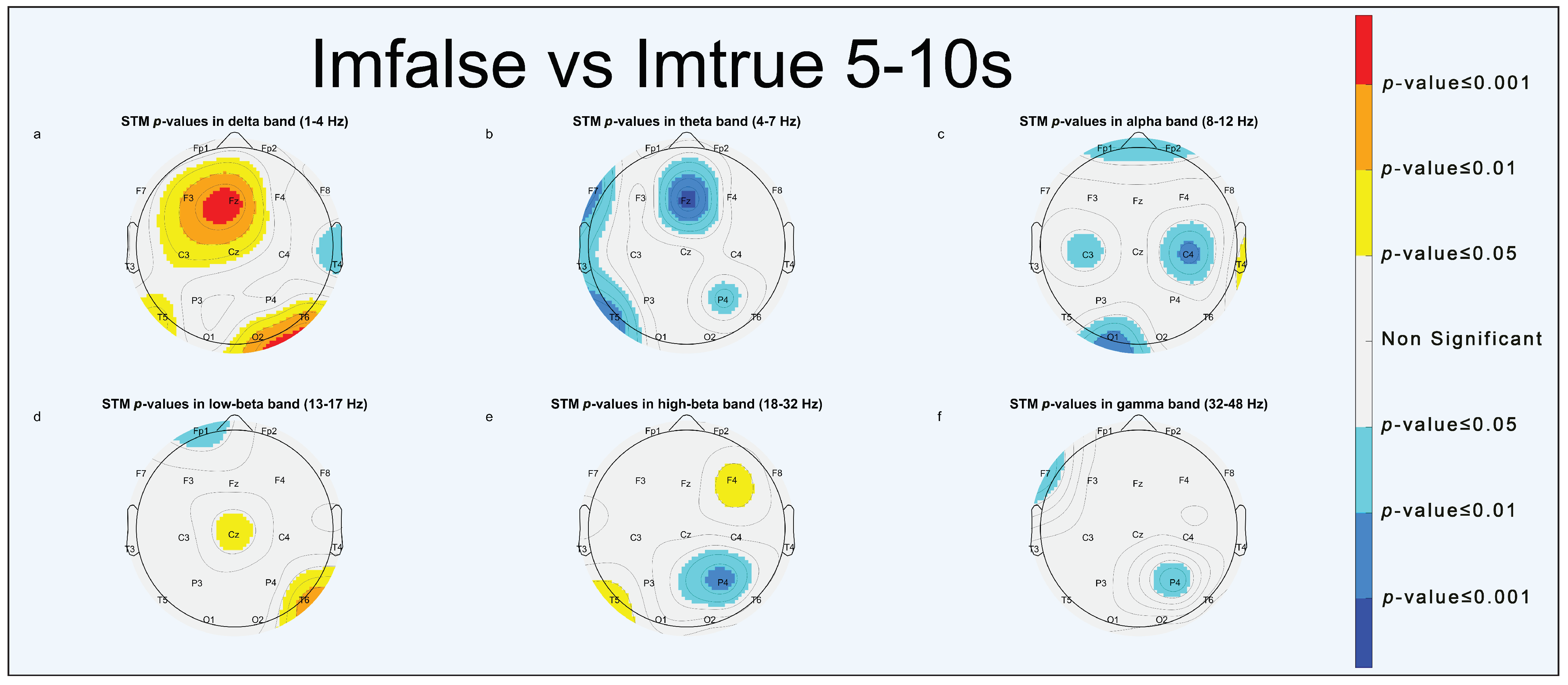
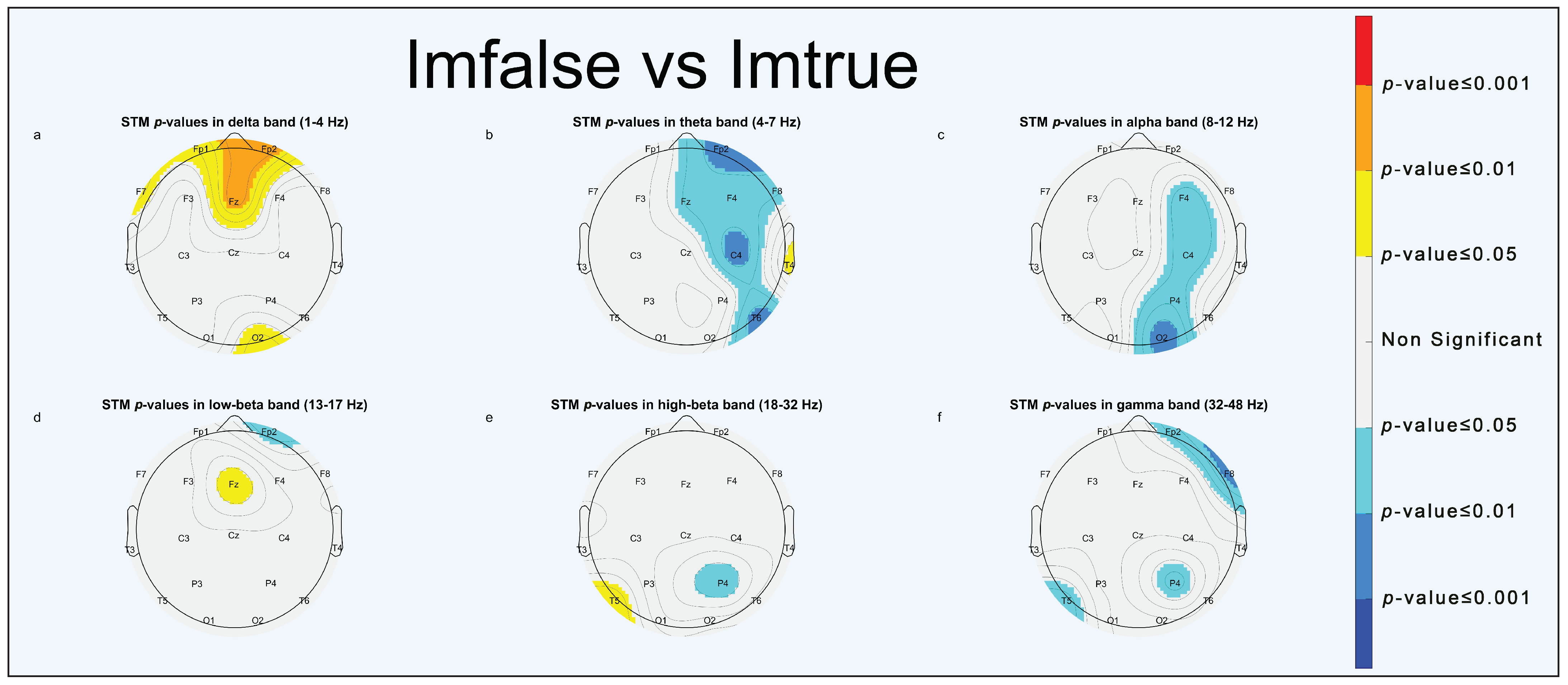
3.1.1. Imfalse vs. Imtrue
3.1.2. Emotional Comparison
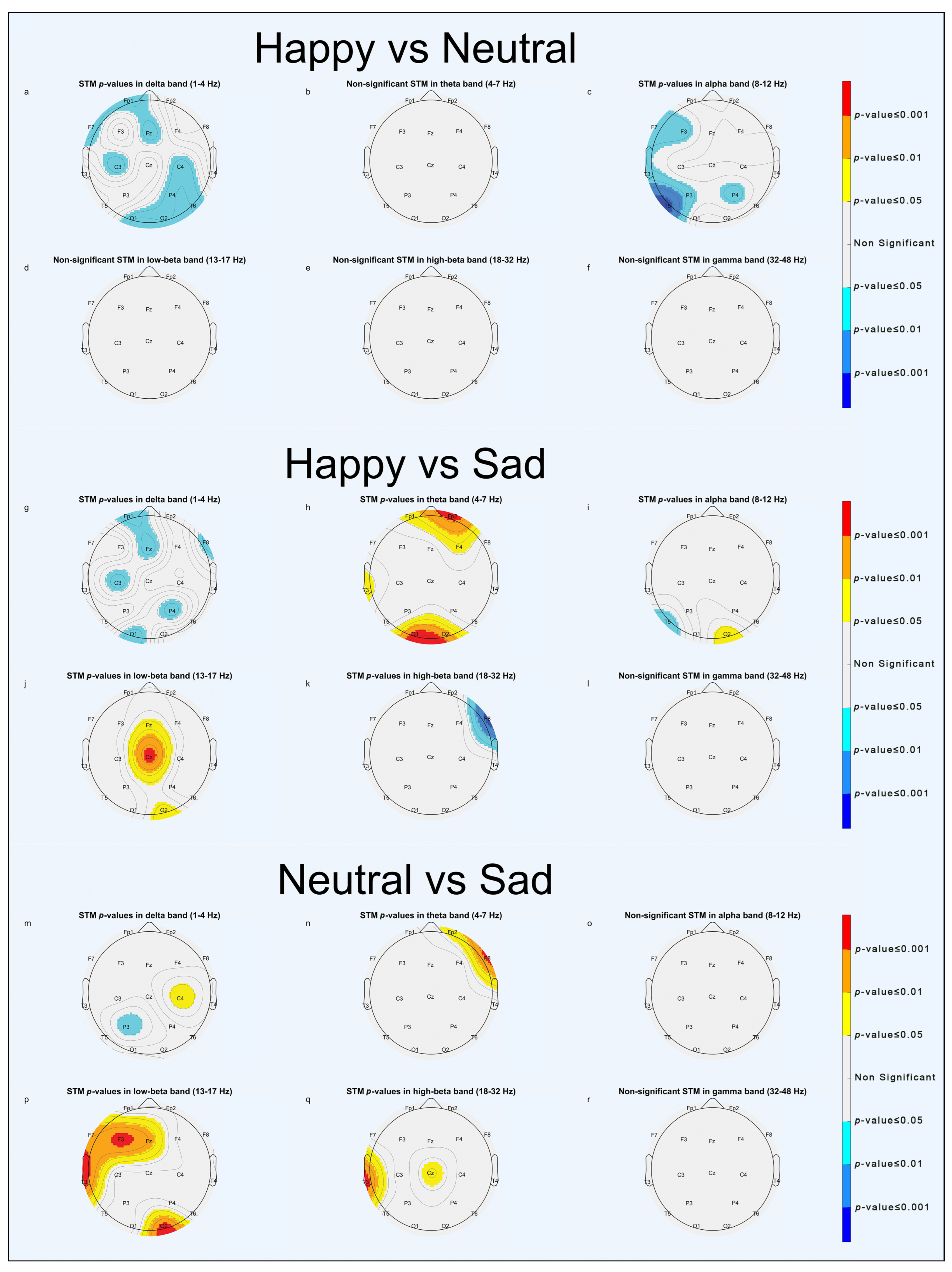
- In the “Happy vs. Neutral” comparison, greater significant activation was in the delta band in the frontal and occipital areas (Figure 7a, Figures S2a and S3a) and in the alpha band in the left temporal and right parietal areas (Figure 7c and Figure S3c) for faces expressing neutral emotions.
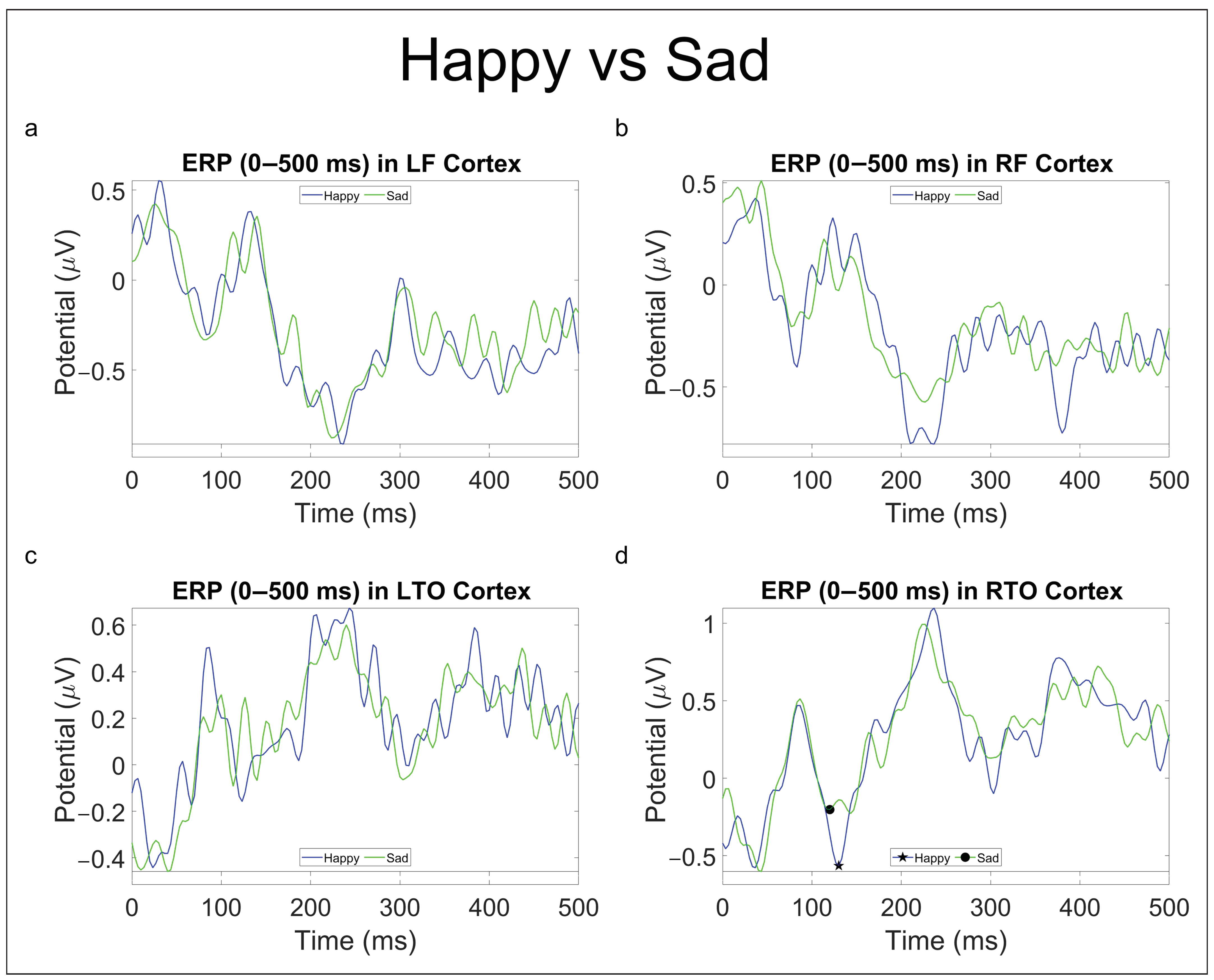
- In the “Happy vs. Sad” comparison, greater significant activities were in the theta band in pre-frontal and left occipital areas (Figure 7h and Figure S3h) and in the low-beta band in the frontal and right occipital areas (Figure 7j and Figure S3j) for faces expressing positive emotions, whereas there was greater significant activation in the alpha band in the left temporal area (Figure 7i and Figure S2i) and in high-beta band in the right frontal area (Figure 7k and Figure S2k) for faces expressing negative emotions. It is worth noting that in the first 5 seconds (Figure S2g–l), faces expressing negative emotions elicited more significant activation, whereas in the last 5 s of the epoch (Figure S3g–l), faces expressing positive emotion were predominant in determining statistical significance.
- In the “Neutral vs. Sad” comparison, significant activations in the low-beta band were found in the right occipital and left temporal areas (Figure 7p, Figures S2p and S3p) for faces expressing neutral emotions.
3.2. ERP Analysis
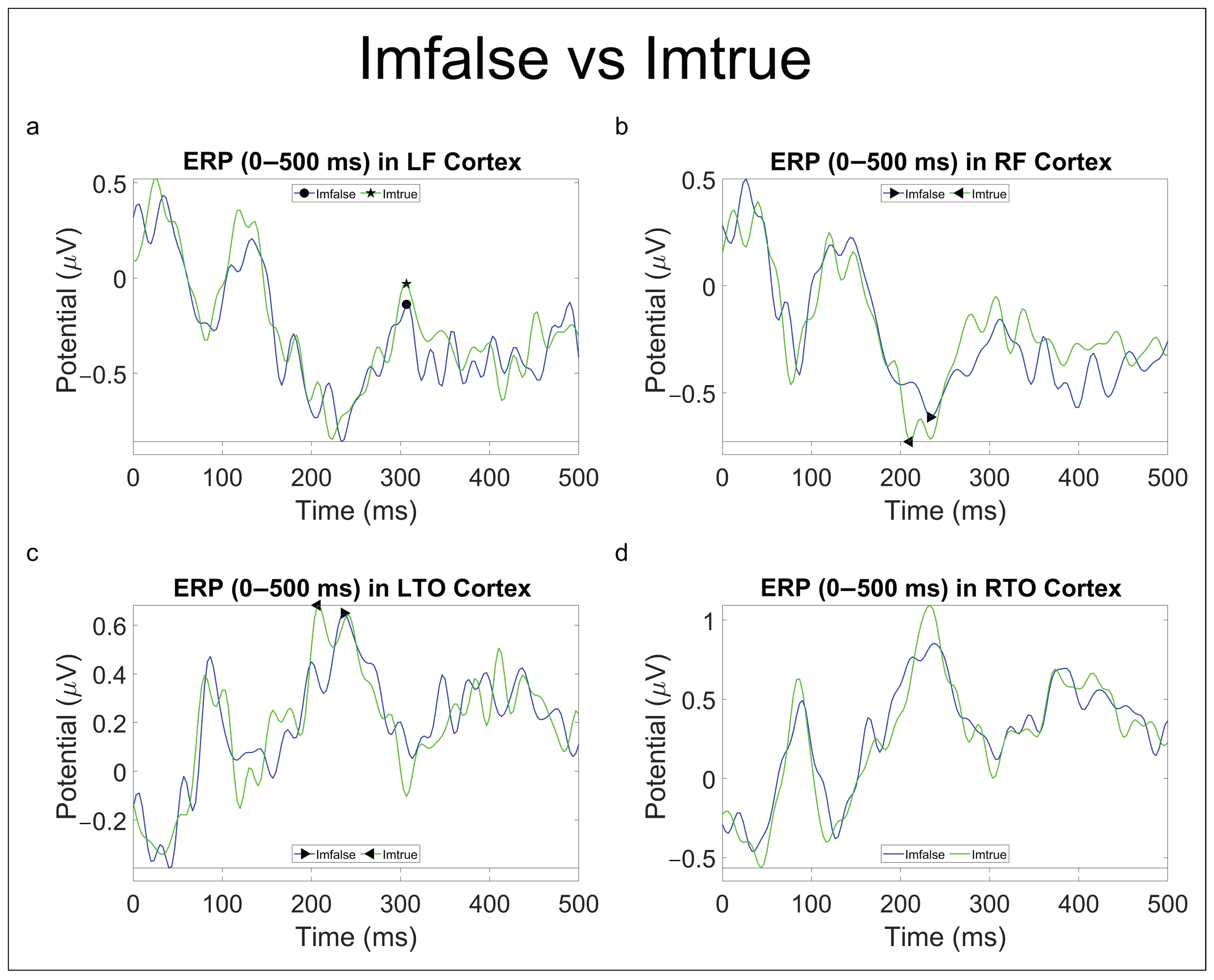
3.2.1. Originality Comparisons
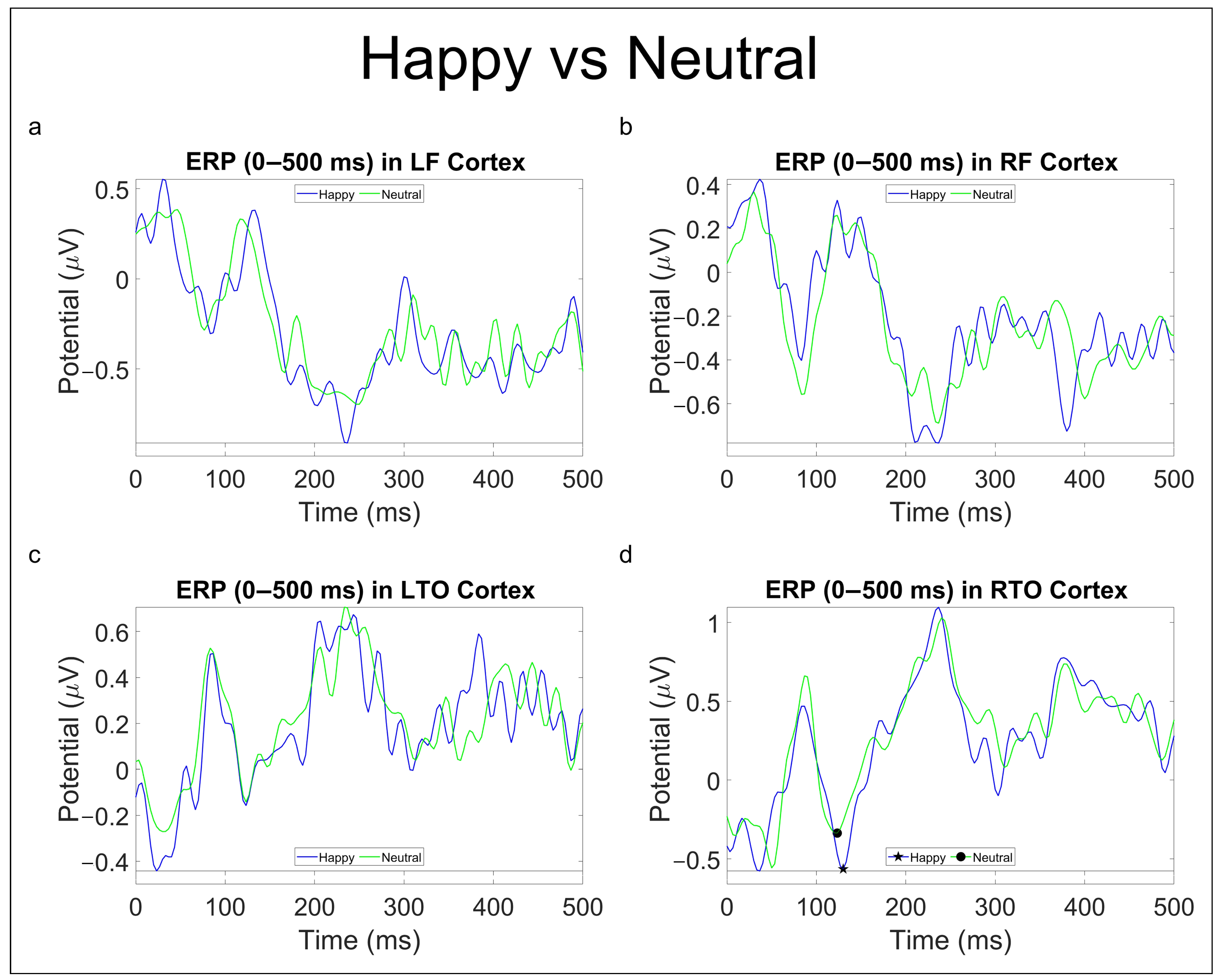
3.2.2. Emotional Comparison
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pascalis, O.; Kelly, D.J. On the development of face processing. Perspect. Psychol. Sci. 2009, 4, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.C.; Raymond, J.E. The role of attention and familiarity in face identification. Percept. Psychophys. 2006, 68, 543–557. [Google Scholar] [CrossRef] [PubMed]
- Kanwisher, N.; Yovel, G. The fusiform face area: A cortical region specialized for the perception of faces. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 2109–2128. [Google Scholar] [CrossRef] [PubMed]
- Babo-Rebelo, M.; Puce, A.; Bullock, D.; Hugueville, L.; Pestilli, F.; Adam, C.; Lehongre, K.; Lambrecq, V.; Dinkelacker, V.; George, N. Visual information routes in the posterior dorsal and ventral face network studied with intracranial neurophysiology and white matter tract endpoints. Cereb. Cortex 2022, 32, 342–366. [Google Scholar] [CrossRef] [PubMed]
- Haxby, J.V.; Hoffman, E.A.; Gobbini, M.I. The distributed human neural system for face perception. Trends Cogn. Sci. 2000, 4, 223–233. [Google Scholar] [CrossRef]
- Adolphs, R. Recognizing emotion from facial expressions: Psychological and neurological mechanisms. Behav. Cogn. Neurosci. Rev. 2002, 1, 21–62. [Google Scholar] [CrossRef]
- Bruce, V.; Young, A. Understanding face recognition. Br. J. Psychol. 1986, 77, 305–327. [Google Scholar] [CrossRef]
- Adolphs, R. Neural systems for recognizing emotion. Curr. Opin. Neurobiol. 2002, 12, 169–177. [Google Scholar] [CrossRef]
- Pessoa, L. On the relationship between emotion and cognition. Nat. Rev. Neurosci. 2008, 9, 148–158. [Google Scholar] [CrossRef]
- Moshel, M.L.; Robinson, A.K.; Carlson, T.A.; Grootswagers, T. Are you for real? Decoding realistic AI-generated faces from neural activity. Vis. Res. 2022, 199, 108079. [Google Scholar] [CrossRef]
- Crookes, K.; Ewing, L.; Gildenhuys, J.-d.; Kloth, N.; Hayward, W.G.; Oxner, M.; Pond, S.; Rhodes, G. How well do computer-generated faces tap face expertise? PLoS ONE 2015, 10, e0141353. [Google Scholar] [CrossRef] [PubMed]
- Makrushin, A.; Siegel, D.; Dittmann, J. Simulation of border control in an ongoing web-based experiment for estimating morphing detection performance of humans. In Proceedings of the 2020 ACM Workshop on Information Hiding and Multimedia Security, Denver, CO, USA, 22–24 June 2020; pp. 91–96. [Google Scholar]
- Farid, H.; Bravo, M.J. Perceptual discrimination of computer generated and photographic faces. Digit. Investig. 2012, 8, 226–235. [Google Scholar] [CrossRef]
- Holmes, O.; Banks, M.S.; Farid, H. Assessing and improving the identification of computer-generated portraits. ACM Trans. Appl. Percept. (TAP) 2016, 13, 1–12. [Google Scholar] [CrossRef]
- Mader, B.; Banks, M.S.; Farid, H. Identifying computer-generated portraits: The importance of training and incentives. Perception 2017, 46, 1062–1076. [Google Scholar] [CrossRef]
- Korshunov, P.; Marcel, S. Deepfake detection: Humans vs. machines. arXiv 2020, arXiv:2009.03155. [Google Scholar]
- Tauscher, J.P.; Castillo, S.; Bosse, S.; Magnor, M. EEG-based Analysis of the Impact of Familiarity in the Perception of Deepfake Videos. In Proceedings of the 2021 IEEE International Conference on Image Processing (ICIP), Anchorage, AK, USA, 19–22 September 2021; IEEE: Piscataway, NJ, USA, 2021; pp. 160–164. [Google Scholar]
- Acunzo, D.; MacKenzie, G.; van Rossum, M.C. Spatial attention affects the early processing of neutral versus fearful faces when they are task-irrelevant: A classifier study of the EEG C1 component. Cogn. Affect. Behav. Neurosci. 2019, 19, 123–137. [Google Scholar] [CrossRef]
- Leppänen, J.M.; Moulson, M.C.; Vogel-Farley, V.K.; Nelson, C.A. An ERP study of emotional face processing in the adult and infant brain. Child Dev. 2007, 78, 232–245. [Google Scholar] [CrossRef]
- Lucey, P.; Cohn, J.F.; Kanade, T.; Saragih, J.; Ambadar, Z.; Matthews, I. The extended cohn-kanade dataset (ck+): A complete dataset for action unit and emotion-specified expression. In Proceedings of the 2010 IEEE Computer Society Conference on Computer Vision and Pattern Recognition-Workshops, San Francisco, CA, USA, 13–18 June 2010; IEEE: Piscataway, NJ, USA, 2010; pp. 94–101. [Google Scholar]
- Unlimited Free Face Mix AI Generator-Playform-AI Art Generative Platform for Artists and Creative People. Free, Unlimited, Easy. Playform. Available online: https://playform.io/facemix (accessed on 1 February 2023).
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Gabard-Durnam, L.J.; Mendez Leal, A.S.; Wilkinson, C.L.; Levin, A.R. The Harvard Automated Processing Pipeline for Electroencephalography (HAPPE): Standardized processing software for developmental and high-artifact data. Front. Neurosci. 2018, 12, 97. [Google Scholar] [CrossRef]
- Moretti, D.V.; Babiloni, C.; Binetti, G.; Cassetta, E.; Dal Forno, G.; Ferreric, F.; Ferri, R.; Lanuzza, B.; Miniussi, C.; Nobili, F.; et al. Individual analysis of EEG frequency and band power in mild Alzheimer’s disease. Clin. Neurophysiol. 2004, 115, 299–308. [Google Scholar] [CrossRef]
- Delorme, A. EEG is better left alone. Sci. Rep. 2023, 13, 2372. [Google Scholar] [PubMed]
- Liu, Z.; Du, W.; Sun, Z.; Hou, G.; Wang, Z. Neural Processing Differences of Facial Emotions Between Human and Vehicles: Evidence From an Event-Related Potential Study. Front. Psychol. 2022, 13, 876252. [Google Scholar] [PubMed]
- Matyjek, M.; Kroczek, B.; Senderecka, M. Socially induced negative affective knowledge modulates early face perception but not gaze cueing of attention. Psychophysiology 2021, 58, e13876. [Google Scholar] [PubMed]
- Morgan, H.M.; Klein, C.; Boehm, S.G.; Shapiro, K.L.; Linden, D.E. Working memory load for faces modulates P300, N170, and N250r. J. Cogn. Neurosci. 2008, 20, 989–1002. [Google Scholar]
- Schweinberger, S.R.; Pickering, E.C.; Jentzsch, I.; Burton, A.M.; Kaufmann, J.M. Event-related brain potential evidence for a response of inferior temporal cortex to familiar face repetitions. Cogn. Brain Res. 2002, 14, 398–409. [Google Scholar]
- Faes, L.; Porta, A.; Nollo, G. Surrogate data approaches to assess the significance of directed coherence: Application to EEG activity propagation. In Proceedings of the 2009 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Minneapolis, MN, USA, 3–6 September 2009; IEEE: Piscataway, NJ, USA, 2009; pp. 6280–6283. [Google Scholar]
- Barton, J.J. Face processing in the temporal lobe. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2022; Volume 187, pp. 191–210. [Google Scholar]
- Collins, A.; Koechlin, E. Reasoning, learning, and creativity: Frontal lobe function and human decision-making. PLoS Biol. 2012, 10, e1001293. [Google Scholar]
- Samal, A.; Iyengar, P.A. Automatic recognition and analysis of human faces and facial expressions: A survey. Pattern Recognit. 1992, 25, 65–77. [Google Scholar]
- Wilmer, J.B.; Germine, L.; Chabris, C.F.; Chatterjee, G.; Williams, M.; Loken, E.; Nakayama, K.; Duchaine, B. Human face recognition ability is specific and highly heritable. Proc. Natl. Acad. Sci. USA 2010, 107, 5238–5241. [Google Scholar]
- Montagrin, A.; Sterpenich, V.; Brosch, T.; Grandjean, D.; Armony, J.; Ceravolo, L.; Sander, D. Goal-relevant situations facilitate memory of neutral faces. Cogn. Affect. Behav. Neurosci. 2018, 18, 1269–1282. [Google Scholar]
- Jiang, Y.; Zhang, H.; Yu, S. Changes in delta and theta oscillations in the brain indicate dynamic switching of attention between internal and external processing. In Proceedings of the 4th International Conference on Biometric Engineering and Applications, Taiyuan, China, 25–27 May 2021; pp. 25–31. [Google Scholar]
- Deffke, I.; Sander, T.; Heidenreich, J.; Sommer, W.; Curio, G.; Trahms, L.; Lueschow, A. MEG/EEG sources of the 170-ms response to faces are co-localized in the fusiform gyrus. Neuroimage 2007, 35, 1495–1501. [Google Scholar]
- Canales-Johnson, A.; Lanfranco, R.C.; Morales, J.P.; Martínez-Pernía, D.; Valdés, J.; Ezquerro-Nassar, A.; Rivera-Rei, Á.; Ibanez, A.; Chennu, S.; Bekinschtein, T.A.; et al. In your phase: Neural phase synchronisation underlies visual imagery of faces. Sci. Rep. 2021, 11, 2401. [Google Scholar] [PubMed]
- Cavanagh, J.F.; Frank, M.J. Frontal theta as a mechanism for cognitive control. Trends Cogn. Sci. 2014, 18, 414–421. [Google Scholar] [PubMed]
- Sergent, J.; Ohta, S.; Macdonald, B. Functional neuroanatomy of face and object processing: A positron emission tomography study. Brain 1992, 115, 15–36. [Google Scholar]
- Brady, N.; Campbell, M.; Flaherty, M. Perceptual asymmetries are preserved in memory for highly familiar faces of self and friend. Brain Cogn. 2005, 58, 334–342. [Google Scholar]
- Knyazev, G.G.; Barchard, K.A.; Razumnikova, O.M.; Mitrofanova, L.G. The relationship of positive and negative expressiveness to the processing of emotion information. Scand. J. Psychol. 2012, 53, 206–215. [Google Scholar] [PubMed]
- Luo, W.; Feng, W.; He, W.; Wang, N.Y.; Luo, Y.J. Three stages of facial expression processing: ERP study with rapid serial visual presentation. Neuroimage 2010, 49, 1857–1867. [Google Scholar] [PubMed]
- Debruille, J.B.; Guillem, F.; Renault, B. ERPs and chronometry of face recognition: Following-up Seeck: et al.: and George: et al. Neuroreport 1998, 9, 3349–3353. [Google Scholar]
- Herrmann, C.; Grigutsch, M.; Busch, N.; Handy, T.C. Event-Related Potentials: A Methods Handbook; Handy, T.C., Ed.; MIT Press: Cambridge, MA, USA, 2005; pp. 229–259. [Google Scholar]
- Dering, C.; Hemmelmann, C.; Pugh, E.; Ziegler, A. Statistical analysis of rare sequence variants: An overview of collapsing methods. Genet. Epidemiol. 2011, 35, S12–S17. [Google Scholar]
- Sommer, W.; Stapor, K.; Kończak, G.; Kotowski, K.; Fabian, P.; Ochab, J.; Bereś, A.; Ślusarczyk, G. The N250 event-related potential as an index of face familiarity: A replication study. R. Soc. Open Sci. 2021, 8, 202356. [Google Scholar]
- Ashley, V.; Vuilleumier, P.; Swick, D. Time course and specificity of event-related potentials to emotional expressions. Neuroreport 2004, 15, 211–216. [Google Scholar]
- Brashier, N.M.; Marsh, E.J. Judging truth. Annu. Rev. Psychol. 2020, 71, 499–515. [Google Scholar] [PubMed]
- Forgas, J.P.; East, R. On being happy and gullible: Mood effects on skepticism and the detection of deception. J. Exp. Soc. Psychol. 2008, 44, 1362–1367. [Google Scholar]
- Bigdely-Shamlo, N.; Mullen, T.; Kothe, C.; Su, K.M.; Robbins, K.A. The PREP pipeline: Standardized preprocessing for large-scale EEG analysis. Front. Neuroinform. 2015, 9, 16. [Google Scholar] [CrossRef]
- da Cruz, J.R.; Chicherov, V.; Herzog, M.H.; Figueiredo, P. An automatic pre-processing pipeline for EEG analysis (APP) based on robust statistics. Clin. Neurophysiol. 2018, 129, 1427–1437. [Google Scholar] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Epochs Label | tt | ff | tf | ft | Total | Percentage |
|---|---|---|---|---|---|---|
| happy | 173 | 172 | 57 | 58 | 460 | 33.3% |
| neutral | 180 | 181 | 50 | 49 | 460 | 33.3% |
| sad | 171 | 172 | 59 | 58 | 460 | 33.3% |
| total | 524 | 525 | 166 | 165 | 1380 | 100% |
| percentage | 38% | 38% | 12% | 12% | 100% |
| Epochs Label | tt | ff | tf | ft | Total | Percentage |
|---|---|---|---|---|---|---|
| happy | 148 | 151 | 51 | 45 | 395 | 32.3% |
| neutral | 159 | 162 | 45 | 43 | 409 | 33.4% |
| sad | 158 | 153 | 56 | 53 | 420 | 34.3% |
| total | 465 | 466 | 152 | 141 | 1224 | 100% |
| percentage | 38% | 38.1% | 12.4% | 11.5% | 100% |
| Comparisons | LF | RF | LTO | RTO |
|---|---|---|---|---|
| Imfalse vs. Imtrue | P300 (T) ↑ | N250 (T) ← | N250 (T) ← | - |
| HappyF vs. HappyT | N100 (F) ← | - | P300 (T) ↑ | - |
| NeutralF vs. NeutralT | - | - | - | N170 (F) ← |
| SadF vs. SadT | N250 (T) ← | - | P100 (F) ↑ | - |
| Comparisons | LF | RTO |
|---|---|---|
| Happy vs. Neutral | - | P100 (H) ↑ |
| Happy vs. Sad | - | P100 (H) ↑ |
| Imfalse_N vs. Imfalse_S | N100 (N) ↑ | - |
| Imtrue_N vs. Imtrue_S | N170 (S) ↑ | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarchi, P.; Lanini, M.C.; Frassineti, L.; Lanatà, A. Real and Deepfake Face Recognition: An EEG Study on Cognitive and Emotive Implications. Brain Sci. 2023, 13, 1233. https://doi.org/10.3390/brainsci13091233
Tarchi P, Lanini MC, Frassineti L, Lanatà A. Real and Deepfake Face Recognition: An EEG Study on Cognitive and Emotive Implications. Brain Sciences. 2023; 13(9):1233. https://doi.org/10.3390/brainsci13091233
Chicago/Turabian StyleTarchi, Pietro, Maria Chiara Lanini, Lorenzo Frassineti, and Antonio Lanatà. 2023. "Real and Deepfake Face Recognition: An EEG Study on Cognitive and Emotive Implications" Brain Sciences 13, no. 9: 1233. https://doi.org/10.3390/brainsci13091233