
The Role of Serotonin in Fear Learning and Memory: A Systematic Review of Human Studies

 ,
,  ,
,  and
and
Abstract
:1. Introduction
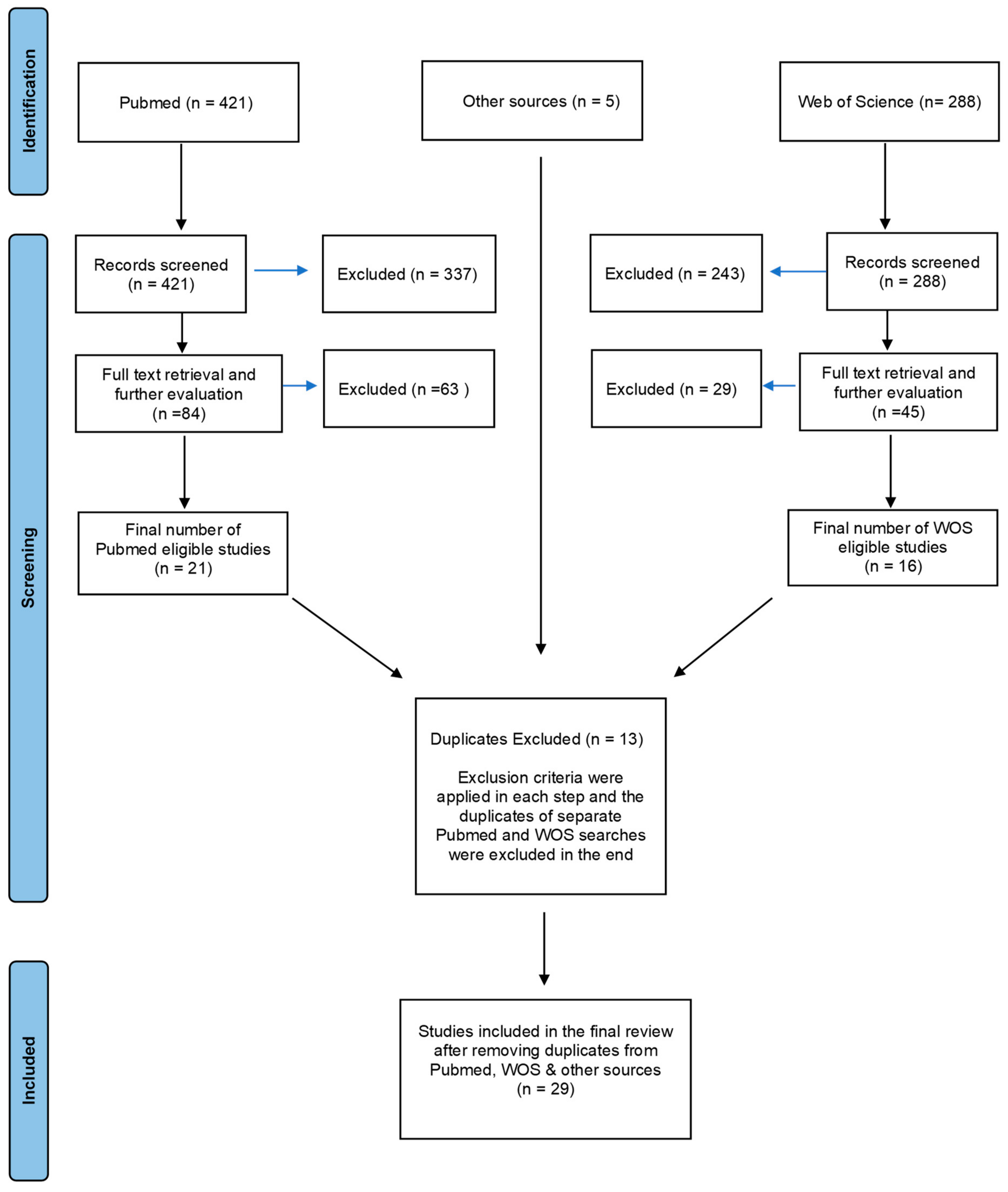
2. Methods
3. Results
3.1. Studies on 5-HT Modulators or Tryptophan Depletion without Genotyping
3.1.1. Effects of SSRIs and Other 5-HT Modulators
3.1.2. Effects of Acute Tryptophan Depletion (ATD) on 5-HT Levels
3.2. Investigations of Genetic Modulation of 5-HT in Fear Learning and Extinction
4. Discussion
4.1. SSRI Studies
4.2. Genetic Studies
4.3. Sex, Serotonin, and FC
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Erland, L.A.E.; Turi, C.E.; Saxena, P.K. Serotonin in plants: Origin, functions, and implications. In Serotonin the Mediator That Spans Evolution; Pilowsky, P.M., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 23–46. [Google Scholar] [CrossRef]
- Azmitia, E.C. Modern views on an ancient chemical: Serotonin effects on cell proliferation, maturation, and apoptosis. Brain Res. Bull. 2001, 56, 413–424. [Google Scholar] [CrossRef]
- Bacqué-cazenave, J.; Bharatiya, R.; Barrière, G.; Delbecque, J.-P.P.; Bouguiyoud, N.; Di Giovanni, G.; Cattaert, D.; De Deurwaerdère, P. Serotonin in animal cognition and behavior. Int. J. Mol. Sci. 2020, 21, 1649. [Google Scholar] [CrossRef] [Green Version]
- Moses, L.; Mohammad-Zadeh, L.F.; Moses, L.; Gwaltney-Brant, S.M. Serotonin: A review. J. Vet. Pharmacol. Ther. 2008, 31, 187–199. [Google Scholar] [CrossRef]
- Deakin, J.F.W. The origins of ‘5-HT and mechanisms of defence’ by Deakin and Graeff: A personal perspective. J. Psychopharmacol. 2013, 27, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, K.W.; Miyazaki, K.; Doya, K. Activation of dorsal raphe serotonin neurons is necessary for waiting for delayed rewards. J. Neurosci. 2012, 32, 10451–10457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branchi, I. The double edged sword of neural plasticity: Increasing serotonin levels leads to both greater vulnerability to depression and improved capacity to recover. Psychoneuroendocrinology 2011, 36, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Puglisi-Allegra, S.; Andolina, D. Serotonin and stress coping. Behav. Brain Res. 2015, 277, 58–67. [Google Scholar] [CrossRef]
- Edwards, D.H.; Spitzer, N. Social Dominance and Serotonin Receptor Genes in Crayfish. Curr. Top. Dev. Biol. 2006, 74, 177–199. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, P.A.; Rillich, J. The decision to fight or flee—Insights into underlying mechanism in crickets. Front. Neurosci. 2012, 6, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miczek, K.A.; Fish, E.W.; De Bold, J.F.; De Almeida, R.M. Social and neural determinants of aggressive behavior: Pharmacotherapeutic targets at serotonin, dopamine and γ-aminobutyric acid systems. Psychopharmacology 2002, 163, 434–458. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.R.; Fricke, R.A.; Edwards, D.H. The effect of social experience on serotonergic modulation of the escape circuit of crayfish. Science 1996, 271, 366–369. [Google Scholar] [CrossRef] [Green Version]
- Huber, R.; Smith, K.; Delago, A.; Isaksson, K.; Kravitz, E.A. Serotonin and aggressive motivation in crustaceans: Altering the decision to retreat. Proc. Natl. Acad. Sci. USA 1997, 94, 5939–5942. [Google Scholar] [CrossRef] [PubMed]
- Bacqué-Cazenave, J.; Cattaert, D.; Delbecque, J.P.; Fossat, P. Alteration of size perception: Serotonin has opposite effects on the aggressiveness of crayfish confronting either a smaller or a larger rival. J. Exp. Biol. 2018, 221, jeb177840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeDoux, J. The emotional brain, fear, and the amygdala. Cell. Mol. Neurobiol. 2003, 23, 727–738. [Google Scholar] [CrossRef] [PubMed]
- LeDoux, J.E. Coming to terms with fear. Proc. Natl. Acad. Sci. USA 2014, 111, 2871–2878. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Nazzi, C.; Thayer, J.F. Fear-induced bradycardia in mental disorders: Foundations, current advances, future perspectives. Neurosci. Biobehav. Rev. 2023, 149, 105163. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S. Neurobiological advances of learned fear in humans. Adv. Clin. Exp. Med. 2022, 31, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Milad, M.R.; Quirk, G.J. Fear extinction as a model for translational neuroscience: Ten years of progress. Annu. Rev. Psychol. 2012, 63, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Vervliet, B.; Baeyens, F.; Van den Bergh, O.; Hermans, D. Extinction, generalization, and return of fear: A critical review of renewal research in humans. Biol. Psychol. 2013, 92, 51–58. [Google Scholar] [CrossRef]
- Davis, L.L.; Suris, A.; Lambert, M.T.; Heimberg, C.; Petty, F. Post-traumatic stress disorder and serotonin: New directions for research and treatment. J. Psychiatry Neurosci. 1997, 22, 318–326. [Google Scholar]
- Indovina, I.; Robbins, T.W.; Núñez-Elizalde, A.O.; Dunn, B.D.; Bishop, S.J. Fear-Conditioning Mechanisms Associated with Trait Vulnerability to Anxiety in Humans. Neuron 2011, 69, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Parsons, R.G.; Ressler, K.J. Implications of memory modulation for post-traumatic stress and fear disorders. Nat. Neurosci. 2013, 16, 146–153. [Google Scholar] [CrossRef] [Green Version]
- Tortella-Feliu, M.; Fullana, M.A.; Pérez-Vigil, A.; Torres, X.; Chamorro, J.; Littarelli, S.A.; Solanes, A.; Ramella-Cravaro, V.; Vilar, A.; González-Parra, J.A.; et al. Risk factors for posttraumatic stress disorder: An umbrella review of systematic reviews and meta-analyses. Neurosci. Biobehav. Rev. 2019, 107, 154–165. [Google Scholar] [CrossRef]
- Vicario, C.M.; Felmingham, K.L. Slower time estimation in post-traumatic stress disorder. Sci. Rep. 2018, 8, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicario, C.M.; Martino, G.; Lucifora, C.; Felmingham, K. Preliminary evidence on the neural correlates of timing deficit in post-traumatic stress disorder. Eur. J. Psychotraumatol. 2022, 13, 2008151. [Google Scholar] [CrossRef]
- Jafari, E.; Alizadehgoradel, J.; Koluri, F.P.; Nikoozadehkordmirza, E.; Refahi, M.; Taherifard, M.; Nejati, V.; Hallajian, A.-H.; Ghanavati, E.; Vicario, C.M. Intensified electrical stimulation targeting lateral and medial prefrontal cortices for the treatment of social anxiety disorder: A randomized, double-blind, parallel-group, dose-comparison study. Brain Stimul. 2021, 14, 974–986. [Google Scholar] [CrossRef] [PubMed]
- Vicario, C.M.; Scavone, V.; Lucifora, C.; Falzone, A.; Pioggia, G.; Gangemi, S.; Craparo, G.; Martino, G. Evidence of abnormal scalar timing property in alexithymia. PLoS ONE 2023, 18, e0278881. [Google Scholar] [CrossRef]
- La Rosa, V.L.; Gori, A.; Faraci, P.; Vicario, C.M.; Craparo, G. Traumatic distress, alexithymia, dissociation, and risk of addiction during the first wave of COVID-19 in Italy: Results from a cross-sectional online survey on a non-clinical adult sample. Int. J. Ment. Health Addict. 2022, 20, 3128–3144. [Google Scholar] [CrossRef]
- Asberg, M.; Thoren, P.; Traskman, L.; Bertilsson, L.; Ringberger, V. “Serotonin depression”—A biochemical subgroup within the affective disorders? Science 1976, 191, 478–480. [Google Scholar] [CrossRef]
- Hartley, C.A.; McKenna, M.C.; Salman, R.; Holmes, A.; Casey, B.J.; Phelps, E.A.; Glatt, C.E. Serotonin transporter polyadenylation polymorphism modulates the retention of fear extinction memory. Proc. Natl. Acad. Sci. USA 2012, 109, 5493–5498. [Google Scholar] [CrossRef] [PubMed]
- Kahn, R.S.; Van Praag, H.M.; Wetzler, S.; Asnis, G.M.; Barr, G. Serotonin and anxiety revisited. Biol. Psychiatry 1988, 23, 189–208. [Google Scholar] [CrossRef] [PubMed]
- Castrén, E. Is mood chemistry. Nat. Rev. Neurosci. 2005, 6, 241–246. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Battaglia, S.; Garofalo, S.; Tortora, F.; Avenanti, A.; di Pellegrino, G. State-dependent TMS over prefrontal cortex disrupts fear-memory reconsolidation and prevents the return of fear. Curr. Biol. 2020, 30, 3672–3679. [Google Scholar] [CrossRef] [PubMed]
- Borgomaneri, S.; Battaglia, S.; Avenanti, A.; di Pellegrino, G. Don’t Hurt Me No More: State-dependent Transcranial Magnetic Stimulation for the treatment of specific phobia. J. Affect. Disord. 2021, 286, 78–79. [Google Scholar] [CrossRef] [PubMed]
- Borgomaneri, S.; Battaglia, S.; Sciamanna, G.; Tortora, F.; Laricchiuta, D. Memories are not written in stone: Re-writing fear memories by means of non-invasive brain stimulation and optogenetic manipulations. Neurosci. Biobehav. Rev. 2021, 127, 334–352. [Google Scholar] [CrossRef]
- Pittig, A.; Treanor, M.; LeBeau, R.T.; Craske, M.G. The role of associative fear and avoidance learning in anxiety disorders: Gaps and directions for future research. Neurosci. Biobehav. Rev. 2018, 88, 117–140. [Google Scholar] [CrossRef]
- Lucifora, C.; Grasso, G.M.; Nitsche, M.A.; D’Italia, G.; Sortino, M.; Salehinejad, M.A.; Falzone, A.; Avenanti, A.; Vicario, C.M. Enhanced fear acquisition in individuals with evening chronotype. A virtual reality fear conditioning/extinction study. J. Affect. Disord. 2022, 311, 344–352. [Google Scholar] [CrossRef]
- Hartley, C.A.; Phelps, E.A. Changing fear: The neurocircuitry of emotion regulation. Neuropsychopharmacology 2010, 35, 136–146. [Google Scholar] [CrossRef] [Green Version]
- Bunney, W.E.; Davis, J.M. Norepinephrine in depressive reactions: A review. Arch. Gen. Psychiatry 1965, 13, 483–494. [Google Scholar] [CrossRef]
- Coppen, A. The biochemistry of affective disorders. Br. J. Psychiatry 1967, 113, 1237–1264. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.-L.; Licinio, J. From monoamines to genomic targets: A paradigm shift for drug discovery in depression. Nat. Rev. Drug Discov. 2004, 3, 136–151. [Google Scholar] [CrossRef] [PubMed]
- Bertini, C.; Làdavas, E. Fear-related signals are prioritised in visual, somatosensory and spatial systems. Neuropsychologia 2021, 150, 107698. [Google Scholar] [CrossRef] [PubMed]
- Borgomaneri, S.; Gazzola, V.; Avenanti, A. Transcranial magnetic stimulation reveals two functionally distinct stages of motor cortex involvement during perception of emotional body language. Brain Struct. Funct. 2015, 220, 2765–2781. [Google Scholar] [CrossRef] [Green Version]
- Borgomaneri, S.; Vitale, F.; Battaglia, S.; Avenanti, A. Early right motor cortex response to happy and fearful facial expressions: A TMS motor-evoked potential study. Brain Sci. 2021, 11, 1203. [Google Scholar] [CrossRef]
- Borhani, K.; Ladavas, E.; Maier, M.E.; Avenanti, A.; Bertini, C. Emotional and movement-related body postures modulate visual processing. Soc. Cogn. Affect. Neurosci. 2015, 10, 1092–1101. [Google Scholar] [CrossRef] [Green Version]
- Tamietto, M.; De Gelder, B. Neural bases of the non-conscious perception of emotional signals. Nat. Rev. Neurosci. 2010, 11, 697–709. [Google Scholar] [CrossRef]
- Lonsdorf, T.B.; Merz, C.J. More than just noise: Inter-individual differences in fear acquisition, extinction and return of fear in humans—Biological, experiential, temperamental factors, and methodological pitfalls. Neurosci. Biobehav. Rev. 2017, 80, 703–728. [Google Scholar] [CrossRef]
- Lonsdorf, T.B.; Merz, C.J.; Fullana, M.A. Fear extinction retention: Is it what we think it is? Biol. Psychiatry 2019, 85, 1074–1082. [Google Scholar] [CrossRef]
- Battaglia, S.; Harrison, B.J.; Fullana, M.A. Does the human ventromedial prefrontal cortex support fear learning, fear extinction or both? A commentary on subregional contributions. Mol. Psychiatry 2022, 27, 784–786. [Google Scholar] [CrossRef]
- Vicario, C.M. Uncovering the neurochemistry of reward and aversiveness. Front. Mol. Neurosci. 2013, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Vicario, C.M. Aberrant disgust response and immune reactivity in cocaine-dependent men might uncover deranged serotoninergic activity. Front. Mol. Neurosci. 2014, 7, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicario, C.M.; Rafal, R.D.; Martino, D.; Avenanti, A. Core, social and moral disgust are bounded: A review on behavioral and neural bases of repugnance in clinical disorders. Neurosci. Biobehav. Rev. 2017, 80, 185–200. [Google Scholar] [CrossRef]
- Bauer, E.P. Serotonin in fear conditioning processes. Behav. Brain Res. 2015, 277, 68–77. [Google Scholar] [CrossRef]
- Cools, R.; Roberts, A.C.; Robbins, T.W. Serotoninergic regulation of emotional and behavioural control processes. Trends Cogn. Sci. 2008, 12, 31–40. [Google Scholar] [CrossRef] [PubMed]
- LeDoux, J.E. Emotion circuits in the brain. Annu. Rev. Neurosci. 2000, 23, 155–184. [Google Scholar] [CrossRef]
- Lonsdorf, T.B.; Menz, M.M.; Andreatta, M.; Fullana, M.A.; Golkar, A.; Haaker, J.; Heitland, I.; Hermann, A.; Kuhn, M.; Kruse, O.; et al. Don’t fear ‘fear conditioning’: Methodological considerations for the design and analysis of studies on human fear acquisition, extinction, and return of fear. Neurosci. Biobehav. Rev. 2017, 77, 247–285. [Google Scholar] [CrossRef]
- Duits, P.; Cath, D.C.; Lissek, S.; Hox, J.J.; Hamm, A.O.; Engelhard, I.M.; Van Den Hout, M.A.; Baas, J.M.P. Updated meta-analysis of classical fear conditioning in the anxiety disorders. Depress. Anxiety 2015, 32, 239–253. [Google Scholar] [CrossRef]
- Lissek, S.; Powers, A.S.; McClure, E.B.; Phelps, E.A.; Woldehawariat, G.; Grillon, C.; Pine, D.S. Classical fear conditioning in the anxiety disorders: A meta-analysis. Behav. Res. Ther. 2005, 43, 1391–1424. [Google Scholar] [CrossRef]
- Kida, S. Reconsolidation/destabilization, extinction and forgetting of fear memory as therapeutic targets for PTSD. Psychopharmacology 2019, 236, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bienvenu, T.C.M.; Dejean, C.; Jercog, D.; Aouizerate, B.; Lemoine, M.; Herry, C. The advent of fear conditioning as an animal model of post-traumatic stress disorder: Learning from the past to shape the future of PTSD research. Neuron 2021, 109, 2380–2397. [Google Scholar] [CrossRef]
- Battaglia, S.; Di Fazio, C.; Vicario, C.M.; Avenanti, A. Neuropharmacological modulation of N-methyl-D-aspartate, noradrenaline and endocannabinoid receptors in fear extinction learning: Synaptic transmission and plasticity. Int. J. Mol. Sci. 2023, 24, 5926. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, M.R.; Di Fazio, C.; Battaglia, S. Activated Tryptophan-Kynurenine Metabolic System in the Human Brain is Associated with Learned Fear. Front. Mol. Neurosci. 2023, 16, 1217090. [Google Scholar] [CrossRef]
- Vicario, C.M.; Makris, S.; Culicetto, L.; Lucifora, C.; Falzone, A.; Martino, G.; Ferraioli, F.; Nitsche, M.A.; Avenanti, A.; Craparo, G.C. Evidence of altered fear extinction learning in individuals with high vaccine hesitancy during COVID-19 pandemic. Clin. Neuropsychiatry 2023, in press. [Google Scholar]
- Maren, S. Neurobiology of Pavlovian fear conditioning. Annu. Rev. Neurosci. 2001, 24, 897–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia, S.; Thayer, J.F. Functional interplay between central and autonomic nervous systems in human fear conditioning. Trends Neurosci. 2022, 45, 504–506. [Google Scholar] [CrossRef]
- Haaker, J.; Maren, S.; Andreatta, M.; Merz, C.J.; Richter, J.; Richter, S.H.; Meir Drexler, S.; Lange, M.D.; Jüngling, K.; Nees, F.; et al. Making translation work: Harmonizing cross-species methodology in the behavioural neuroscience of Pavlovian fear conditioning. Neurosci. Biobehav. Rev. 2019, 107, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Vicario, C.M.; Martino, G. Dopamine and serotonin in fear extinction: Some key questions to be addressed. AIMS Neurosci. 2020, 7, 271. [Google Scholar] [CrossRef]
- Zabik, N.L.; Peters, C.; Iadipaolo, A.; Marusak, H.A.; Rabinak, C.A. Comparison of behavioral and brain indices of fear renewal during a standard vs. novel immersive reality Pavlovian fear extinction paradigm in healthy adults. Behav. Brain Res. 2023, 437, 114154. [Google Scholar] [CrossRef]
- Lucifora, C.; Grasso, G.M.; Perconti, P.; Plebe, A. Moral dilemmas in self-driving cars. Riv. Internaz. Filos. Psicol. 2020, 11, 238–250. [Google Scholar] [CrossRef]
- Lucifora, C.; Angelini, L.; Meteier, Q.; Vicario, C.M.; Khaled, O.A.; Mugellini, E.; Grasso, G.M. Cyber-therapy: The use of artificial intelligence in psychological practice. In Proceedings of the 4th International Conference on Intelligent Human Systems Integration (IHSI 2021): Integrating People and Intelligent Systems, Palermo, Italy, 22–24 February 2021; Springer: Berlin/Heidelberg, Germany, 2021; pp. 127–132. [Google Scholar]
- Grasso, G.M.; Lucifora, C.; Perconti, P.; Plebe, A. Integrating human acceptable morality in autonomous vehicles. In Proceedings of the 3rd International Conference on Intelligent Human Systems Integration (IHSI 2020): Integrating People and Intelligent Systems, Modena, Italy, 19–21 February 2020; Springer: Berlin/Heidelberg, Germany, 2020; pp. 41–45. [Google Scholar]
- Grasso, G.M.; Lucifora, C.; Perconti, P.; Plebe, A. Evaluating Mentalization during Driving. In Proceedings of the 5th International Conference on Vehicle Technology and Intelligent Transport Systems (VEHITS 2019), Crete, Greece, 3–5 May 2019; pp. 536–541. [Google Scholar]
- Daher, K.; Capallera, M.; Lucifora, C.; Casas, J.; Meteier, Q.; El Kamali, M.; El Ali, A.; Grosso, G.M.; Chollet, G.; Abou Khaled, O. Empathic interactions in automated vehicles# EmpathicCHI. In Proceedings of the Extended Abstracts of the 2021 CHI Conference on Human Factors in Computing Systems, Yokohama, Japan, 8–13 May 2021; pp. 1–4. [Google Scholar]
- Lucifora, C.; Grasso, G.M.; Perconti, P.; Plebe, A. Moral reasoning and automatic risk reaction during driving. Cogn. Technol. Work 2020, 23, 705–713. [Google Scholar] [CrossRef]
- Battaglia, S.; Garofalo, S.; di Pellegrino, G.; Starita, F. Revaluing the role of vmPFC in the acquisition of pavlovian threat conditioning in humans. J. Neurosci. 2020, 40, 8491–8500. [Google Scholar] [CrossRef]
- Fullana, M.A.; Dunsmoor, J.E.; Schruers, K.R.J.; Savage, H.S.; Bach, D.R.; Harrison, B.J. Human fear conditioning: From neuroscience to the clinic. Behav. Res. Ther. 2020, 124, 103528. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Jimenez, B.; Albajes-Eizagirre, A.; Lazarov, A.; Zhu, X.; Harrison, B.J.; Radua, J.; Neria, Y.; Fullana, M.A. Neural signatures of conditioning, extinction learning, and extinction recall in posttraumatic stress disorder: A meta-analysis of functional magnetic resonance imaging studies. Psychol. Med. 2020, 50, 1442–1451. [Google Scholar] [CrossRef]
- Vervliet, B.; Boddez, Y. Memories of 100 years of human fear conditioning research and expectations for its future 2020. Behav. Res. Ther. 2020, 135, 103732. [Google Scholar] [CrossRef]
- Battaglia, S.; Orsolini, S.; Borgomaneri, S.; Barbieri, R.; Diciotti, S.; di Pellegrino, G. Characterizing cardiac autonomic dynamics of fear learning in humans. Psychophysiology 2022, 59, e14122. [Google Scholar] [CrossRef]
- Sehlmeyer, C.; Schöning, S.; Zwitserlood, P.; Pfleiderer, B.; Kircher, T.; Arolt, V.; Konrad, C. Human fear conditioning and extinction in neuroimaging: A systematic review. PLoS ONE 2009, 4, e5865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, M.R.; Nearing, K.I.; LeDoux, J.E.; Phelps, E.A. Neural Circuitry Underlying the Regulation of Conditioned Fear and Its Relation to Extinction. Neuron 2008, 59, 829–838. [Google Scholar] [CrossRef] [Green Version]
- LeDoux, J. Fear and the brain: Where have we been, and where are we going? Biol. Psychiatry 1998, 44, 1229–1238. [Google Scholar] [CrossRef]
- Corcoran, K.A.; Quirk, G.J. Recalling Safety: Cooperative Functions of the Ventromedial Prefrontal Cortex and the Hippocampus in Extinction. CNS Spectr. 2007, 12, 200–206. [Google Scholar] [CrossRef]
- Etkin, A.; Egner, T.; Kalisch, R. Emotional processing in anterior cingulate and medial prefrontal cortex. Trends Cogn. Sci. 2011, 15, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Marković, V.; Vicario, C.M.; Yavari, F.; Salehinejad, M.A.; Nitsche, M.A. A systematic review on the effect of transcranial direct current and magnetic stimulation on fear memory and extinction. Front. Hum. Neurosci. 2021, 15, 655947. [Google Scholar] [CrossRef] [PubMed]
- Quirk, G.J.; Mueller, D. Neural mechanisms of extinction learning and retrieval. Neuropsychopharmacology 2008, 33, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Vicario, C.M.; Nitsche, M.A.; Hoysted, I.; Yavari, F.; Avenanti, A.; Salehinejad, M.A.; Felmingham, K.L. Anodal transcranial direct current stimulation over the ventromedial prefrontal cortex enhances fear extinction in healthy humans: A single blind sham-controlled study. Brain Stimul. 2019, 13, 489–491. [Google Scholar] [CrossRef] [Green Version]
- Milad, M.R.; Quirk, G.J.; Pitman, R.K.; Orr, S.P.; Fischl, B.; Rauch, S.L. A role for the human dorsal anterior cingulate cortex in fear expression. Biol. Psychiatry 2007, 62, 1191–1194. [Google Scholar] [CrossRef] [PubMed]
- Dixsaut, L.; Gräff, J. The Medial Prefrontal Cortex and Fear Memory: Dynamics, Connectivity, and Engrams. Int. J. Mol. Sci. 2021, 22, 12113. [Google Scholar] [CrossRef]
- Johansen, J.P.; Hamanaka, H.; Monfils, M.H.; Behnia, R.; Deisseroth, K.; Blair, H.T.; LeDoux, J.E. Optical activation of lateral amygdala pyramidal cells instructs associative fear learning. Proc. Natl. Acad. Sci. USA 2010, 107, 12692–12697. [Google Scholar] [CrossRef]
- Tye, K.M.; Prakash, R.; Kim, S.-Y.Y.; Fenno, L.E.; Grosenick, L.; Zarabi, H.; Thompson, K.R.; Gradinaru, V.; Ramakrishnan, C.; Deisseroth, K. Amygdala circuitry mediating reversible and bidirectional control of anxiety. Nature 2011, 471, 358–362. [Google Scholar] [CrossRef] [Green Version]
- Rogan, M.T.; Stäubli, U.V.; LeDoux, J.E. Fear conditioning induces associative long-term potentiation in the amygdala. Nature 1997, 390, 604–607. [Google Scholar] [CrossRef]
- Bouton, M.E. Context and behavioral processes in extinction. Learn. Mem. 2004, 11, 485–494. [Google Scholar] [CrossRef] [Green Version]
- Battaglia, S.; Cardellicchio, P.; Di Fazio, C.; Nazzi, C.; Fracasso, A.; Borgomaneri, S. Stopping in (e) motion: Reactive action inhibition when facing valence-independent emotional stimuli. Front. Behav. Neurosci. 2022, 16, 998714. [Google Scholar] [CrossRef]
- Battaglia, S.; Cardellicchio, P.; Di Fazio, C.; Nazzi, C.; Fracasso, A.; Borgomaneri, S. The influence of vicarious fear-learning in “infecting” reactive action inhibition. Front. Behav. Neurosci. 2022, 16, 946263. [Google Scholar] [CrossRef]
- Tovote, P.; Fadok, J.P.; Lüthi, A. Neuronal circuits for fear and anxiety. Nat. Rev. Neurosci. 2015, 16, 317–331. [Google Scholar] [CrossRef]
- Quirk, G.J.; Russo, G.K.; Barron, J.L.; Lebron, K. The role of ventromedial prefrontal cortex in the recovery of extinguished fear. J. Neurosci. 2000, 20, 6225–6231. [Google Scholar] [CrossRef] [PubMed]
- Milad, M.R.; Wright, C.I.; Orr, S.P.; Pitman, R.K.; Quirk, G.J.; Rauch, S.L. Recall of fear extinction in humans activates the ventromedial prefrontal cortex and hippocampus in concert. Biol. Psychiatry 2007, 62, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar] [CrossRef] [PubMed]
- Cornea-Hébert, V.; Riad, M.; Wu, C.; Singh, S.K.; Descarries, L. Cellular and subcellular distribution of the serotonin 5-HT2A receptor in the central nervous system of adult rat. J. Comp. Neurol. 1999, 409, 187–209. [Google Scholar] [CrossRef]
- Mascagni, F.; McDonald, A.J. A novel subpopulation of 5-HT type 3A receptor subunit immunoreactive interneurons in the rat basolateral amygdala. Neuroscience 2007, 144, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- Rainnie, D.G. Serotonergic modulation of neurotransmission in the rat basolateral amygdala. J. Neurophysiol. 1999, 82, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Bocchio, M.; McHugh, S.B.; Bannerman, D.M.; Sharp, T.; Capogna, M. Serotonin, amygdala and fear: Assembling the puzzle. Front. Neural Circuits 2016, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Parent, A.; Descarries, L.; Beaudet, A. Organization of ascending serotonin systems in the adult rat brain. A radioautographic study after intraventricular administration of [3H] 5-hydroxytryptamine. Neuroscience 1981, 6, 115–138. [Google Scholar] [CrossRef]
- Kawahara, H.; Yoshida, M.; Yokoo, H.; Nishi, M.; Tanaka, M. Psychological stress increases serotonin release in the rat amygdala and prefrontal cortex assessed by in vivo microdialysis. Neurosci. Lett. 1993, 162, 81–84. [Google Scholar] [CrossRef]
- Yokoyama, M.; Suzuki, E.; Sato, T.; Maruta, S.; Watanabe, S.; Miyaoka, H. Amygdalic levels of dopamine and serotonin rise upon exposure to conditioned fear stress without elevation of glutamate. Neurosci. Lett. 2005, 379, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Mo, B.; Feng, N.; Renner, K.; Forster, G. Restraint stress increases serotonin release in the central nucleus of the amygdala via activation of corticotropin-releasing factor receptors. Brain Res. Bull. 2008, 76, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, S.; Inoue, T.; Koyama, T. Effects of conditioned fear stress on serotonin neurotransmission and freezing behavior in rats. Eur. J. Pharmacol. 1999, 378, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, M.; Matsumoto, M.; Togashi, H.; Saito, H. Effects of conditioned fear stress on 5-HT release in the rat prefrontal cortex. Pharmacol. Biochem. Behav. 1995, 51, 515–519. [Google Scholar] [CrossRef] [PubMed]
- McGonigle, P.; Ruggeri, B. Animal models of human disease: Challenges in enabling translation. Biochem. Pharmacol. 2014, 87, 162–171. [Google Scholar] [CrossRef]
- Bezchlibnyk-Butler, K.; Aleksic, I.; Kennedy, S.H. Citalopram—A review of pharmacological and clinical effects. J. Psychiatry Neurosci. 2000, 25, 241. [Google Scholar]
- Gorman, J.M. Treating generalized anxiety disorder. J. Clin. Psychiatry 2003, 64, 24–29. [Google Scholar]
- Kent, J.M.; Coplan, J.D.; Gorman, J.M. Clinical utility of the selective serotonin reuptake inhibitors in the spectrum of anxiety. Biol. Psychiatry 1998, 44, 812–824. [Google Scholar] [CrossRef]
- Klumpp, H.; Fitzgerald, D.A.; Cook, E.; Shankman, S.A.; Angstadt, M.; Phan, K.L. Serotonin transporter gene alters insula activity to threat in social anxiety disorder. Neuroreport 2014, 25, 926. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, D.V.; Raj, B.A.; Trehan, R.R.; Knapp, E.L. Serotonin in panic disorder and social phobia. Int. Clin. Psychopharmacol. 1993, 8, 63–78. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Altman, D.; Antes, G.; Atkins, D.; Barbour, V.; Barrowman, N.; Berlin, J.A.; et al. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensman, R.; Guimarães, F.S.; Wang, M.; Deakin, J.F.W. Effects of ritanserin on aversive classical conditioning in humans. Psychopharmacology 1991, 104, 220–224. [Google Scholar] [CrossRef]
- Hetem, L.A.B.; De Souza, C.J.; Guimaràes, F.S.; Zuardi, A.W.; Graeff, F.G. Effect of d-fenfluramine on human experimental anxiety. Psychopharmacology 1996, 127, 276–282. [Google Scholar] [CrossRef]
- Hellewell, J.S.E.; Guimaraes, F.S.; Wang, M.; Deakin, J.F.W. Comparison of buspirone with diazepam and fluvoxamine on aversive classical conditioning in humans. J. Psychopharmacol. 1999, 13, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Hetem, L.A.B.; Guimarães, F.S.; Graeff, F.G. Opposite effects of nefazodone in two human models of anxiety. Psychopharmacology 2001, 156, 454–460. [Google Scholar] [CrossRef]
- Grillon, C.; Levenson, J.; Pine, D.S. A single dose of the selective serotonin reuptake inhibitor citalopram exacerbates anxiety in humans: A fear-potentiated startle study. Neuropsychopharmacology 2007, 32, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grillon, C.; Chavis, C.; Covington, M.F.; Pine, D.S. Two-week treatment with the selective serotonin reuptake inhibitor citalopram reduces contextual anxiety but not cued fear in healthy volunteers: A fear-potentiated startle study. Neuropsychopharmacology 2009, 34, 964–971. [Google Scholar] [CrossRef]
- Bui, E.; Orr, S.P.; Jacoby, R.J.; Keshaviah, A.; LeBlanc, N.J.; Milad, M.R.; Pollack, M.H.; Simon, N.M. Two weeks of pretreatment with escitalopram facilitates extinction learning in healthy individuals. Hum. Psychopharmacol. Clin. Exp. 2013, 28, 447–456. [Google Scholar] [CrossRef]
- Åhs, F.; Frick, A.; Furmark, T.; Fredrikson, M. Human serotonin transporter availability predicts fear conditioning. Int. J. Psychophysiol. 2015, 98, 515–519. [Google Scholar] [CrossRef]
- Lundquist, P.; Wilking, H.; Höglund, A.U.; Sandell, J.; Bergström, M.; Hartvig, P.; Långström, B. Potential of [11C] DASB for measuring endogenous serotonin with PET: Binding studies. Nucl. Med. Biol. 2005, 32, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Hindi Attar, C.; Finckh, B.; Büchel, C. The influence of serotonin on fear learning. PLoS ONE 2012, 7, e42397. [Google Scholar] [CrossRef] [PubMed]
- Gorka, S.M.; Lieberman, L.; Klumpp, H.; Kinney, K.L.; Kennedy, A.E.; Ajilore, O.; Francis, J.; Duffecy, J.; Craske, M.G.; Nathan, J.; et al. Reactivity to unpredictable threat as a treatment target for fear-based anxiety disorders. Psychol. Med. 2017, 47, 2450–2460. [Google Scholar] [CrossRef]
- Kałużna-Czaplińska, J.; Gątarek, P.; Chirumbolo, S.; Chartrand, M.S.; Bjørklund, G. How important is tryptophan in human health? Crit. Rev. Food Sci. Nutr. 2019, 59, 72–88. [Google Scholar] [CrossRef] [PubMed]
- Young, S.N. Acute tryptophan depletion in humans: A review of theoretical, practical and ethical aspects. J. Psychiatry Neurosci. JPN 2013, 38, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Robinson, O.J.; Overstreet, C.; Allen, P.S.; Pine, D.S.; Grillon, C. Acute tryptophan depletion increases translational indices of anxiety but not fear: Serotonergic modulation of the bed nucleus of the stria terminalis? Neuropsychopharmacology 2012, 37, 1963–1971. [Google Scholar] [CrossRef]
- Kanen, J.W.; Arntz, F.E.; Yellowlees, R.; Christmas, D.M.; Price, A.; Apergis-Schoute, A.M.; Sahakian, B.J.; Cardinal, R.N.; Robbins, T.W. Effect of tryptophandepletion on conditioned threat memory expression: Role of intolerance of uncertainty. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2021, 6, 590–598. [Google Scholar] [CrossRef]
- Garpenstrand, H.; Annas, P.; Ekblom, J.; Oreland, L.; Fredrikson, M. Human fear conditioning is related to dopaminergic and serotonergic biological markers. Behav. Neurosci. 2001, 115, 358–364. [Google Scholar] [CrossRef]
- Lonsdorf, T.B.; Weike, A.I.; Nikamo, P.; Schalling, M.; Hamm, A.O.; Öhman, A. Genetic gating of human fear learning and extinction: Possible implications for gene-environment interaction in anxiety disorder. Psychol. Sci. 2009, 20, 198–206. [Google Scholar] [CrossRef]
- Crişan, L.G.; Panǎ, S.; Vulturar, R.; Heilman, R.M.; Szekely, R.; Drugǎ, B.; Dragoş, N.; Miu, A.C. Genetic contributions of the serotonin transporter to social learning of fear and economic decision making. Soc. Cogn. Affect. Neurosci. 2009, 4, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Klumpers, F.; Heitland, I.; Oosting, R.S.; Kenemans, J.L.; Baas, J.M.P. Genetic variation in serotonin transporter function affects human fear expression indexed by fear-potentiated startle. Biol. Psychol. 2012, 89, 277–282. [Google Scholar] [CrossRef]
- Agren, T.; Furmark, T.; Eriksson, E.; Fredrikson, M. Human fear reconsolidation and allelic differences in serotonergic and dopaminergic genes. Transl. Psychiatry 2012, 2, e76. [Google Scholar] [CrossRef] [Green Version]
- Agren, T. Human reconsolidation: A reactivation and update. Brain Res. Bull. 2014, 105, 70–82. [Google Scholar] [CrossRef]
- Hermann, A.; Küpper, Y.; Schmitz, A.; Walter, B.; Vaitl, D.; Hennig, J.; Stark, R.; Tabbert, K. Functional Gene Polymorphisms in the Serotonin System and Traumatic Life Events Modulate the Neural Basis of Fear Acquisition and Extinction. PLoS ONE 2012, 7, e44352. [Google Scholar] [CrossRef]
- Klucken, T.; Alexander, N.; Schweckendiek, J.; Merz, C.J.; Kagerer, S.; Osinsky, R.; Walter, B.; Vaitl, D.; Hennig, J.; Stark, R. Individual differences in neural correlates of fear conditioning as a function of 5-HTTLPR and stressful life events. Soc. Cogn. Affect. Neurosci. 2013, 8, 318–325. [Google Scholar] [CrossRef] [Green Version]
- Glotzbach-Schoon, E.; Andreatta, M.; Reif, A.; Ewald, H.; Tröger, C.; Baumann, C.; Deckert, J.; Mühlberger, A.; Pauli, P. Contextual fear conditioning in virtual reality is affected by 5HTTLPR and NPSR1 polymorphisms: Effects on fear-potentiated startle. Front. Behav. Neurosci. 2013, 7, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heitland, I.; Groenink, L.; Bijlsma, E.Y.; Oosting, R.S.; Baas, J.M.P. Human Fear Acquisition Deficits in Relation to Genetic Variants of the Corticotropin Releasing Hormone Receptor 1 and the Serotonin Transporter. PLoS ONE 2013, 8, e63772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heitland, I.; Groenink, L.; van Gool, J.M.; Domschke, K.; Reif, A.; Baas, J.M.P. Human fear acquisition deficits in relation to genetic variants of the corticotropin-releasing hormone receptor 1 and the serotonin transporter—Revisited. Genes Brain Behav. 2016, 15, 209–220. [Google Scholar] [CrossRef]
- Straube, B.; Reif, A.; Richter, J.; Lueken, U.; Weber, H.; Arolt, V.; Jansen, A.; Zwanzger, P.; Domschke, K.; Pauli, P.; et al. The functional-1019C/G HTR1A polymorphism and mechanisms of fear. Transl. Psychiatry 2014, 4, e490. [Google Scholar] [CrossRef] [Green Version]
- Klumpers, F.; Kroes, M.C.; Heitland, I.; Everaerd, D.; Akkermans, S.E.A.; Oosting, R.S.; Van Wingen, G.; Franke, B.; Kenemans, J.L.; Fernández, G.; et al. Dorsomedial prefrontal cortex mediates the impact of serotonin transporter linked polymorphic region genotype on anticipatory threat reactions. Biol. Psychiatry 2015, 78, 582–589. [Google Scholar] [CrossRef]
- Baas, J.M.P.P.; Heitland, I. The impact of cue learning, trait anxiety and genetic variation in the serotonin 1A receptor on contextual fear. Int. J. Psychophysiol. 2015, 98, 506–514. [Google Scholar] [CrossRef]
- Wannemüller, A.; Moser, D.; Kumsta, R.; Jöhren, H.P.; Margraf, J. The Return of Fear: Variation of the Serotonin Transporter Gene Predicts Outcome of a Highly Standardized Exposure-Based One-Session Fear Treatment. Psychother. Psychosom. 2018, 87, 95–104. [Google Scholar] [CrossRef]
- Schipper, P.; Hiemstra, M.; Bosch, K.; Nieuwenhuis, D.; Adinolfi, A.; Glotzbach, S.; Borghans, B.; Lopresto, D.; Fernández, G.; Klumpers, F.; et al. The association between serotonin transporter availability and the neural correlates of fear bradycardia. Proc. Natl. Acad. Sci. USA 2019, 116, 25941–25947. [Google Scholar] [CrossRef] [Green Version]
- Sanders, M.J.; Wiltgen, B.J.; Fanselow, M.S. The place of the hippocampus in fear conditioning. Eur. J. Pharmacol. 2003, 463, 217–223. [Google Scholar] [CrossRef]
- Fullana, M.A.; Albajes-Eizagirre, A.; Soriano-Mas, C.; Vervliet, B.; Cardoner, N.; Benet, O.; Radua, J.; Harrison, B.J. Fear extinction in the human brain: A meta-analysis of fMRI studies in healthy participants. Neurosci. Biobehav. Rev. 2018, 88, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Ney, L.J.; Vicario, C.M.; Nitsche, M.A.; Felmingham, K.L. Timing matters: Transcranial direct current stimulation after extinction learning impairs subsequent fear extinction retention. Neurobiol. Learn. Mem. 2021, 177, 107356. [Google Scholar] [CrossRef]
- Van’t Wout, M.; Mariano, T.Y.; Garnaat, S.L.; Reddy, M.K.; Rasmussen, S.A.; Greenberg, B.D. Can Transcranial Direct Current Stimulation Augment Extinction of Conditioned Fear? Brain Stimul. 2016, 9, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Milad, M.R.; Quirk, G.J. Neurons in medial prefrontal cortex signal memory for fear extinction. Nature 2002, 420, 70–74. [Google Scholar] [CrossRef]
- Sengupta, A.; Holmes, A. A discrete dorsal raphe to basal amygdala 5-HT circuit calibrates aversive memory. Neuron 2019, 103, 489–505.e7. [Google Scholar] [CrossRef]
- Abela, A.R.; Browne, C.J.; Sargin, D.; Prevot, T.D.; Ji, X.D.; Li, Z.; Lambe, E.K.; Fletcher, P.J. Median raphe serotonin neurons promote anxiety-like behavior via inputs to the dorsal hippocampus. Neuropharmacology 2020, 168, 107985. [Google Scholar] [CrossRef]
- Blakely, R.D.; Berson, H.E.; Fremeau, R.T.; Caron, M.G.; Peek, M.M.; Prince, H.K.; Bradley, C.C. Cloning and expression of a functional serotonin transporter from rat brain. Nature 1991, 354, 66–70. [Google Scholar] [CrossRef]
- Lesch, K.-P.; Bengel, D.; Heils, A.; Sabol, S.Z.; Greenberg, B.D.; Petri, S.; Benjamin, J.; Müller, C.R.; Hamer, D.H.; Murphy, D.L. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 1996, 274, 1527–1531. [Google Scholar] [CrossRef]
- Jakubovski, E.; Johnson, J.A.; Nasir, M.; Müller-Vahl, K.; Bloch, M.H. Systematic review and meta-analysis: Dose–response curve of SSRIs and SNRIs in anxiety disorders. Depress. Anxiety 2019, 36, 198–212. [Google Scholar] [CrossRef]
- Edwards, J.G.; Anderson, I. Systematic review and guide to selection of selective serotonin reuptake inhibitors. Drugs 1999, 57, 507–533. [Google Scholar] [CrossRef]
- Frazer, A. Pharmacology of antidepressants. J. Clin. Psychopharmacol. 1997, 17, 2S–18S. [Google Scholar] [CrossRef]
- Hiemke, C.; Härtter, S. Pharmacokinetics of selective serotonin reuptake inhibitors. Pharmacol. Ther. 2000, 85, 11–28. [Google Scholar] [CrossRef]
- Attenburrow, M.-J.; Mitter, P.; Whale, R.; Terao, T.; Cowen, P. Low-dose citalopram as a 5-HT neuroendocrine probe. Psychopharmacology 2001, 155, 323–326. [Google Scholar] [CrossRef]
- Schulkin, J.; Morgan, M.A.; Rosen, J.B. A neuroendocrine mechanism for sustaining fear. Trends Neurosci. 2005, 28, 629–635. [Google Scholar] [CrossRef]
- Crockett, M.J.; Clark, L.; Roiser, J.P.; Robinson, O.J.; Cools, R.; Chase, H.W.; Den Ouden, H.; Apergis-Schoute, A.; Campbell-Meikeljohn, D.; Seymour, B. Converging evidence for central 5-HT effects in acute tryptophan depletion. Mol. Psychiatry 2012, 17, 121–123. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, S.; Benkelfat, C.; Young, S.N.; Leyton, M.; de Mzengeza, S.; De Montigny, C.; Blier, P.; Diksic, M. Differences between males and females in rates of serotonin synthesis in human brain. Proc. Natl. Acad. Sci. USA 1997, 94, 5308–5313. [Google Scholar] [CrossRef]
- Bel, N.; Artigas, F. Reduction of serotonergic function in rat brain by tryptophan depletion: Effects in control and fluvoxamine-treated rats. J. Neurochem. 1996, 67, 669–676. [Google Scholar] [CrossRef]
- Hettema, J.M.; Annas, P.; Neale, M.C.; Kendler, K.S.; Fredrikson, M. A twin study of the genetics of fear conditioning. Arch. Gen. Psychiatry 2003, 60, 702–708. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.A.; Hen, R. Genetic approaches to the study of anxiety. Annu. Rev. Neurosci. 2004, 27, 193–222. [Google Scholar] [CrossRef]
- Dougherty, L.R.; Tolep, M.R.; Bufferd, S.J.; Olino, T.M.; Dyson, M.; Traditi, J.; Rose, S.; Carlson, G.A.; Klein, D.N. Preschool anxiety disorders: Comprehensive assessment of clinical, demographic, temperamental, familial, and life stress correlates. J. Clin. Child Adolesc. Psychol. 2013, 42, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, A.; Nakao, K.; Tokuyama, M.; Takeda, M. Prediction of first episode of panic attack among white-collar workers. Psychiatry Clin. Neurosci. 2005, 59, 119–126. [Google Scholar] [CrossRef]
- Nugent, N.R.; Tyrka, A.R.; Carpenter, L.L.; Price, L.H. Gene–environment interactions: Early life stress and risk for depressive and anxiety disorders. Psychopharmacology 2011, 214, 175–196. [Google Scholar] [CrossRef] [Green Version]
- Lonsdorf, T.B.; Kalisch, R. A review on experimental and clinical genetic associations studies on fear conditioning, extinction and cognitive-behavioral treatment. Transl. Psychiatry 2011, 1, e41. [Google Scholar] [CrossRef] [Green Version]
- Hariri, A.R.; Mattay, V.S.; Tessitore, A.; Kolachana, B.; Fera, F.; Goldman, D.; Egan, M.F.; Weinberger, D.R. Serotonin transporter genetic variation and the response of the human amygdala. Science 2002, 297, 400–403. [Google Scholar] [CrossRef] [Green Version]
- Canli, T.; Lesch, K.-P. Long story short: The serotonin transporter in emotion regulation and social cognition. Nat. Neurosci. 2007, 10, 1103–1109. [Google Scholar] [CrossRef]
- Dannlowski, U.; Konrad, C.; Kugel, H.; Zwitserlood, P.; Domschke, K.; Schöning, S.; Ohrmann, P.; Bauer, J.; Pyka, M.; Hohoff, C. Emotion specific modulation of automatic amygdala responses by 5-HTTLPR genotype. Neuroimage 2010, 53, 893–898. [Google Scholar] [CrossRef]
- Heinz, A.; Braus, D.F.; Smolka, M.N.; Wrase, J.; Puls, I.; Hermann, D.; Klein, S.; Grüsser, S.M.; Flor, H.; Schumann, G. Amygdala-prefrontal coupling depends on a genetic variation of the serotonin transporter. Nat. Neurosci. 2005, 8, 20–21. [Google Scholar] [CrossRef]
- Munafo, M.R.; Clark, T.; Flint, J. Does measurement instrument moderate the association between the serotonin transporter gene and anxiety-related personality traits? A meta-analysis. Mol. Psychiatry 2005, 10, 415–419. [Google Scholar] [CrossRef] [Green Version]
- Ippolito, G.; Bertaccini, R.; Tarasi, L.; Di Gregorio, F.; Trajkovic, J.; Battaglia, S.; Romei, V. The role of alpha oscillations among the main neuropsychiatric disorders in the adult and developing human brain: Evidence from the last 10 years of research. Biomedicines 2022, 10, 3189. [Google Scholar] [CrossRef]
- Di Gregorio, F.; La Porta, F.; Petrone, V.; Battaglia, S.; Orlandi, S.; Ippolito, G.; Romei, V.; Piperno, R.; Lullini, G. Accuracy of EEG biomarkers in the detection of clinical outcome in disorders of consciousness after severe acquired brain injury: Preliminary results of a pilot study using a machine learning approach. Biomedicines 2022, 10, 1897. [Google Scholar] [CrossRef]
- Tanaka, M.; Diano, M.; Battaglia, S. Editorial: Insights into structural and functional organization of the brain: Evidence from neuroimaging and non-invasive brain stimulation techniques. Front. Psychiatry 2023, 14, 1225755. [Google Scholar] [CrossRef]
- Di Gregorio, F.; Battaglia, S. Advances in EEG-based functional connectivity approaches to the study of the central nervous system in health and disease. Adv. Clin. Exp. Med. 2023, 32, 607–612. [Google Scholar] [CrossRef]
- Bennett, A.J.; Lesch, K.P.; Heils, A.; Long, J.C.; Lorenz, J.G.; Shoaf, S.E.; Champoux, M.; Suomi, S.J.; Linnoila, M.V.; Higley, J.D. Early experience and serotonin transporter gene variation interact to influence primate CNS function. Mol. Psychiatry 2002, 7, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Songtachalert, T.; Roomruangwong, C.; Carvalho, A.F.; Bourin, M.; Maes, M. Anxiety Disorders: Sex Differences in Serotonin and Tryptophan Metabolism. Curr. Top. Med. Chem. 2018, 18, 1704–1715. [Google Scholar] [CrossRef] [Green Version]
- Mitsushima, D.; Yamada, K.; Takase, K.; Funabashi, T.; Kimura, F. Sex differences in the basolateral amygdala: The extracellular levels of serotonin and dopamine, and their responses to restraint stress in rats. Eur. J. Neurosci. 2006, 24, 3245–3254. [Google Scholar] [CrossRef]
- Willadsen, M.; Üngör, M.; Sługocka, A.; Schwarting, R.K.W.; Homberg, J.R.; Wöhr, M. Fear Extinction and Predictive Trait-Like Inter-Individual Differences in Rats Lacking the Serotonin Transporter. Int. J. Mol. Sci. 2021, 22, 7088. [Google Scholar] [CrossRef]
- Barton, C.; Sklenicka, J.; Sayegh, P.; Yaffe, K.; Schubert, C.C.; Boustani, M.; Callahan, C.M.; Perkins, A.J.; Carney, C.P.; Fox, C.; et al. A double-blind trial of bupropion versus desipramine for bipolar depression. J. Clin. Psychiatry 2003, 55, 391–393. [Google Scholar]
- Pettersson, R.; Hagsäter, S.M.; Eriksson, E. Serotonin depletion eliminates sex differences with respect to context-conditioned immobility in rat. Psychopharmacology 2016, 233, 1513–1521. [Google Scholar] [CrossRef]
- Donner, N.C.; Lowry, C.A. Sex differences in anxiety and emotional behavior. Pflugers Arch. 2013, 465, 601–626. [Google Scholar] [CrossRef]
- Vicario, C.M.; Nitsche, M.A.; Felmingham, K. Forgetting fear associations through tES: Which memory process might be critical? Transl. Psychiatry 2017, 7, e1046. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Godoy, M.; Aunger, R.; Curtis, V. Serotonin—A link between disgust and immunity? Med. Hypotheses 2007, 68, 61–66. [Google Scholar] [CrossRef]
- Limebeer, C.L.; Parker, L.A.; Fletcher, P.J. 5,7-dihydroxytryptamine lesions of the dorsal and median raphe nuclei interfere with lithium-induced conditioned gaping, but not conditioned taste avoidance, in rats. Behav. Neurosci. 2004, 118, 1391. [Google Scholar] [CrossRef] [PubMed]
- Vicario, C.M. Altered insula response to sweet taste processing in recovered anorexia and bulimia nervosa: A matter of disgust sensitivity? Am. J. Psychiatry 2013, 170, 1497. [Google Scholar] [CrossRef]
- Vicario, C.M.; Rafal, R.D.; Borgomaneri, S.; Paracampo, R.; Kritikos, A.; Avenanti, A. Pictures of disgusting foods and disgusted facial expressions suppress the tongue motor cortex. Soc. Cogn. Affect. Neurosci. 2017, 12, 352–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicario, C.M.; Rafal, R.D.; Di Pellegrino, G.; Lucifora, C.; Salehinejad, M.A.; Nitsche, M.A.; Avenanti, A. Indignation for moral violations suppresses the tongue motor cortex: Preliminary TMS evidence. Soc. Cogn. Affect. Neurosci. 2022, 17, 151–159. [Google Scholar] [CrossRef]
- Vicario, C.M.; Lucifora, C. Neuroethics: What the study of brain disorders can tell about moral behavior. AIMS Neurosci. 2021, 8, 543–547. [Google Scholar] [CrossRef]
- Lucifora, C.; Martino, G.; Curcuruto, A.; Salehinejad, M.A.; Vicario, C.M. How self-control predicts moral decision making: An exploratory study on healthy participants. Int. J. Environ. Res. Public Health 2021, 18, 3840. [Google Scholar] [CrossRef]
- Vicario, C.M.; Turrini, S.; Lucifora, C.; Culicetto, L.; Ferraioli, F.; Falzone, A.; Nitsche, M.A.; Avenanti, A. When defeat leaves a bad taste in the mouth: Modulation of tongue corticobulbar output during monetary loss in a gambling task. Brain Stimul. 2022, 15, 1448–1450. [Google Scholar] [CrossRef]
- Vicario, C.M.; Rafal, R.D.; Avenanti, A. Counterfactual thinking affects the excitability of the motor cortex. Cortex 2015, 65, 139–148. [Google Scholar] [CrossRef]
- Coppini, S.; Lucifora, C.; Vicario, C.M.; Gangemi, A. Experiments on real-life emotions challenge Ekman’s model. Sci. Rep. 2023, 13, 9511. [Google Scholar] [CrossRef]
- Barchetta, S.; Martino, G.; Craparo, G.; Salehinejad, M.A.; Nitsche, M.A.; Vicario, C.M. Alexithymia is linked with a negative bias for past and current events in healthy humans. Int. J. Environ. Res. Public Health 2021, 18, 6696. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Vitale, F.; Avenanti, A. Early changes in corticospinal excitability when seeing fearful body expressions. Sci. Rep. 2015, 5, 14122. [Google Scholar] [CrossRef] [Green Version]
- Borgomaneri, S.; Vitale, F.; Avenanti, A. Behavioral inhibition system sensitivity enhances motor cortex suppression when watching fearful body expressions. Brain Struct. Funct. 2017, 222, 3267–3282. [Google Scholar] [CrossRef]
- Andreatta, M.; Pauli, P. Contextual modulation of conditioned responses in humans: A review on virtual reality studies. Clin. Psychol. Rev. 2021, 90, 102095. [Google Scholar] [CrossRef]
- Andreatta, M.; Winkler, M.H.; Collins, P.; Gromer, D.; Gall, D.; Pauli, P.; Gamer, M. VR for studying the neuroscience of emotional responses. In Current Topics in Behavioral Neurosciences; Geyer, M.A., Marsden, C.A., Ellenbroek, B.A., Barnes, T.R.E., Andersen, S.L., Paulus, M.P., Olivier, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2023. [Google Scholar] [CrossRef]
- Fiori, F.; Chiappini, E.; Soriano, M.; Paracampo, R.; Romei, V.; Borgomaneri, S.; Avenanti, A. Long-latency interhemispheric interactions between motor-related areas and the primary motor cortex: A dual site TMS study. Sci. Rep. 2017, 7, 14936. [Google Scholar] [CrossRef] [Green Version]
- Fiori, F.; Chiappini, E.; Avenanti, A. Enhanced action performance following TMS manipulation of associative plasticity in ventral premotor-motor pathway. Neuroimage 2018, 183, 847–858. [Google Scholar] [CrossRef]
- Chiappini, E.; Borgomaneri, S.; Marangon, M.; Turrini, S.; Romei, V.; Avenanti, A. Driving associative plasticity in premotor-motor connections through a novel paired associative stimulation based on long-latency cortico-cortical interactions. Brain Stimul. 2020, 13, 1461–1463. [Google Scholar] [CrossRef]
- Valchev, N.; Tidoni, E.; Hamilton, A.F.D.C.; Gazzola, V.; Avenanti, A. Primary somatosensory cortex necessary for the perception of weight from other people’s action: A continuous theta-burst TMS experiment. Neuroimage 2017, 152, 195–206. [Google Scholar] [CrossRef]
- Turrini, S.; Fiori, F.; Chiappini, E.; Lucero, B.; Santarnecchi, E.; Avenanti, A. Cortico-cortical paired associative stimulation (ccPAS) over premotor-motor areas affects local circuitries in the human motor cortex via Hebbian plasticity. Neuroimage 2023, 271, 120027. [Google Scholar] [CrossRef]
- Lefaucheur, J.P.; Aleman, A.; Baeken, C.; Benninger, D.H.; Brunelin, J.; Di Lazzaro, V.; Filipović, S.R.; Grefkes, C.; Hasan, A.; Hummel, F.C.; et al. Evidence-based guidelines on the therapeutic use of repetitive transcranial magnetic stimulation (rTMS): An update (2014–2018). Clin. Neurophysiol. 2020, 131, 474–528. [Google Scholar] [CrossRef]
- Avenanti, A.; Coccia, M.; Ladavas, E.; Provinciali, L.; Ceravolo, M.G. Low-frequency rTMS promotes use-dependent motor plasticity in chronic stroke: A randomized trial. Neurology 2012, 78, 256–264. [Google Scholar] [CrossRef]
- Salehinejad, M.A.; Nejati, V.; Mosayebi-Samani, M.; Mohammadi, A.; Wischnewski, M.; Kuo, M.F.; Avenanti, A.; Vicario, C.M.; Nitsche, M.A. Transcranial direct current stimulation in ADHD: A systematic review of efficacy, safety, and protocol-induced electrical field modeling results. Neurosci. Bull. 2020, 36, 1191–1212. [Google Scholar] [CrossRef]
- Casula, A.; Milazzo, B.M.; Martino, G.; Sergi, A.; Lucifora, C.; Tomaiuolo, F.; Quartarone, A.; Nitsche, M.A.; Vicario, C.M. Non-Invasive brain stimulation for the modulation of aggressive behavior-A systematic review of randomized sham-controlled studies. Life 2023, 13, 1220. [Google Scholar] [CrossRef]
- Turrini, S.; Wong, B.; Eldaief, M.; Press, D.Z.; Sinclair, D.A.; Koch, G.; Avenanti, A.; Santarnecchi, E. The multifactorial nature of healthy brain ageing: Brain changes, functional decline and protective factors. Ageing Res. Rev. 2023, 88, 101939. [Google Scholar] [CrossRef]
- Vicario, C.M.; Salehinejad, M.A.; Felmingham, K.; Martino, G.; Nitsche, M.A. A systematic review on the therapeutic effectiveness of non-invasive brain stimulation for the treatment of anxiety disorders. Neurosci. Biobehav. Rev. 2019, 96, 219–231. [Google Scholar] [CrossRef]
- Vicario, C.M.; Salehinejad, M.A.; Avenanti, A.; Nitsche, M.A. Transcranial direct current stimulation (tDCS) in anxiety disorders. In Non Invasive Brain Stimulation in Psychiatry and Clinical Neurosciences; Dell’Osso, B., Di Lorenzo, G., Eds.; Springer: Cham, Switzerland, 2020; pp. 301–317. [Google Scholar] [CrossRef]
- Vicario, C.M.; Salehinejad, M.A.; Lucifora, C.; Martino, G.; Falzone, A.M.; Grasso, G.; Nitsche Michael, A. Combining virtual reality exposure therapy with non-invasive brain stimulation for the treatment of post-traumatic stress disorder and related syndromes: A perspective. In Translational Methods for PTSD Research; Pinna, G., Ed.; Neuromethods; Humana Press: New York, NY, USA, 2023; Volume 198, pp. 231–245. [Google Scholar] [CrossRef]


{kind=link}
| Study | Design | Outcome Measure(s) | Type of Manipulation | Effect of Manipulation | N (Male/Female) | Type of CS/US | Brief Statement of Findings |
|---|---|---|---|---|---|---|---|
| Hensman et al. (1991) [118] | Between group double blind | SCR | Ritanserin (10 mg) | 5-HT antagonist | 30 (15/15) | Tone/White noise | Reduced SCR amplitude and spontaneous fluctuations in the extinction phase |
| Hetem et al. (1996) [119] | Between group double blind | SCR | d-fenfluramine (15 mg or 30 mg) | 5-HT agonist | 42 (27/15) | Tone/White noise | Decreased anxiety during a public speaking task but no effects on FC |
| Hellewell et al. (1999) [120] | Between group single blind | SCR | (a) Buspirone (5 mg), (b) Diazepam (2 mg), (c) Fluvoxamine (25 mg) | (a) Azapirone (b) Benzodia-zepine (c) SSRI | 50 (25/25) | Tone/White noise | Decreased SCR in both habituation and extinction |
| Silva et al. (2001) [121] | Between group double blind | SCR | Nefazodone (100 mg or 200 mg) | SSRI | 29 (9/20) | Tone/White noise | Decreased level of SCR fluctuation in both habituation and extinction |
| Grillon et al. (2007) [122] | Within group Double blind cross over | Eye blink startle response | Single dose of Citalopram (20 mg) | SSRI | 18 (9/9) | Geometric colored shapes/Electric Shock | Increased phasic and sustained fear-potentiated startle |
| Grillon et al. (2008) [123] | Between group double blind | Eye blink startle response | Two weeks of Citalopram (20 mg) | SSRI | 28 (19/9) | Geometric colored shapes/Electric shock | Decreased sustained anxiety but not phasic cued fear potentiation |
| Bui et al. (2012) [124] | Double blind randomized placebo controlled | SCR | Two weeks of Escitalopram (10 mg) | SSRI | 38 (23/15) | Yellow circle, white square/Electric shock | Facilitation of extinction learning but not acquisition |
| Åhs et al. (2015) [125] | Within group | SCR PET | NA ** | NA | 16 (8/8) | Geometric shapes/Electric shock | Lower 5-HTT in amygdala, insula, and dorsal anterior cingulate cortex predicts increased SCR during fear learning |
| Gorka et al. (2017) [128] | Mixed group | Eye blink startle response | Two weeks of Sertraline, Fluoxetine, Paroxetine, Escitalopram, or Citalopram (variable dose) | SSRI | 72 (52/20) | Geometric colored shapes/Electric shock | No effect of SSRI administration on magnitude of startle response |
| Study | Design | Outcome Measure(s) | Type of Manipulation | N (Male/Female) | Type CS/US | Brief Statement of Findings |
|---|---|---|---|---|---|---|
| Robinson et al. (2012) [131] | Within group | Eye blink startle response | Dietary | 20 (13/7) | Geometric shapes/Electric shock | Increased startle potentiation to contextual threat (sustained anxiety) but not explicit threat cues (cued fear) |
| Hindi Attar et al. (2012) [127] | Mixed design Between subject Double blind | SCR fMRI | Dietary | 39 (39/0) | Geometrical shapes/Increased temperature | Decreased fear response during fear acquisition in both fMRI and autonomic responses |
| Kanen et al. (2021) [132] | Between group double blind | SCR | Dietary | 47 (29/18) | Geometrical shapes/Electric shock | Decreased fear response in spontaneous recovery |
| Study | Outcome Measure(s) | Polymor-phism | N (Male/Female) | Type of CS/US | Brief Statement of Findings |
|---|---|---|---|---|---|
| Garpenstrand et al. (2001) [133] | SCR | 5-HTT MAOA | 40 (14/26) | Geometric shapes/Electric shock | Both 5-HTT and MAOA polymorphisms could explain variation in fear learning (S allele carriers showed increased fear response during acquisition) |
| Lonsdorf et al. (2009) [134] | Eye blink startle response SCR | 5-HTT | 48 (25/23) | Face stimuli/Electric shock | Only carriers of S allele exhibited conditioned startle potentiation in both acquisition and extinction |
| Crisan et al. (2009) [135] | SCR Electrocardiogram | 5-HTT | 32 (9/23) | Geometric shape/Electric shock | Increased fear response during social FC task in 5-HT S-carriers |
| Klumpers et al. (2011) [136] | Eye blink startle response | 5-HTT | 82 (22/62) | Face stimuli/Electric shock | S carriers showed stronger FPS during fear learning but no deficit in fear downregulation |
| Agren et al. (2012) [137] | SCR | 5-HTT COMT | 66 (28/38) | Images of colorful lamps/Electric shock | S allele carriers who underwent extinction outside reconsolidation time window showed an increased fear response in the reacquisition phase (day 3). |
| Hartley et al. (2012) [31] | SCR | STPP | 141 (54/87) | Geometric shapes/Electric shock | Increased spontaneous fear recovery and impaired extinction retention in G allele carriers of STPP |
| Hermann et al. (2012) [88] | SCR fMRI | 5-HTT TPH2 | 74 (37/37) | Geometric shapes/Electric shock | Increased Insula activation in response to both CS and US in S allele carriers |
| Klucken et al. (2013) [140] | SCR fMRI | 5-HTT | 47 (47/0) | Geometric shapes/Aversive pictures | Greater response to CS+ in right amygdala, left thalamus, bilateral insula and bilateral occipital cortex in S allele carriers |
| Glotzbach-Schoon et al. (2013) [141] | Eye blink startle response SCR | 5-HTT NPSR1 | 80 (31/49) | Context ***/Electric shocks | Only carriers of both S+/T+ alleles showed higher startle responses in acquisition |
| Heitland et al. (2013) [142] | Eye blink startle response | 5-HTT CRHR1 | 146 (59/87) | Context ***/Electric shock | Increased FPS to threat context in participants carrying both G allele and S allele during uninstructed FC |
| Heitland et al. (2016) [143] | Eye blink startle response | 5-HTT CRHR1 | 205 (59/87) | Context ***/Electric shock | Additional interaction between CRHR1 and triallelic 5-HTTLPR variant on acquisition |
| Straube et al. (2014) [144] | fMRI SCR | HTR1A | 39 (13/26) | Geometric shapes/White noise | Increased activation of the amygdala and hippocampus in response to fear cues and safety cues before and after treatment in GG carriers |
| Klumpers et al. (2015) [145] | SCR Eye blink startle response fMRI | 5-HTT | 168 (120–48) | Geometric shapes, face stimuli/Electric shock | Fear related dmPFC was increased in S allele carriers during threat learning |
| Baas & Heitland (2015) [146] | Eye blink startle response | 5-HTT | 150 (60/90) | Context ***/Electric shock | Enhanced contextual FPS response in C-carriers |
| Wannemueller et al. (2018) [147] | Self-reports | 5-HTT | 157 (25/132) | Fear mental images | Phobic S allele carriers showed greater return of fear 7 months after undergoing exposure-based phobia treatments |
| Schipper et al. (2019) [148] | Heart rate | 5-HTT | 104 (104/0) | Visual stimuli/Electric shock | S allele carriers displayed significantly stronger fear bradycardia to CS+ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tortora, F.; Hadipour, A.L.; Battaglia, S.; Falzone, A.; Avenanti, A.; Vicario, C.M. The Role of Serotonin in Fear Learning and Memory: A Systematic Review of Human Studies. Brain Sci. 2023, 13, 1197. https://doi.org/10.3390/brainsci13081197
Tortora F, Hadipour AL, Battaglia S, Falzone A, Avenanti A, Vicario CM. The Role of Serotonin in Fear Learning and Memory: A Systematic Review of Human Studies. Brain Sciences. 2023; 13(8):1197. https://doi.org/10.3390/brainsci13081197
Chicago/Turabian StyleTortora, Francesco, Abed L. Hadipour, Simone Battaglia, Alessandra Falzone, Alessio Avenanti, and Carmelo M. Vicario. 2023. "The Role of Serotonin in Fear Learning and Memory: A Systematic Review of Human Studies" Brain Sciences 13, no. 8: 1197. https://doi.org/10.3390/brainsci13081197