
The ‘Postural Rhythm’ of the Ground Reaction Force during Upright Stance and Its Conversion to Body Sway—The Effect of Vision, Support Surface and Adaptation to Repeated Trials


Abstract
:1. Introduction
2. Methods
2.1. Participants and Procedures
2.2. Data Acquisition and Analysis
2.3. Frequency–Domain Analysis
2.4. Time–Domain Analysis
2.5. Statistical Analysis
3. Results
3.1. The Vertical Component of the Ground Reaction Force (VGRF)
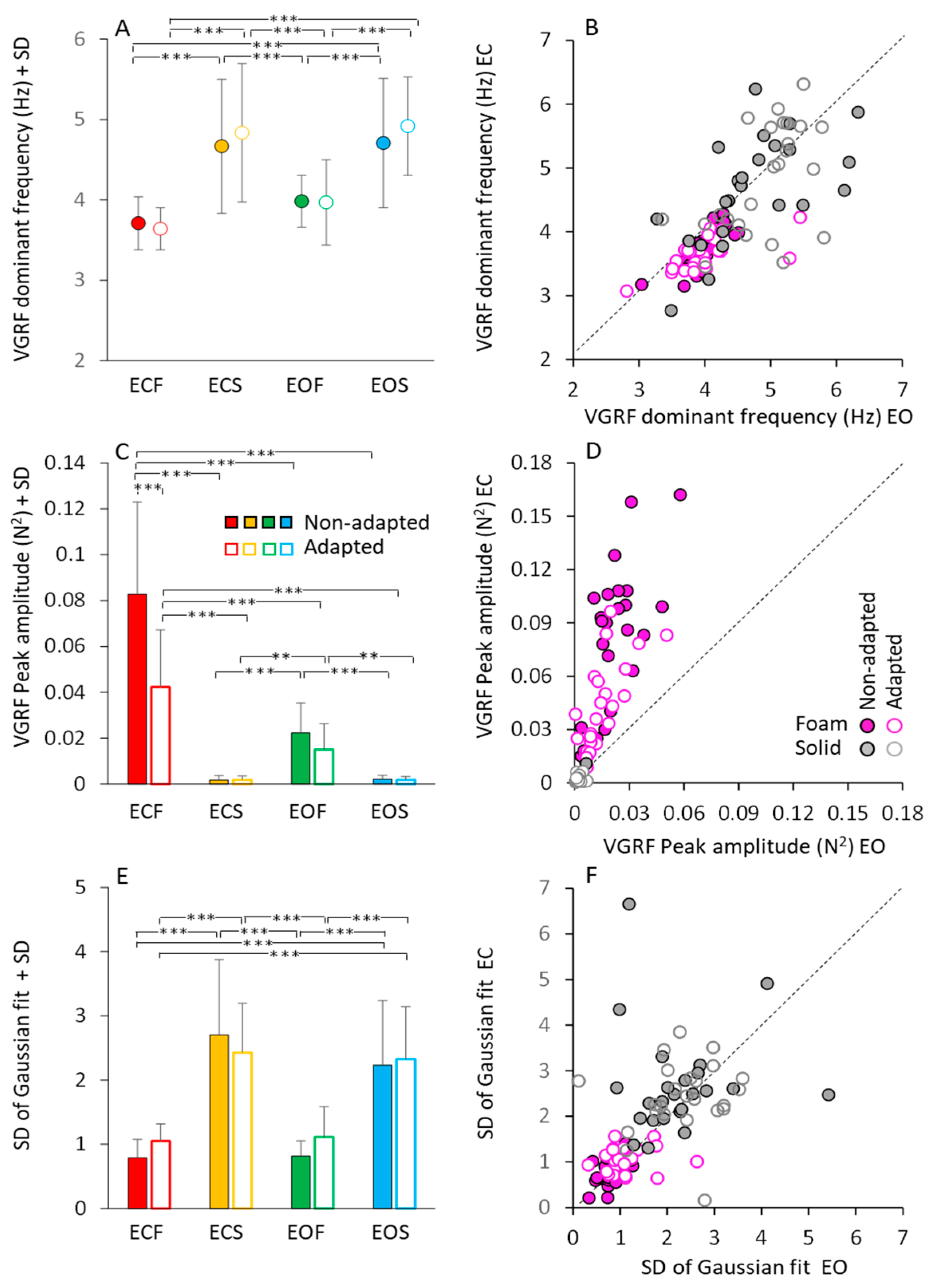
3.2. The Dominant Frequency of the VGRF Oscillation
3.3. Modulation of Peak Amplitude and Dominant Frequency of the VGRF Spectrum by Vision, Support Surface, and Adaptation
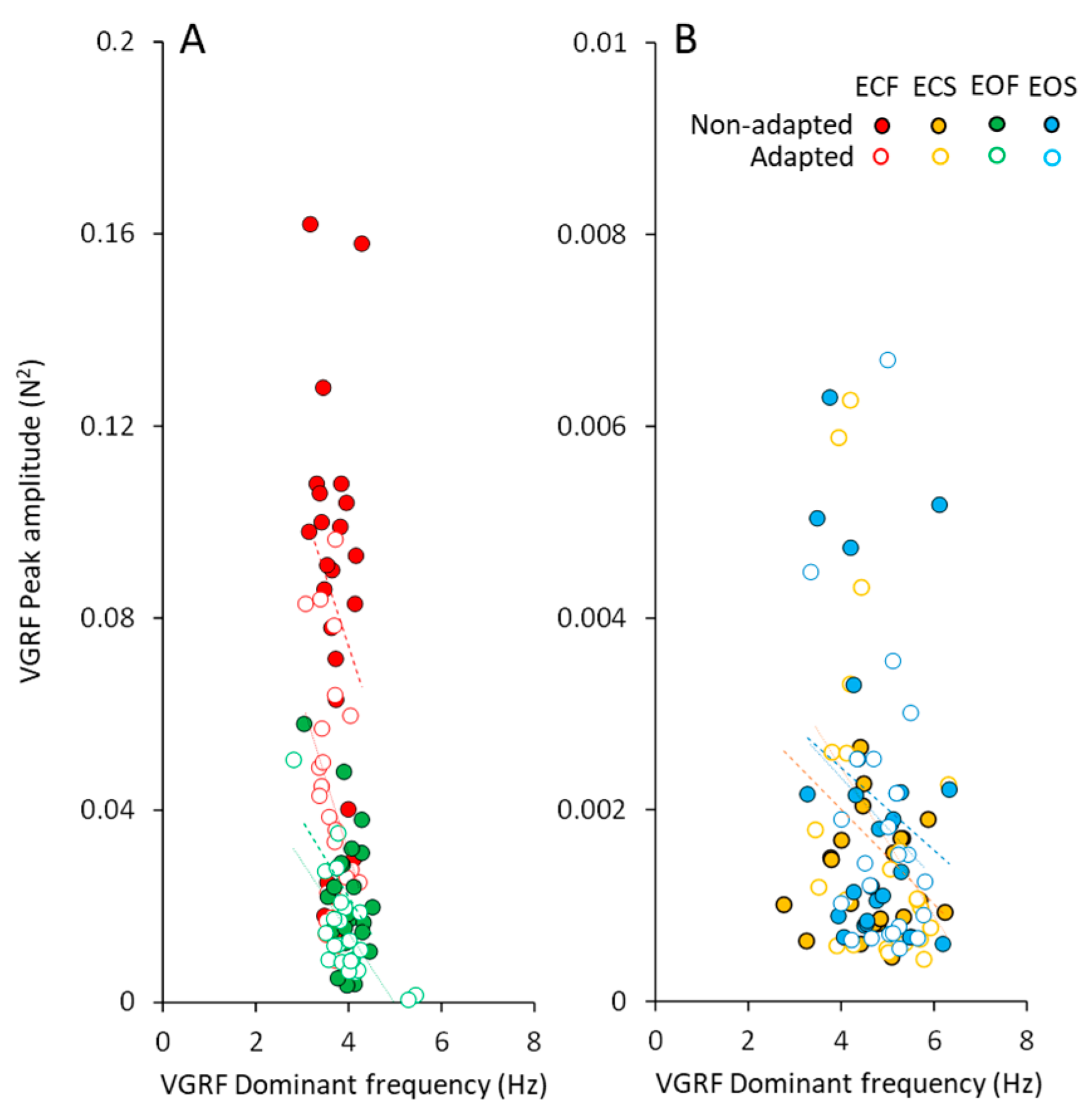
3.4. Body Weight Modifies Both Frequency and Amplitude of the VGRF Spectrum
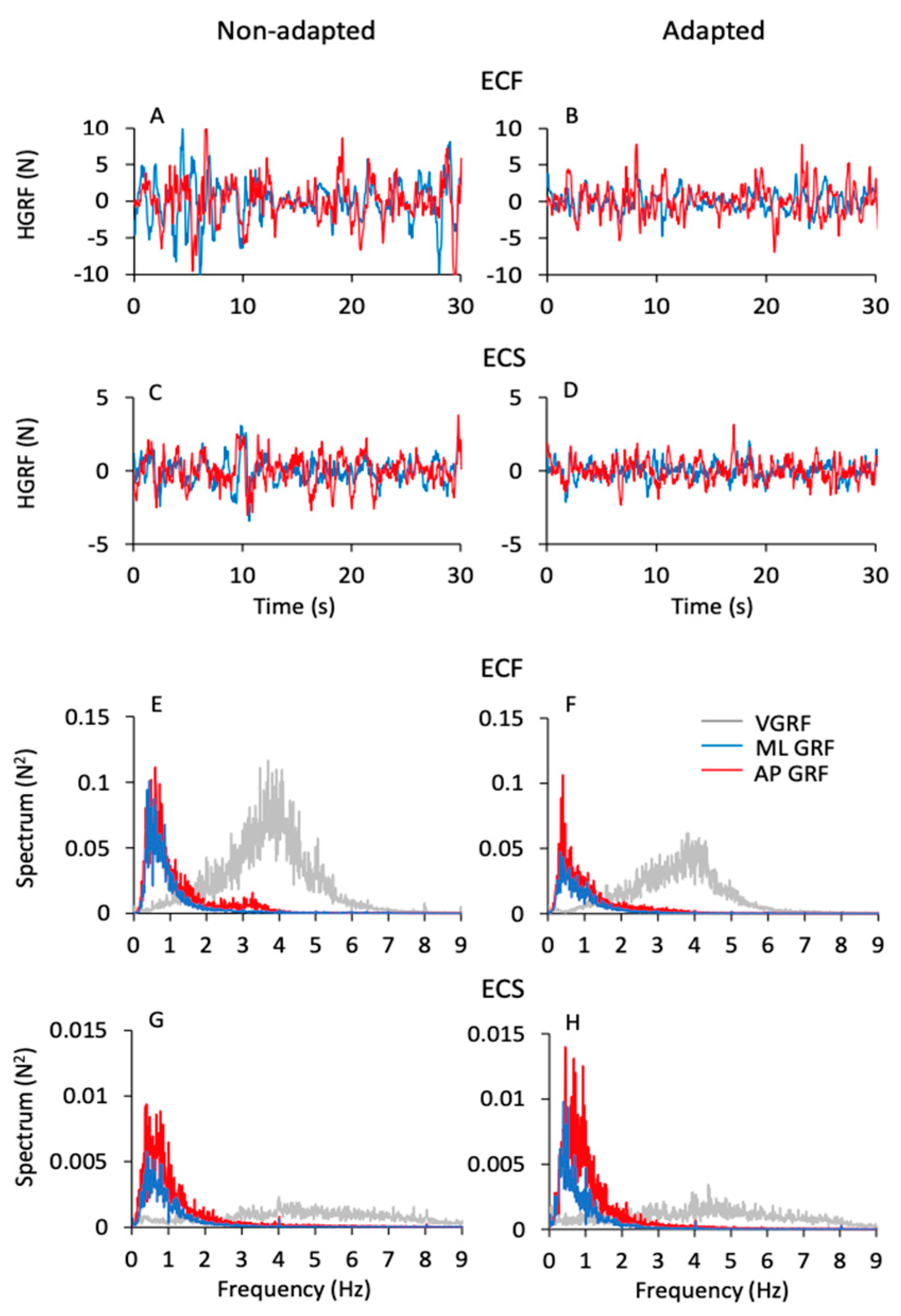
3.5. Frequency and Amplitude of the Horizontal Ground Reaction Force (HGRF) Do Not Match Those of the VGRF
3.6. The Frequency Spectra of the Horizontal Components of the HGRF (ML and AP) Do Not Reflect Those of the CoP Oscillations
3.7. The Cross-Correlation between the Oscillations of the HGRF and Those of the CoP
3.8. The HGRFs Have an Ill-Defined Relationship with the Excursions of the CoP
3.9. Vision, Support Surface and Adaptation Equally Affect HGRF and CoP Excursions
4. Discussion
4.1. Recap and Outline of the Main Findings
4.2. The VGRF Frequency Undergoes Minor Changes between Compliant and Hard Support
Body Weight Substantially Increases VGRF Oscillation Amplitude on Compliant Support but Minimally Affects VGRF Oscillation Frequency
4.3. The Different Spectrum Profiles of the Oscillations of VGRF, HGRF, and CoP
The CoP Frequencies and the Rambling–Trembling Hypothesis
4.4. The Cross-Correlation between HGRF and CoP Time Series
4.5. The HGRF Module Determines the CoP Position
4.6. The Effects of Adaptation Are Obvious Only in the Most Critical Standing Condition
4.6.1. Vision
4.6.2. Vestibular Input
4.6.3. Attentional Demands
4.7. Is the VGRF Rhythm Automatic or Voluntary?
4.8. The Postural Rhythm May Not Be Specific to Young Participants
4.9. Limitations
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Creath, R.; Kiemel, T.; Horak, F.; Peterka, R.; Jeka, J. A unified view of quiet and perturbed stance: Simultaneous co-existing excitable modes. Neurosci. Lett. 2005, 377, 75–80. [Google Scholar] [CrossRef]
- Günther, M.; Wagner, H. Dynamics of quiet human stance: Computer simulations of a triple inverted pendulum model. Comput. Methods Biomech. Biomed. Eng. 2016, 19, 819–834. [Google Scholar] [CrossRef]
- Sozzi, S.; Honeine, J.-L.; Do, M.-C.; Schieppati, M. Leg muscle activity during tandem stance and the control of body balance in the frontal plane. Clin. Neurophysiol. 2013, 124, 1175–1186. [Google Scholar] [CrossRef]
- Paillard, T. Relationship between Muscle Function, Muscle Typology and Postural Performance According to Different Postural Conditions in Young and Older Adults. Front. Physiol. 2017, 8, 585. [Google Scholar] [CrossRef] [Green Version]
- Mierau, A.; Pester, B.; Hülsdünker, T.; Schiecke, K.; Strüder, H.K.; Witte, H. Cortical Correlates of Human Balance Control. Brain Topogr. 2017, 30, 434–446. [Google Scholar] [CrossRef] [Green Version]
- Windhorst, U. Muscle proprioceptive feedback and spinal networks. Brain Res. Bull. 2007, 73, 155–202. [Google Scholar] [CrossRef]
- De Nunzio, A.M.; Schieppati, M. Time to reconfigure balancing behaviour in man: Changing visual condition while riding a continuously moving platform. Exp. Brain Res. 2007, 178, 18–36. [Google Scholar] [CrossRef]
- Sozzi, S.; Monti, A.; De Nunzio, A.M.; Do, M.-C.; Schieppati, M. Sensori-motor integration during stance: Time adaptation of control mechanisms on adding or removing vision. Hum. Mov. Sci. 2011, 30, 172–189. [Google Scholar] [CrossRef]
- Honeine, J.-L.; Schieppati, M. Time-interval for integration of stabilizing haptic and visual information in subjects balancing under static and dynamic conditions. Front. Syst. Neurosci. 2014, 8, 190. [Google Scholar] [CrossRef] [Green Version]
- Sozzi, S.; Schieppati, M. Balance Adaptation While Standing on a Compliant Base Depends on the Current Sensory Condition in Healthy Young Adults. Front. Hum. Neurosci. 2022, 16, 839799. [Google Scholar] [CrossRef]
- Chiba, R.; Takakusaki, K.; Ota, J.; Yozu, A.; Haga, N. Human upright posture control models based on multisensory inputs; in fast and slow dynamics. Neurosci. Res. 2016, 104, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Forbes, P.A.; Chen, A.; Blouin, J.-S. Sensorimotor control of standing balance. Handb. Clin. Neurol. 2018, 159, 61–83. [Google Scholar] [CrossRef]
- Ivanenko, Y.; Gurfinkel, V.S. Human Postural Control. Front. Neurosci. 2018, 12, 171. [Google Scholar] [CrossRef]
- Felicetti, G.; Thoumie, P.; Do, M.; Schieppati, M. Cutaneous and muscular afferents from the foot and sensory fusion processing: Physiology and pathology in neuropathies. J. Peripher. Nerv. Syst. 2021, 26, 17–34. [Google Scholar] [CrossRef]
- Morasso, P.G.; Schieppati, M. Can Muscle Stiffness Alone Stabilize Upright Standing? J. Neurophysiol. 1999, 82, 1622–1626. [Google Scholar] [CrossRef] [Green Version]
- Rougier, P.-R. What insights can be gained when analysing the resultant centre of pressure trajectory? Neurophysiol. Clin. 2008, 38, 363–373. [Google Scholar] [CrossRef]
- Sozzi, S.; Ghai, S.; Schieppati, M. Incongruity of Geometric and Spectral Markers in the Assessment of Body Sway. Front. Neurol. 2022, 13, 929132. [Google Scholar] [CrossRef]
- Crétual, A. Which biomechanical models are currently used in standing posture analysis? Neurophysiol. Clin. 2015, 45, 285–295. [Google Scholar] [CrossRef]
- Shin, S.; Milosevic, M.; Chung, C.-M.; Lee, Y. Contractile properties of superficial skeletal muscle affect postural control in healthy young adults: A test of the rambling and trembling hypothesis. PLoS ONE 2019, 14, e0223850. [Google Scholar] [CrossRef] [Green Version]
- Sozzi, S.; Nardone, A.; Schieppati, M. Specific Posture-Stabilising Effects of Vision and Touch Are Revealed by Distinct Changes of Body Oscillation Frequencies. Front. Neurol. 2021, 12, 756984. [Google Scholar] [CrossRef]
- Kiemel, T.; Elahi, A.J.; Jeka, J.J.; Engelhart, D.; Pasma, J.H.; Schouten, A.C.; Aarts, R.G.K.M.; Meskers, C.G.M.; Maier, A.B.; van der Kooij, H.; et al. Identification of the Plant for Upright Stance in Humans: Multiple Movement Patterns From a Single Neural Strategy. J. Neurophysiol. 2008, 100, 3394–3406. [Google Scholar] [CrossRef] [Green Version]
- Murnaghan, C.; Horslen, B.; Inglis, J.; Carpenter, M. Exploratory behavior during stance persists with visual feedback. Neuroscience 2011, 195, 54–59. [Google Scholar] [CrossRef]
- Bonnet, C.T. Advantages and disadvantages of stiffness instructions when studying postural control. Gait Posture 2016, 46, 208–210. [Google Scholar] [CrossRef]
- Peterka, R.J. Sensory integration for human balance control. Handb. Clin. Neurol. 2018, 159, 27–42. [Google Scholar]
- Karlsson, A.; Frykberg, G. Correlations between force plate measures for assessment of balance. Clin. Biomech. 2000, 15, 365–369. [Google Scholar] [CrossRef]
- Önell, A. The vertical ground reaction force for analysis of balance? Gait Posture 2000, 12, 7–13. [Google Scholar] [CrossRef]
- Hong, C.-Y.; Guo, L.-Y.; Song, R.; Nagurka, M.L.; Sung, J.-L.; Yen, C.-W. Assessing postural stability via the correlation patterns of vertical ground reaction force components. Biomed. Eng. Online 2016, 15, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Pagnacco, G.; Heiss, D.G.; Oggero, E. Muscular contractions and their effect on the vertical ground reaction force during quiet stance—Part I: Hypothesis and experimental investigation. Biomed. Sci. Instrum. 2001, 37, 227–232. [Google Scholar]
- Pagnacco, G.; Heiss, D.G.; Oggero, E. Muscular contractions and their effect on the vertical ground reaction force during quiet stance—Part II: Mathematical model. Biomed. Sci. Instrum. 2001, 37, 233–238. [Google Scholar]
- Minamisawa, T.; Sawahata, H.; Takakura, K.; Yamaguchi, T. Characteristics of temporal fluctuation of the vertical ground reaction force during quiet stance in Parkinson’s disease. Gait Posture 2012, 35, 308–311. [Google Scholar] [CrossRef]
- Giacomozzi, C.; Martelli, F.; Lillia, M.; Fadda, A. Fourier analysis of vertical forces to integrate balance measurements. J. Foot Ankle Res. 2014, 7, A23. [Google Scholar] [CrossRef] [Green Version]
- Minamisawa, T.; Chiba, N.; Inoue, K.; Nakanowatari, T.; Suzuki, E. Analysis of Vertical Micro Acceleration While Standing Reveals Age-Related Changes. Geriatrics 2020, 5, 105. [Google Scholar] [CrossRef]
- Sozzi, S.; Do, M.-C.; Schieppati, M. Vertical ground reaction force oscillation during standing on hard and compliant surfaces: The “postural rhythm”. Front. Neurol. 2022, 13, 975752. [Google Scholar] [CrossRef]
- Agarwal, G.C.; Gottlieb, G.L. Oscillation of the human ankle joint in response to applied sinusoidal torque on the foot. J. Physiol. 1977, 268, 151–176. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Griffin, M. Dynamic response of the standing human body exposed to vertical vibration: Influence of posture and vibration magnitude. J. Sound Vib. 1998, 212, 85–107. [Google Scholar] [CrossRef]
- Kasagi, Y.; Shimoyama, I.; Yoshida, S.; Nakazawa, K.; Fukutake, T. Effect of Aging on the Vibration of Human Body in the Vertical Direction during Quiet Stance. Chiba Med. J. 2005, 3, 10–12. [Google Scholar]
- Minamisawa, T.; Chiba, N.; Suzuki, E. Association of bilateral lower limb coordination while standing with body sway control and aging. Somatosens. Mot. Res. 2021, 38, 294–302. [Google Scholar] [CrossRef]
- Oggero, E.; Carrick, F.R.; Pagnacco, G. Frequency content of standard posturographic measures—Biomed 2013. Biomed. Sci. Instrum. 2013, 49, 48–53. [Google Scholar]
- Hülsdünker, T.; Mierau, A.; Strüder, H.K. Higher Balance Task Demands are Associated with an Increase in Individual Alpha Peak Frequency. Front. Hum. Neurosci. 2016, 9, 695. [Google Scholar] [CrossRef] [Green Version]
- Boehm, W.L.; Nichols, K.M.; Gruben, K.G. Frequency-dependent contributions of sagittal-plane foot force to upright human standing. J. Biomech. 2019, 83, 305–309. [Google Scholar] [CrossRef]
- Krakauer, J.W.; Mazzoni, P. Human sensorimotor learning: Adaptation, skill, and beyond. Curr. Opin. Neurobiol. 2011, 21, 636–644. [Google Scholar] [CrossRef]
- Lin, C.-C.; Roche, J.L.; Steed, D.P.; Musolino, M.C.; Marchetti, G.F.; Furman, G.R.; Redfern, M.S.; Whitney, S.L. Test-retest reliability of postural stability on two different foam pads. J. Nat. Sci. 2015, 1, e43. [Google Scholar]
- Lajoie, Y.; Richer, N.; Jehu, D.; Polskaia, N.; Saunders, D. Letter to the Editor: On “Advantages and disadvantages of stiffness instructions when studying postural control” by C.T. Bonnet: You just can’t win: Advantages and disadvantages of the postural stability requirement. Gait Posture 2016, 46, 215–218. [Google Scholar] [CrossRef]
- Lee, I.-C.; Pacheco, M.M.; Newell, K.M. The precision demands of viewing distance modulate postural coordination and control. Hum. Mov. Sci. 2019, 66, 425–439. [Google Scholar] [CrossRef]
- Caron, O.; Faure, B.; Brenière, V. Estimating the center of gravity of the body on the basis of the center of pressure in standing posture. J. Biomech. 1997, 30, 1169–1171. [Google Scholar] [CrossRef]
- Ruhe, A.; Fejer, R.; Walker, B. The test–retest reliability of centre of pressure measures in bipedal static task conditions—A systematic review of the literature. Gait Posture 2010, 32, 436–445. [Google Scholar] [CrossRef] [Green Version]
- Słomka, K.; Juras, G.; Sobota, G.; Bacik, B. The reliability of a rambling–trembling analysis of center of pressure measures. Gait Posture 2013, 37, 210–213. [Google Scholar] [CrossRef]
- Carpenter, M.G.; Frank, J.S.; Winter, D.A.; Peysar, G.W. Sampling duration effects on centre of pressure summary measures. Gait Posture 2001, 13, 35–40. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.-G. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Shiozawa, K.; Lee, J.; Russo, M.; Sternad, D.; Hogan, N. Frequency-dependent force direction elucidates neural control of balance. J. Neuroeng. Rehabil. 2021, 18, 1–12. [Google Scholar] [CrossRef]
- Pagnacco, G.; Sorbello, D.; Oggero, E.; Morr, D.R.; Berme, N. Average power spectral density of physiological tremor in normal subjects. Biomed. Sci. Instrum. 1997, 34, 99–103. [Google Scholar]
- Krizková, M.; Hlavacka, F. Binaural monopolar galvanic vestibular stimulation reduces body sway during human stance. Physiol. Res. 1994, 43, 187–192. [Google Scholar]
- Horak, F.B.; Hlavacka, F. Somatosensory Loss Increases Vestibulospinal Sensitivity. J. Neurophysiol. 2001, 86, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Isableu, B.; Vuillerme, N. Differential integration of kinaesthetic signals to postural control. Exp. Brain Res. 2006, 174, 763–768. [Google Scholar] [CrossRef]
- Kelly, L.A.; Kuitunen, S.; Racinais, S.; Cresswell, A.G. Recruitment of the plantar intrinsic foot muscles with increasing postural demand. Clin. Biomech. 2012, 27, 46–51. [Google Scholar] [CrossRef]
- Anson, E.; Bigelow, R.T.; Swenor, B.; Deshpande, N.; Studenski, S.; Jeka, J.J.; Agrawal, Y. Loss of Peripheral Sensory Function Explains Much of the Increase in Postural Sway in Healthy Older Adults. Front. Aging Neurosci. 2017, 9, 202. [Google Scholar] [CrossRef] [Green Version]
- Anson, E.; Bigelow, R.T.; Studenski, S.; Deshpande, N.; Agrawal, Y. Failure on the Foam Eyes Closed Test of Standing Balance Associated With Reduced Semicircular Canal Function in Healthy Older Adults. Ear Hear. 2019, 40, 340–344. [Google Scholar] [CrossRef]
- Gatev, P.; Thomas, S.; Kepple, T.; Hallett, M. Feedforward ankle strategy of balance during quiet stance in adults. J. Physiol. 1999, 514, 915–928. [Google Scholar] [CrossRef]
- Schieppati, M.; Nardone, A.; Siliotto, R.; Grasso, M. Early and late stretch responses of human foot muscles induced by perturbation of stance. Exp. Brain Res. 1995, 105, 411–422. [Google Scholar] [CrossRef]
- Okai, L.A.; Kohn, A. Quantifying the Contributions of the Flexor Digitorum Brevis Muscle on Postural Stability. Mot. Control. 2015, 19, 161–172. [Google Scholar] [CrossRef]
- Fransson, P.-A.; Gomez, S.; Patel, M.; Johansson, L. Changes in multi-segmented body movements and EMG activity while standing on firm and foam support surfaces. Eur. J. Appl. Physiol. 2007, 101, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, M.; Fransson, P.A.; Johansson, R.; Magnusson, M. Foam posturography: Standing on foam is not equivalent to standing with decreased rapidly adapting mechanoreceptive sensation. Exp. Brain Res. 2011, 208, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Günther, M.; Grimmer, S.; Siebert, T.; Blickhan, R. All leg joints contribute to quiet human stance: A mechanical analysis. J. Biomech. 2009, 42, 2739–2746. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Chiang, J.-H. The significance of somatosensory stimulations to the human foot in the control of postural reflexes. Exp. Brain Res. 1997, 114, 163–169. [Google Scholar] [CrossRef]
- Mademli, L.; Mavridi, D.; Bohm, S.; Patikas, D.A.; Santuz, A.; Arampatzis, A. Standing on unstable surface challenges postural control of tracking tasks and modulates neuromuscular adjustments specific to task complexity. Sci. Rep. 2021, 11, 6122. [Google Scholar] [CrossRef]
- Bottaro, A.; Yasutake, Y.; Nomura, T.; Casadio, M.; Morasso, P. Bounded stability of the quiet standing posture: An intermittent control model. Hum. Mov. Sci. 2008, 27, 473–495. [Google Scholar] [CrossRef]
- Michimoto, K.; Suzuki, Y.; Kiyono, K.; Kobayashi, Y.; Morasso, P.; Nomura, T. Reinforcement learning for stabilizing an inverted pendulum naturally leads to intermittent feedback control as in human quiet standing. In Proceedings of the 2016 38th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Orlando, FL, USA, 18 October 2016; Volume 2016, pp. 37–40. [Google Scholar] [CrossRef]
- Tigrini, A.; Verdini, F.; Fioretti, S.; Mengarelli, A. Center of pressure plausibility for the double-link human stance model under the intermittent control paradigm. J. Biomech. 2021, 128, 110725. [Google Scholar] [CrossRef]
- Rakhra, S.K.; Singer, J.C. The effect of ageing on between-limb centre of pressure coordination in standing balance: Is there evidence for reactive control challenges among older adults? Hum. Mov. Sci. 2022, 86, 103019. [Google Scholar] [CrossRef]
- Gerber, E.D.; Nichols, P.; Giraldo, C.; Sidener, L.; Huang, C.-K.; Luchies, C.W. Rambling-trembling center-of-pressure decomposition reveals distinct sway responses to simulated somatosensory deficit. Gait Posture 2022, 91, 276–283. [Google Scholar] [CrossRef]
- Borel, L.; Ribot-Ciscar, E. Improving postural control by applying mechanical noise to ankle muscle tendons. Exp. Brain Res. 2016, 234, 2305–2314. [Google Scholar] [CrossRef] [Green Version]
- Bove, M.; Trompetto, C.; Abbruzzese, G.; Schieppati, M. The posture-related interaction between Ia-afferent and descending input on the spinal reflex excitability in humans. Neurosci. Lett. 2006, 397, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, G.R. Modulation of tendon tap reflex activation of soleus motor neurons with reduced stability tandem stance. Hum. Mov. Sci. 2019, 64, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Alizadehsaravi, L.; Bruijn, S.M.; Maas, H.; van Dieën, J.H. Modulation of soleus muscle H-reflexes and ankle muscle co-contraction with surface compliance during unipedal balancing in young and older adults. Exp. Brain Res. 2020, 238, 1371–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farenc, I.; Rougier, P.; Berger, L. The influence of gender and body characteristics on upright stance. Ann. Hum. Biol. 2003, 30, 279–294. [Google Scholar] [CrossRef]
- Hue, O.; Simoneau, M.; Marcotte, J.; Berrigan, F.; Doré, J.; Marceau, P.; Marceau, S.; Tremblay, A.; Teasdale, N. Body weight is a strong predictor of postural stability. Gait Posture 2007, 26, 32–38. [Google Scholar] [CrossRef]
- Zultowski, I.; Aruin, A. Carrying loads and postural sway in standing: The effect of load placement and magnitude. Work 2008, 30, 359–368. [Google Scholar]
- Mignardot, J.-B.; Olivier, I.; Promayon, E.; Nougier, V. Obesity Impact on the Attentional Cost for Controlling Posture. PLoS ONE 2010, 5, e14387. [Google Scholar] [CrossRef]
- Menegoni, F.; Tacchini, E.; Bigoni, M.; Vismara, L.; Priano, L.; Galli, M.; Capodaglio, P. Mechanisms underlying center of pressure displacements in obese subjects during quiet stance. J. Neuroeng. Rehabil. 2011, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Teasdale, N. Du maintien postural à l’exécution du mouvement: Le cas de l’obésité. Neurophysiol. Clin. 2011, 41, 199. [Google Scholar] [CrossRef]
- Cieślińska-Świder, J.M.; Błaszczyk, J.W. Posturographic characteristics of the standing posture and the effects of the treatment of obesity on obese young women. PLoS ONE 2019, 14, e0220962. [Google Scholar] [CrossRef] [Green Version]
- D’Hondt, E.; Deforche, B.; De Bourdeaudhuij, I.; Gentier, I.; Tanghe, A.; Shultz, S.; Lenoir, M. Postural balance under normal and altered sensory conditions in normal-weight and overweight children. Clin. Biomech. 2011, 26, 84–89. [Google Scholar] [CrossRef]
- Rugelj, D.; Sevšek, F. The effect of load mass and its placement on postural sway. Appl. Ergon. 2011, 42, 860–866. [Google Scholar] [CrossRef]
- Pau, M.; Kim, S.; Nussbaum, M.A. Does load carriage differentially alter postural sway in overweight vs. normal-weight schoolchildren? Gait Posture 2012, 35, 378–382. [Google Scholar] [CrossRef]
- Jeong, H.; Johnson, A.W.; Feland, J.B.; Petersen, S.R.; Staten, J.M.; Bruening, D.A. Added body mass alters plantar shear stresses, postural control, and gait kinetics: Implications for obesity. PLoS ONE 2021, 16, e0246605. [Google Scholar] [CrossRef]
- Stiles, R.N.; Randall, J.E. Mechanical factors in human tremor frequency. J. Appl. Physiol. 1967, 23, 324–330. [Google Scholar] [CrossRef]
- Elble, R.J.; Randall, J.E. Mechanistic components of normal hand tremor. Electroencephalogr. Clin. Neurophysiol. 1978, 44, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Raethjen, J.; Pawlas, F.; Lindemann, M.; Wenzelburger, R.; Deuschl, G. Determinants of physiologic tremor in a large normal population. Clin. Neurophysiol. 2000, 111, 1825–1837. [Google Scholar] [CrossRef] [PubMed]
- Takanokura, M.; Sakamoto, K. Neuromuscular control of physiological tremor during elastic load. Med. Sci. Monit. 2005, 11, CR143–CR152. [Google Scholar] [PubMed]
- Deuschl, G.; Becktepe, J.S.; Dirkx, M.; Haubenberger, D.; Hassan, A.; Helmich, R.C.; Muthuraman, M.; Panyakaew, P.; Schwingenschuh, P.; Zeuner, K.E.; et al. The clinical and electrophysiological investigation of tremor. Clin. Neurophysiol. 2022, 136, 93–129. [Google Scholar] [CrossRef] [PubMed]
- Lakie, M.; Vernooij, C.A.; Osler, C.J.; Stevenson, A.T.; Scott, J.P.R.; Reynolds, R.F. Increased gravitational force reveals the mechanical, resonant nature of physiological tremor. J. Physiol. 2015, 593, 4411–4422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bach, T.; Chapman, A.; Calvert, T. Mechanical resonance of the human body during voluntary oscillations about the ankle joint. J. Biomech. 1983, 16, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Kiiski, J.; Heinonen, A.; Järvinen, T.L.; Kannus, P.; Sievänen, H. Transmission of vertical whole body vibration to the human body. Journal of bone and mineral research. 2008, 23, 1318–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subashi, G.H.; Matsumoto, Y.; Griffin, M.J. Modelling resonances of the standing body exposed to vertical whole-body vibration: Effects of pos-ture. Journal of Sound and Vibration. 2008, 317, 400–418. [Google Scholar] [CrossRef]
- Masani, K.; Vette, A.H.; Kawashima, N.; Popovic, M.R. Neuromusculoskeletal Torque-Generation Process Has a Large Destabilizing Effect on the Control Mechanism of Quiet Standing. J. Neurophysiol. 2008, 100, 1465–1475. [Google Scholar] [CrossRef] [Green Version]
- Vette, A.H.; Masani, K.; Nakazawa, K.; Popovic, M.R. Neural-Mechanical Feedback Control Scheme Generates Physiological Ankle Torque Fluctuation During Quiet Stance. IEEE Trans. Neural Syst. Rehabilitation Eng. 2010, 18, 86–95. [Google Scholar] [CrossRef]
- Day, B.L.; Steiger, M.J.; Thompson, P.D.; Marsden, C.D. Effect of vision and stance width on human body motion when standing: Implications for afferent control of lateral sway. J. Physiol. 1993, 469, 479–499. [Google Scholar] [CrossRef]
- Winter, D.A.; Prince, F.; Frank, J.S.; Powell, C.; Zabjek, K.F. Unified theory regarding A/P and M/L balance in quiet stance. J. Neurophysiol. 1996, 75, 2334–2343. [Google Scholar] [CrossRef]
- Carpenter, M.G.; Allum, J.H.J.; Honegger, F. Directional sensitivity of stretch reflexes and balance corrections for normal subjects in the roll and pitch planes. Exp. Brain Res. 1999, 129, 93–113. [Google Scholar] [CrossRef]
- Bogaart, M.V.D.; Bruijn, S.M.; Spildooren, J.; van Dieën, J.H.; Meyns, P. Effects of age and surface instability on the control of the center of mass. Hum. Mov. Sci. 2022, 82, 102930. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Fransson, P.; Lush, D.; Gomez, S. The effect of foam surface properties on postural stability assessment while standing. Gait Posture 2008, 28, 649–656. [Google Scholar] [CrossRef] [PubMed]
- van Dieën, J.H.; van Leeuwen, M.; Faber, G.S. Learning to balance on one leg: Motor strategy and sensory weighting. J. Neurophysiol. 2015, 114, 2967–2982. [Google Scholar] [CrossRef] [PubMed]
- Ivanenko, Y.P.; Poppele, R.E.; Lacquaniti, F. Motor Control Programs and Walking. Neurosci. 2006, 12, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.P.; Whitney, R.J. Postural movements during normal standing in man. J. Anat. 1959, 93 Pt 4, 524–539. [Google Scholar]
- Hufschmidt, A.; Dichgans, J.; Mauritz, K.-H.; Hufschmidt, M. Some methods and parameters of body sway quantification and their neurological applications. Arch. Psychiatr. Nervenkr. 1980, 228, 135–150. [Google Scholar] [CrossRef] [PubMed]
- Schumann, T.; Redfern, M.S.; Furman, J.M.; El-Jaroudi, A.; Chaparro, L.F. Time-frequency analysis of postural sway. J. Biomech. 1995, 28, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Dakin, C.J.; Luu, B.L.; Doel, K.v.D.; Inglis, J.T.; Blouin, J.-S. Frequency-Specific Modulation of Vestibular-Evoked Sway Responses in Humans. J. Neurophysiol. 2010, 103, 1048–1056. [Google Scholar] [CrossRef]
- Domínguez, M.O.; Magro, J.B.; Gaona, J.R.; Pastor, J.B. Spectrum analysis in postural strategy on static tests in a healthy population. Acta Otorrinolaringol. Esp. 2013, 64, 124–132. [Google Scholar] [CrossRef]
- Warnica, M.J.; Weaver, T.B.; Prentice, S.D.; Laing, A.C. The influence of ankle muscle activation on postural sway during quiet stance. Gait Posture 2014, 39, 1115–1121. [Google Scholar] [CrossRef]
- Yamamoto, T.; Smith, C.E.; Suzuki, Y.; Kiyono, K.; Tanahashi, T.; Sakoda, S.; Morasso, P.; Nomura, T. Universal and individual characteristics of postural sway during quiet standing in healthy young adults. Physiol. Rep. 2015, 3, e12329. [Google Scholar] [CrossRef]
- Hsu, W.L.; Scholz, J.P.; Schöner, G.; Jeka, J.J.; Kiemel, T. Control and estimation of posture during quiet stance depends on multijoint coordination. J. Neurophysiol. 2007, 97, 3024–3035. [Google Scholar] [CrossRef] [Green Version]
- Pinter, I.J.; van Swigchem, R.; van Soest, A.J.K.; Rozendaal, L.A. The Dynamics of Postural Sway Cannot Be Captured Using a One-Segment Inverted Pendulum Model: A PCA on Segment Rotations During Unperturbed Stance. J. Neurophysiol. 2008, 100, 3197–3208. [Google Scholar] [CrossRef] [Green Version]
- Tsianos, G.A.; Loeb, G.E. Muscle and Limb Mechanics. Compr. Physiol. 2017, 7, 429–462. [Google Scholar] [CrossRef]
- King, D.L.; Zatsiorsky, V.M. Extracting gravity line displacement from stabilographic recordings. Gait Posture 1997, 6, 27–38. [Google Scholar] [CrossRef]
- Zatsiorsky, V.M.; Duarte, M. Rambling and Trembling in Quiet Standing. Mot. Control. 2000, 4, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Solnik, S.; Pazin, N.; Coelho, C.J.; Rosenbaum, D.A.; Zatsiorsky, V.M.; Latash, M.L. Postural sway and perceived comfort in pointing tasks. Neurosci. Lett. 2014, 569, 18–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morasso, P.G.; Spada, G.; Capra, R. Computing the COM from the COP in postural sway movements. Hum. Mov. Sci. 1999, 18, 759–767. [Google Scholar] [CrossRef]
- Yu, E.; Abe, M.; Masani, K.; Kawashima, N.; Eto, F.; Haga, N.; Nakazawa, K. Evaluation of postural control in quiet standing using centre of mass acceleration: Comparison among the young, the elderly, and people with stroke. Arch. Phys. Med. Rehabil. 2008, 89, 1133–1139. [Google Scholar] [CrossRef]
- Elias, L.A.; Watanabe, R.N.; Kohn, A. Spinal Mechanisms May Provide a Combination of Intermittent and Continuous Control of Human Posture: Predictions from a Biologically Based Neuromusculoskeletal Model. PLOS Comput. Biol. 2014, 10, e1003944. [Google Scholar] [CrossRef]
- Ko, J.-H.; Challis, J.H.; Newell, K.M. Transition of COM–COP relative phase in a dynamic balance task. Hum. Mov. Sci. 2014, 38, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zatsiorsky, V.M.; King, D.L. An algorithm for determining gravity line location from posturographic recordings. J. Biomech. 1998, 31, 161–164. [Google Scholar] [CrossRef]
- Lafond, D.; Duarte, M.; Prince, F. Comparison of three methods to estimate the center of mass during balance assessment. J. Biomech. 2004, 37, 1421–1426. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kiemel, T.; Jeka, J. The influence of sensory information on two-component coordination during quiet stance. Gait Posture 2007, 26, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masani, K.; Popovic, M.R.; Nakazawa, K.; Kouzaki, M.; Nozaki, D. Importance of body sway velocity information in controlling ankle extensor ac-tivities during quiet stance. J. Neurophysiol. 2003, 90, 3774–3782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suissa, D.; Günther, M.; Shapiro, A.; Melzer, I.; Schmitt, S. On Laterally Perturbed Human Stance: Experiment, Model, and Control. Appl. Bionics Biomech. 2018, 2018, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Morasso, P. Centre of pressure versus centre of mass stabilization strategies: The tightrope balancing case. R. Soc. Open Sci. 2020, 7, 200111. [Google Scholar] [CrossRef]
- Schieppati, M.; Hugon, M.; Grasso, M.; Nardone, A.; Galante, M. The limits of equilibrium in young and elderly normal subjects and in parkinsonians. Electroencephalogr. Clin. Neurophysiol. Evoked Potentials Sect. 1994, 93, 286–298. [Google Scholar] [CrossRef]
- Castro, P.; Kaski, D.; Schieppati, M.; Furman, M.; Arshad, Q.; Bronstein, A. Subjective stability perception is related to postural anxiety in older subjects. Gait Posture 2019, 68, 538–544. [Google Scholar] [CrossRef]
- Albert, S.T.; Jang, J.; Modchalingam, S.; Hart, B.M.; Henriques, D.; Lerner, G.; Della-Maggiore, V.; Haith, A.M.; Krakauer, J.W.; Shadmehr, R. Competition between parallel sensorimotor learning systems. Elife 2022, 11, e65361. [Google Scholar] [CrossRef]
- Pipereit, K.; Bock, O.; Vercher, J.-L. The contribution of proprioceptive feedback to sensorimotor adaptation. Exp. Brain Res. 2006, 174, 45–52. [Google Scholar] [CrossRef]
- Allison, L.K.; Kiemel, T.; Jeka, J.J. Sensory-Challenge Balance Exercises Improve Multisensory Reweighting in Fall-Prone Older Adults. J. Neurol. Phys. Ther. 2018, 42, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Pagnacco, G.; Carrick, F.R.; Pascolo, P.B.; Rossi, R.; Oggero, E. Learning effect of standing on foam during posturographic testing: Preliminary findings. Biomed. Sci. Instrum. 2012, 48, 332–339. [Google Scholar] [PubMed]
- Assländer, L.; Gruber, M.; Giboin, L.-S. Reductions in body sway responses to a rhythmic support surface tilt perturbation can be caused by other mechanisms than prediction. Exp. Brain Res. 2020, 238, 465–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, M.; Sozzi, S. Temporal features of postural adaptation strategy to prolonged and repeatable balance perturbation. Neurosci. Lett. 2016, 628, 110–115. [Google Scholar] [CrossRef]
- Wang, G.; Yang, Y.; Wang, J.; Hao, Z.; Luo, X.; Liu, J. Dynamic changes of brain networks during standing balance control under visual conflict. Front. Neurosci. 2022, 16, 1003996. [Google Scholar] [CrossRef] [PubMed]
- Taube, W.; Gruber, M.; Gollhofer, A. Spinal and supraspinal adaptations associated with balance training and their functional relevance. Acta Physiol. 2008, 193, 101–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shadmehr, R.; Smith, M.A.; Krakauer, J.W. Error Correction, Sensory Prediction, and Adaptation in Motor Control. Annu. Rev. Neurosci. 2010, 33, 89–108. [Google Scholar] [CrossRef] [Green Version]
- Mierau, A.; Hülsdünker, T.; Strüder, H.K. Changes in cortical activity associated with adaptive behavior during repeated balance perturbation of unpredictable timing. Front. Behav. Neurosci. 2015, 9, 272. [Google Scholar] [CrossRef] [Green Version]
- Richmond, S.B.; Otto, G.; Dames, K.D. Characterization of trial duration in traditional and emerging postural control measures. J. Biomech. 2023, 147, 111438. [Google Scholar] [CrossRef]
- Sozzi, S.; Nardone, A.; Schieppati, M. Adaptation of balancing behaviour during continuous perturbations of stance. Supra-postural visual tasks and platform translation frequency modulate adaptation rate. PLoS ONE 2020, 15, e0236702. [Google Scholar] [CrossRef]
- Taylor, A.N.W.; Low, D.C.; Walsh, G.S.; Holt, N. The impact of anxiety on postural control: CO2 challenge model. Psychophysiology 2023, 60, e14192. [Google Scholar] [CrossRef]
- Trimble, M.H.; Koceja, D.M. Modulation of the Triceps Surae H-Reflex with Training. Int. J. Neurosci. 1994, 76, 293–303. [Google Scholar] [CrossRef]
- Houdijk, H.; Brown, S.E.; van Dieën, J.H. Relation between postural sway magnitude and metabolic energy cost during upright standing on a compliant surface. J. Appl. Physiol. 2015, 119, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Maurer, C.; Mergner, T.; Bolha, B.; Hlavacka, F. Human balance control during cutaneous stimulation of the plantar soles. Neurosci. Lett. 2001, 302, 45–48. [Google Scholar] [CrossRef]
- Viseux, F.J.F.; Lemaire, A.; Barbier, F.; Charpentier, P.; Leteneur, S.; Villeneuve, P. How can the stimulation of plantar cutaneous receptors improve postural control? Review and clinical commentary. Neurophysiol. Clin. 2019, 49, 263–268. [Google Scholar] [CrossRef]
- Fabre, M.; Antoine, M.; Robitaille, M.G.; Ribot-Ciscar, E.; Ackerley, R.; Aimonetti, J.-M.; Chavet, P.; Blouin, J.; Simoneau, M.; Mouchnino, L. Large Postural Sways Prevent Foot Tactile Information From Fading: Neurophysiological Evidence. Cereb. Cortex Commun. 2020, 2, tgaa094. [Google Scholar] [CrossRef] [PubMed]
- Macefield, V.G. The roles of mechanoreceptors in muscle and skin in human proprioception. Curr. Opin. Physiol. 2021, 21, 48–56. [Google Scholar] [CrossRef]
- Sakanaka, T.E.; Lakie, M.; Reynolds, R.F. Idiosyncratic Characteristics of Postural Sway in Normal and Perturbed Standing. Front. Hum. Neurosci. 2021, 15, 660470. [Google Scholar] [CrossRef]
- Babadi, S.; Vahdat, S.; Milner, T.E. Neural Substrates of Muscle Co-contraction during Dynamic Motor Adaptation. J. Neurosci. 2021, 41, 5667–5676. [Google Scholar] [CrossRef] [PubMed]
- Vette, A.H.; Sayenko, D.G.; Jones, M.; Abe, M.O.; Nakazawa, K.; Masani, K. Ankle muscle co-contractions during quiet standing are associated with decreased postural steadiness in the elderly. Gait Posture 2017, 55, 31–36. [Google Scholar] [CrossRef]
- Horlings, C.G.; van Engelen, B.G.; Allum, J.H.; Bloem, B.R. A weak balance: The contribution of muscle weakness to postural instability and falls. Nat. Clin. Pr. Neurol. 2008, 4, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Wright, W.G. Tonic Neuromuscular Processing Affects Postural Adaptation Differently in Aging and Parkinson’s Disease. Front. Neurol. 2019, 9, 1130. [Google Scholar] [CrossRef] [Green Version]
- Schmid, M.; Nardone, A.; De Nunzio, A.M.; Schieppati, M. Equilibrium during static and dynamic tasks in blind subjects: No evidence of cross-modal plasticity. Brain 2007, 130 Pt 8, 2097–2107. [Google Scholar] [CrossRef]
- Schieppati, M.; Schmid, M.; Sozzi, S. Rapid processing of haptic cues for postural control in blind subjects. Clin. Neurophysiol. 2014, 125, 1427–1439. [Google Scholar] [CrossRef]
- Kollegger, H.; Wöber, C.; Baumgartner, C.; Deecke, L. Stabilizing and Destabilizing Effects of Vision and Foot Position on Body Sway of Healthy Young Subjects: A Posturographic Study. Eur. Neurol. 1989, 29, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-W.; Kwon, Y.; Jeon, H.-M.; Bang, M.-J.; Jun, J.-H.; Eom, G.-M.; Lim, D.-H. Feet distance and static postural balance: Implication on the role of natural stance. Bio-Medical Mater. Eng. 2014, 24, 2681–2688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lord, S.R.; Clark, R.D.; Webster, I.W. Visual Acuity and Contrast Sensitivity in Relation to Falls in an Elderly Population. Age Ageing 1991, 20, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Nordahl, S.H.; Aasen, T.; Dyrkorn, B.M.; Eidsvik, S.; Molvaer, O.I. Static stabilometry and repeated testing in a normal population. Aviat Space En-viron Med. 2000, 71, 889–893. [Google Scholar]
- Wagner, A.R.; Akinsola, O.; Chaudhari, A.M.W.; Bigelow, K.E.; Merfeld, D.M. Measuring Vestibular Contributions to Age-Related Balance Impairment: A Review. Front. Neurol. 2021, 12, 635305. [Google Scholar] [CrossRef]
- Karmali, F.; Goodworth, A.D.; Valko, Y.; Leeder, T.; Peterka, R.J.; Merfeld, D.M. The role of vestibular cues in postural sway. J. Neurophysiol. 2021, 125, 672–686. [Google Scholar] [CrossRef]
- Welgampola, M.S.; Colebatch, J.G. Vestibulospinal reflexes: Quantitative effects of sensory feedback and postural task. Exp. Brain Res. 2001, 139, 345–353. [Google Scholar] [CrossRef]
- Naranjo, E.N.; Allum, J.H.J.; Inglis, J.T.; Carpenter, M.G. Increased gain of vestibulospinal potentials evoked in neck and leg muscles when standing under height-induced postural threat. Neuroscience 2015, 293, 45–54. [Google Scholar] [CrossRef]
- Cullen, K.E.; Zobeiri, O.A. Proprioception and the predictive sensing of active self-motion. Curr. Opin. Physiol. 2021, 20, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Diener, H.C.; Dichgans, J.; Guschlbauer, B.; Bacher, M. Role of visual and static vestibular influences on dynamic posture control. Hum. Neuro-Biol. 1986, 5, 105–113. [Google Scholar]
- Wiesmeier, I.K.; Dalin, D.; Emaurer, C. Elderly Use Proprioception Rather than Visual and Vestibular Cues for Postural Motor Control. Front. Aging Neurosci. 2015, 7, 97. [Google Scholar] [CrossRef] [PubMed]
- Lacour, M.; Barthelemy, J.; Borel, L.; Magnan, J.; Xerri, C.; Chays, A.; Ouaknine, M. Sensory strategies in human postural control before and after unilateral vestibular neurotomy. Exp. Brain Res. 1997, 115, 300–310. [Google Scholar] [CrossRef]
- Kabbaligere, R.; Lee, B.-C.; Layne, C.S. Balancing sensory inputs: Sensory reweighting of ankle proprioception and vision during a bipedal posture task. Gait Posture 2017, 52, 244–250. [Google Scholar] [CrossRef]
- Shafer, R.L.; Solomon, E.M.; Newell, K.M.; Lewis, M.H.; Bodfish, J.W. Visual feedback during motor performance is associated with increased complexity and adaptability of motor and neural output. Behav. Brain Res. 2019, 376, 112214. [Google Scholar] [CrossRef]
- Ohba, H.; Nakagawa, K.; Iida, K.; Yuge, L. Primary somatosensory cortex sensitivity may increase upon completion of a motor task. Neurosci. Lett. 2023, 801, 137160. [Google Scholar] [CrossRef]
- Fransson, P.-A.; Hafstrom, A.; Karlberg, M.; Magnusson, M.; Tjader, A.; Johansson, R. Postural control adaptation during galvanic vestibular and vibratory proprioceptive stimulation. IEEE Trans. Biomed. Eng. 2003, 50, 1310–1319. [Google Scholar] [CrossRef]
- Jankowska, E. On the distribution of information from muscle spindles in the spinal cord; how much does it depend on random factors? J. Anat. 2015, 227, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Edgerton, V.R.; Gad, P. Spinal automaticity of movement control and its role in recovering function after spinal injury. Expert Rev. Neurother. 2022, 22, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Doumas, M.; Valkanidis, T.C.; Hatzitaki, V. Putting proprioception for balance to the test: Contrasting and combining sway referencing and tendon vibration. Gait Posture 2019, 67, 201–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaback, M.; Adkin, A.L.; Chua, R.; Inglis, J.T.; Carpenter, M.G. Facilitation and Habituation of Cortical and Subcortical Control of Standing Balance Following Repeated Exposure to a Height-related Postural Threat. Neuroscience 2022, 487, 8–25. [Google Scholar] [CrossRef] [PubMed]
- Lajoie, Y.; Teasdale, N.; Bard, C.; Fleury, M. Attentional demands for static and dynamic equilibrium. Exp. Brain Res. 1993, 97, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, J.; Kreuzpointner, M.-A.; Krewer, C.; Bardins, S.; Schepermann, A.; Koenig, E.; Müller, F.; Jahn, K. The subjective postural vertical in standing: Reliability and normative data for healthy subjects. Atten. Percept. Psychophys 2015, 77, 953–960. [Google Scholar] [CrossRef] [Green Version]
- Ghai, S.; Ghai, I.; Effenberg, A.O. Effects of dual tasks and dual-task training on postural stability: A systematic review and meta-analysis. Clin. Interv. Aging 2017, ume 12, 557–577. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, D.; Belur, P.; Myers, P.S.; Earhart, G.M.; Rawson, K.S. The impact of age, surface characteristics, and dual-tasking on postural sway. Arch. Gerontol. Geriatr. 2020, 87, 103973. [Google Scholar] [CrossRef]
- Nedelkou, A.; Hatzitaki, V.; Chatzinikolaou, K.; Grouios, G. Does somatosensory feedback from the plantar foot sole contribute to verticality perception? Somatosens. Mot. Res. 2021, 38, 214–222. [Google Scholar] [CrossRef]
- Rubega, M.; Formaggio, E.; Di Marco, R.; Bertuccelli, M.; Tortora, S.; Menegatti, E.; Cattelan, M.; Bonato, P.; Masiero, S.; Del Felice, A. Cortical correlates in upright dynamic and static balance in the elderly. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Vuillerme, N.; Nougier, V. Attentional demand for regulating postural sway: The effect of expertise in gymnastics. Brain Res. Bull. 2004, 63, 161–165. [Google Scholar] [CrossRef]
- Maki, B.E.; McIlroy, W.E. Influence of arousal and attention on the control of postural sway. J. Vestib. Res. 1996, 6, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Fraizer, E.; Mitra, S. Methodological and interpretive issues in posture-cognition dual-tasking in upright stance. Gait Posture 2008, 27, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Li, D.K.; Mak, T.C.; Wong, T.W. Does an Externally Focused Dual-task Mitigate Real-time Conscious Postural Control in Older Adults? Exp. Aging Res. 2022, 48, 295–310. [Google Scholar] [CrossRef]
- St-Amant, G.; Rahman, T.; Polskaia, N.; Fraser, S.; Lajoie, Y. Unveilling the cerebral and sensory contributions to automatic postural control during dual-task standing. Hum. Mov. Sci. 2020, 70, 102587. [Google Scholar] [CrossRef]
- Potvin-Desrochers, A.; Richer, N.; Lajoie, Y. Cognitive tasks promote automatization of postural control in young and older adults. Gait Posture 2017, 57, 40–45. [Google Scholar] [CrossRef]
- McAuley, J.H.; Britton, T.C.; Rothwell, J.C.; Findley, L.J.; Marsden, C.D. The timing of primary orthostatic tremor bursts has a task-specific plasticity. Brain 2000, 123 Pt 2, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Gerschlager, W.; Brown, P. Orthostatic tremor—A review. Handb. Clin. Neurol. 2011, 100, 457–462. [Google Scholar] [CrossRef]
- Feil, K.; Böttcher, N.; Guri, F.; Krafczyk, S.; Schöberl, F.; Zwergal, A.; Strupp, M. Long-term course of orthostatic tremor in serial posturographic measurement. Park. Relat. Disord. 2015, 21, 905–910. [Google Scholar] [CrossRef]
- Hassan, A.; Caviness, J. Slow Orthostatic Tremor: Review of the Current Evidence. Tremor Other Hyperkinetic Mov. 2019, 9, 1–17. [Google Scholar] [CrossRef]
- Schöberl, F.; Feil, K.; Xiong, G.; Bartenstein, P.; la Fougère, C.; Jahn, K.; Brandt, T.; Strupp, M.; Dieterich, M.; Zwergal, A. Pathological ponto-cerebello-thalamo-cortical activations in primary orthostatic tremor during lying and stance. Brain 2017, 140, 83–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leu-Semenescu, S.; Roze, E.; Vidailhet, M.; Legrand, A.-P.; Trocello, J.-M.; Cochen, V.; Sangla, S.; Apartis, E. Myoclonus or tremor in orthostatism: An under-recognized cause of unsteadiness in Parkinson’s disease. Mov. Disord. 2007, 22, 2063–2069. [Google Scholar] [CrossRef]
- Erro, R.; Bhatia, K.P.; Cordivari, C. Shaking on Standing: A Critical Review. Mov. Disord. Clin. Pr. 2014, 1, 173–179. [Google Scholar] [CrossRef]
- Rezvanian, S.; Lockhart, T.; Frames, C.; Soangra, R.; Lieberman, A. Motor Subtypes of Parkinson’s Disease Can Be Identified by Frequency Component of Postural Stability. Sensors 2018, 18, 1102. [Google Scholar] [CrossRef] [Green Version]
- Ouchi, Y.; Okada, H.; Yoshikawa, E.; Nobezawa, S.; Futatsubashi, M. Brain activation during maintenance of standing postures in humans. Brain 1999, 122, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, J.; Wu, G.; Kelly, K. Evidence for beta corticomuscular coherence during human standing balance: Effects of stance width, vision, and support surface. Neuroscience 2015, 298, 1–11. [Google Scholar] [CrossRef]
- Dijkstra, B.W.; Bekkers, E.M.; Gilat, M.; de Rond, V.; Hardwick, R.M.; Nieuwboer, A. Functional neuroimaging of human postural control: A systematic review with meta-analysis. Neurosci. Biobehav. Rev. 2020, 115, 351–362. [Google Scholar] [CrossRef]
- Balestrucci, P.; Daprati, E.; Lacquaniti, F.; Maffei, V. Effects of visual motion consistent or inconsistent with gravity on postural sway. Exp. Brain Res. 2017, 235, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Solis-Escalante, T.; Stokkermans, M.; Cohen, M.X.; Weerdesteyn, V. Cortical responses to whole-body balance perturbations index perturbation magnitude and predict reactive stepping behavior. Eur. J. Neurosci. 2021, 54, 8120–8138. [Google Scholar] [CrossRef] [PubMed]
- Uiga, L.; Capio, C.M.; Ryu, D.; Wilson, M.R.; Masters, R.S. The role of conscious control in maintaining stable posture. Hum. Mov. Sci. 2018, 57, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.A.; Patton, J.L. Learned changes in the complexity of movement organization during multijoint, standing pulls. Biol. Cybern. 1997, 77, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Henry, S.M.; Fung, J.; Horak, F.B. EMG Responses to Maintain Stance During Multidirectional Surface Translations. J. Neurophysiol. 1998, 80, 1939–1950. [Google Scholar] [CrossRef] [Green Version]
- Varghese, J.P.; Beyer, K.B.; Williams, L.; Miyasike-Dasilva, V.; McIlroy, W.E. Standing still: Is there a role for the cortex? Neurosci. Lett. 2015, 590, 18–23. [Google Scholar] [CrossRef]
- Amiri, P.; Kearney, R.E. Identification of Central and Stretch Reflex Contributions to Human Postural Control. IEEE Trans. Neural Syst. Rehabilitation Eng. 2021, 29, 497–507. [Google Scholar] [CrossRef]
- Cherif, A.; Zenzeri, J.; Loram, I. What is the contribution of voluntary and reflex processes to sensorimotor control of balance? Front. Bioeng. Biotechnol. 2022, 10, 973716. [Google Scholar] [CrossRef]
- Enomoto, H.; Ugawa, Y.; Hanajima, R.; Yuasa, K.; Mochizuki, H.; Terao, Y.; Shiio, Y.; Furubayashi, T.; Iwata, N.; Kanazawa, I. Decreased sensory cortical excitability after 1 Hz rTMS over the ipsilateral primary motor cortex. Clin. Neurophysiol. 2001, 112, 2154–2158. [Google Scholar] [CrossRef] [Green Version]
- Bolton, D.A.; Brown, K.E.; McIlroy, W.E.; Staines, W.R. Transient inhibition of the dorsolateral prefrontal cortex disrupts somatosensory modulation during standing balance as measured by electroencephalography. Neuroreport 2012, 23, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Kaulmann, D.; Hermsdörfer, J.; Johannsen, L. Disruption of right posterior parietal cortex by continuous Theta Burst Stimulation alters the control of body balance in quiet stance. Eur. J. Neurosci. 2017, 45, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Solis-Escalante, T.; De Kam, D.; Weerdesteyn, V. Classification of Rhythmic Cortical Activity Elicited by Whole-Body Balance Perturbations Suggests the Cortical Representation of Direction-Specific Changes in Postural Stability. IEEE Trans. Neural Syst. Rehabilitation Eng. 2020, 28, 2566–2574. [Google Scholar] [CrossRef] [PubMed]
- Hülsdünker, T.; Mierau, A.; Neeb, C.; Kleinöder, H.; Strüder, H. Cortical processes associated with continuous balance control as revealed by EEG spectral power. Neurosci. Lett. 2015, 592, 2154–2158. [Google Scholar] [CrossRef]
- Peterson, S.M.; Ferris, D.P. Differentiation in Theta and Beta Electrocortical Activity between Visual and Physical Perturbations to Walking and Standing Balance. Eneuro 2018, 5, ENEURO.0207-18.2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büchel, D.; Lehmann, T.; Ullrich, S.; Cockcroft, J.; Louw, Q.; Baumeister, J. Stance leg and surface stability modulate cortical activity during human single leg stance. Exp. Brain Res. 2021, 239, 1193–1202. [Google Scholar] [CrossRef]
- Bosch, T.J.; Kammermeier, S.; Groth, C.; Leedom, M.; Hanson, E.K.; Berg-Poppe, P.; Singh, A. Cortical and Cerebellar Oscillatory Responses to Postural Instability in Parkinson’s Disease. Front. Neurol. 2021, 12, 752271. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Tsai, Y.-Y.; Chang, G.-C.; Hwang, I.-S. Cortical reorganization to improve dynamic balance control with error amplification feedback. J. Neuroeng. Rehabilitation 2022, 19, 3. [Google Scholar] [CrossRef] [PubMed]
- Sherman, D.A.; Lehmann, T.; Baumeister, J.; Grooms, D.R.; Norte, G.E. Somatosensory perturbations influence cortical activity associated with single-limb balance performance. Exp. Brain Res. 2022, 240, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Stokkermans, M.; Solis-Escalante, T.; Cohen, M.X.; Weerdesteyn, V. Midfrontal theta dynamics index the monitoring of postural stability. Cereb. Cortex 2022, 33, 3454–3466. [Google Scholar] [CrossRef]
- Barone, J.; Rossiter, H.E. Understanding the Role of Sensorimotor Beta Oscillations. Front. Syst. Neurosci. 2021, 15, 655886. [Google Scholar] [CrossRef]
- Hu, Y.; Petruzzello, S.J.; Hernandez, M.E. Beta cortical oscillatory activities and their relationship to postural control in a standing balance de-manding test: Influence of aging. Front. Aging Neurosci. 2023, 15, 285. [Google Scholar] [CrossRef]
- Kouzaki, M.; Masani, K. Postural sway during quiet standing is related to physiological tremor and muscle volume in young and elderly adults. Gait Posture 2012, 35, 11–17. [Google Scholar] [CrossRef]
- Matsuda, K.; Suzuki, Y.; Yoshikawa, N.; Yamamoto, T.; Kiyono, K.; Tanahashi, T.; Endo, T.; Fukada, K.; Nomura, K.; Sakoda, S.; et al. Postural flexibility during quiet standing in healthy elderly and patients with Parkinson’s disease. In Proceedings of the 2016 38th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Orlando, FL, USA, 16–20 August 2016; Volume 2016, pp. 29–32. [Google Scholar] [CrossRef]
- Diener, H.C.; Dichgans, J.; Bacher, M.; Hülser, J.; Liebich, H. Mechanisms of postural ataxia after intake of alcohol. Int. J. Leg. Med. 1983, 90, 159–165. [Google Scholar] [CrossRef]
- Vassar, R.L.; Rose, J. Motor systems and postural instability. Handb. Clin. Neurol. 2014, 125, 237–251. [Google Scholar] [CrossRef]
- Krafczyk, S.; Schlamp, V.; Dieterich, M.; Haberhauer, P.; Brandt, T. Increased body sway at 3.5–8 Hz in patients with phobic postural vertigo. Neurosci. Lett. 1999, 259, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Pauelsen, M.; Jafari, H.; Strandkvist, V.; Nyberg, L.; Gustafsson, T.; Vikman, I.; Röijezon, U. Frequency domain shows: Fall-related concerns and sensorimotor decline explain inability to adjust postural control strategy in older adults. PLoS ONE 2020, 15, e0242608. [Google Scholar] [CrossRef] [PubMed]
- Nisticò, R.; Quattrone, A.; Crasà, M.; De Maria, M.; Vescio, B.; Quattrone, A. Evaluation of rest tremor in different positions in Parkinson’s disease and essential tremor plus. Neurol. Sci. 2022, 43, 3621–3627. [Google Scholar] [CrossRef]
- Kang, S.Y.; Song, S.-K.; Kim, J.-S.; Sohn, Y.H. Unilateral Standing Leg Tremor as the Initial Manifestation of Parkinson Disease. J. Mov. Disord. 2009, 2, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Yungher, D.A.; Morris, T.R.; Dilda, V.; Shine, J.M.; Naismith, S.L.; Lewis, S.J.G.; Moore, S.T. Temporal Characteristics of High-Frequency Lower-Limb Oscillation during Freezing of Gait in Parkinson’s Disease. Park. Dis. 2014, 2014, 606427. [Google Scholar] [CrossRef] [Green Version]
- Giladi, N.; Hausdorff, J.M. The role of mental function in the pathogenesis of freezing of gait in Parkinson’s disease. J. Neurol. Sci. 2006, 248, 173–176. [Google Scholar] [CrossRef]
- Mancini, M.; Weiss, A.; Herman, T.; Hausdorff, J.M. Turn Around Freezing: Community-Living Turning Behavior in People with Parkinson’s Disease. Front. Neurol. 2018, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Grasso, M.; Mazzini, L.; Schieppati, M. Muscle relaxation in Parkinson’s disease: A reaction time study. Mov. Disord. 1996, 11, 411–420. [Google Scholar] [CrossRef]
- Tolleson, C.M.; Dobolyi, D.G.; Roman, O.C.; Kanoff, K.; Barton, S.; Wylie, S.A.; Kubovy, M.; Claassen, D.O. Dysrhythmia of timed movements in Parkinson’s disease and freezing of gait. Brain Res. 2015, 1624, 222–231. [Google Scholar] [CrossRef] [Green Version]
- Maidan, I.; Patashov, D.; Shustak, S.; Fahoum, F.; Gazit, E.; Shapiro, B.; Levy, A.; Sosnik, R.; Giladi, N.; Hausdorff, J.; et al. A new approach to quantifying the EEG during walking: Initial evidence of gait related potentials and their changes with aging and dual tasking. Exp. Gerontol. 2019, 126, 110709. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Neto, A.; Ugrinowitsch, C.; Coelho, D.B.; de Lima-Pardini, A.C.; Barbosa, E.R.; Teixeira, L.A.; Amaro, E.; Horak, F.B.; Mancini, M.; Nucci, M.P.; et al. Freezing of gait, gait initiation, and gait automaticity share a similar neural substrate in Parkinson’s disease. Hum. Mov. Sci. 2022, 86, 103018. [Google Scholar] [CrossRef]
- Grafton, S.T.; Ralston, A.B.; Ralston, J.D. Monitoring of postural sway with a head-mounted wearable device: Effects of gender, participant state, and concussion. Med. Devices Évid. Res. 2019, 12, 151–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodworth, A.D.; Mellodge, P.; Peterka, R.J. Stance width changes how sensory feedback is used for multisegmental balance control. J. Neurophysiol. 2014, 112, 525–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Newell, K.M. Inter-foot coordination dynamics of quiet standing postures. Neurosci. Biobehav. Rev. 2014, 47, 194–202. [Google Scholar] [CrossRef]
- Promsri, A.; Haid, T.; Federolf, P. Complexity, Composition, and Control of Bipedal Balancing Movements as the Postural Control System Adapts to Unstable Support Surfaces or Altered Feet Positions. Neuroscience 2020, 430, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Peterka, R.J.; Black, F.O. Age-related changes in human posture control: Sensory organization tests. J. Vestib. Res. 1990, 1, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Abrahamová, D.; Hlavačka, F. Age-related changes of human balance during quiet stance. Physiol. Res. 2008, 57, 957–964. [Google Scholar] [CrossRef]
- Poulain, I.; Giraudet, G. Age-related changes of visual contribution in posture control. Gait Posture 2008, 27, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.B.; Taylor, W.R.; Madigan, M.L.; Nussbaum, M.A. The spectral content of postural sway during quiet stance: Influences of age, vision and somatosensory inputs. J. Electromyogr. Kinesiol. 2012, 22, 131–136. [Google Scholar] [CrossRef]
- Sarabon, N.; Panjan, A.; Latash, M. The effects of aging on the rambling and trembling components of postural sway: Effects of motor and sensory challenges. Gait Posture 2013, 38, 637–642. [Google Scholar] [CrossRef]
- Tomomitsu, M.S.; Alonso, A.C.; Morimoto, E.; Bobbio, T.G.; Greve, J.M. Static and dynamic postural control in low-vision and normal-vision adults. Clinics 2013, 68, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, C.; Egami, N.; Demura, S.; Yamasoba, T.; Iwasaki, S. The effect of aging on the center-of-pressure power spectrum in foam posturography. Neurosci. Lett. 2015, 585, 92–97. [Google Scholar] [CrossRef]
- Ozdemir, R.A.; Contreras-Vidal, J.L.; Paloski, W.H. Cortical control of upright stance in elderly. Mech. Ageing Dev. 2018, 169, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, E.V.; Rose, J.; Pfefferbaum, A. Effect of Vision, Touch and Stance on Cerebellar Vermian-related Sway and Tremor: A Quantitative Physiological and MRI Study. Cereb. Cortex 2006, 16, 1077–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colnaghi, S.; Honeine, J.-L.; Sozzi, S.; Schieppati, M. Body Sway Increases After Functional Inactivation of the Cerebellar Vermis by cTBS. Cerebellum 2017, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Barmack, N.H.; Pettorossi, V.E. Adaptive Balance in Posterior Cerebellum. Front. Neurol. 2021, 12, 635259. [Google Scholar] [CrossRef]
- Minassian, K.; Hofstoetter, U.S.; Dzeladini, F.; Guertin, P.A.; Ijspeert, A. The Human Central Pattern Generator for Locomotion: Does It Exist and Contribute to Walking? Neurosci. 2017, 23, 649–663. [Google Scholar] [CrossRef]
- Honeine, J.-L.; Schieppati, M.; Crisafulli, O.; Do, M.-C. The Neuro-Mechanical Processes That Underlie Goal-Directed Medio-Lateral APA during Gait Initiation. Front. Hum. Neurosci. 2016, 10, 445. [Google Scholar] [CrossRef] [Green Version]
- Welter, M.-L.; Do, M.C.; Chastan, N.; Torny, F.; Bloch, F.; du Montcel, S.T.; Agid, Y. Control of vertical components of gait during initiation of walking in normal adults and patients with progressive supranuclear palsy. Gait Posture 2007, 26, 393–399. [Google Scholar] [CrossRef]
- Bishop, M.D.; Brunt, D.; Kukulka, C.; Tillman, M.D.; Pathare, N. Braking impulse and muscle activation during unplanned gait termination in human subjects with parkinsonism. Neurosci. Lett. 2003, 348, 89–92. [Google Scholar] [CrossRef]
- Nieuwboer, A.; Dom, R.; De Weerdt, W.; Desloovere, K.; Janssens, L.; Stijn, V. Electromyographic profiles of gait prior to onset of freezing episodes in patients with Parkinson’s disease. Brain 2004, 127, 1650–1660. [Google Scholar] [CrossRef]
- Warabi, T.; Furuyama, H.; Kato, M. Gait bradykinesia: Difficulty in switching posture/gait measured by the anatomical y-axis vector of the sole in Parkinson’s disease. Exp. Brain Res. 2020, 238, 139–151. [Google Scholar] [CrossRef]
- Shine, J.; Handojoseno, A.; Nguyen, T.; Tran, Y.; Naismith, S.; Nguyen, H.; Lewis, S. Abnormal patterns of theta frequency oscillations during the temporal evolution of freezing of gait in Parkinson’s disease. Clin. Neurophysiol. 2014, 125, 569–576. [Google Scholar] [CrossRef]
- Raymakers, J.; Samson, M.; Verhaar, H. The assessment of body sway and the choice of the stability parameter(s). Gait Posture 2005, 21, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Stelmach, G.E.; Teasdale, N.; Di Fabio, R.P.; Phillips, J. Age Related Decline in Postural Control Mechanisms. Int. J. Aging Hum. Dev. 1989, 29, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Maetzler, W.; Nieuwhof, F.; Hasmann, S.E.; Bloem, B.R. Emerging therapies for gait disability and balance impairment: Promises and pitfalls. Mov. Disord. 2013, 28, 1576–1586. [Google Scholar] [CrossRef]
- Hupfeld, K.E.; McGregor, H.R.; Hass, C.J.; Pasternak, O.; Seidler, R.D. Sensory system-specific associations between brain structure and balance. Neurobiol. Aging 2022, 119, 102–116. [Google Scholar] [CrossRef]
- Viseux, F.J.F.; Delval, A.; Defebvre, L.; Simoneau, M. Postural instability in Parkinson’s disease: Review and bottom-up rehabilitative approaches. Neurophysiol. Clin. 2020, 50, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Patikas, D.A.; Papavasileiou, A.; Ekizos, A.; Hatzitaki, V.; Arampatzis, A. Swaying slower reduces the destabilizing effects of a compliant surface on voluntary sway dynamics. PLoS ONE 2019, 14, e0226263. [Google Scholar] [CrossRef] [Green Version]
- Donath, L.; Kurz, E.; Roth, R.; Zahner, L.; Faude, O. Leg and trunk muscle coordination and postural sway during increasingly difficult stand-ing balance tasks in young and older adults. Maturitas 2016, 91, 60–68. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | Equation | R2 | Slope Different from Zero | Difference between Slope of Non-Adapted and Adapted Trials |
|---|---|---|---|---|
| Dominant frequency | ||||
| Foam Non-adapted | y = 0.81x + 0.48 | 0.63 | p < 0.001 | p < 0.01 |
| Foam Adapted | y = 0.32x + 2.37 | 0.42 | p < 0.001 | |
| Solid Non-adapted | y = 0.64x + 1.65 | 0.38 | p < 0.001 | p = 0.83 |
| Solid Adapted | y = 0.71x + 1.36 | 0.25 | p = 0.012 | |
| Peak amplitude | ||||
| Foam Non-adapted | y = 2.01x + 0.04 | 0.4 | p < 0.001 | p = 0.47 |
| Foam Adapted | y = 1.55x + 0.02 | 0.49 | p < 0.001 | |
| Solid Non-adapted | y = 0.75x + 0.00005 | 0.37 | p < 0.01 | p = 0.21 |
| Solid Adapted | y = 0.36x + 0.001 | 0.1 | p = 0.13 | |
| SD of Gaussian fit | ||||
| Foam Non-adapted | y = 0.59x + 0.31 | 0.24 | p = 0.015 | p = 0.053 |
| Foam Adapted | y = 0.07x + 0.96 | 0.018 | p = 0.53 | |
| Solid Non-adapted | y = 0.03x + 2.6 | 0.0009 | p = 0.89 | p = 0.83 |
| Solid Adapted | y = 0.1x + 2.18 | 0.01 | p = 0.59 | |
| Conditions | Equation | R2 | Slope Different from Zero | Difference between Slope of Non-Adapted and Adapted Trials | Difference in Slope between Conditions |
|---|---|---|---|---|---|
| ECF Non-adapted | y = −0.018x + 4.88 | 0.39 | p < 0.01 | p = 0.53 | vs. ECS: p = 0.63 vs. EOF: p = 0.77 vs. EOS: p = 0.79 |
| ECF Adapted | y = −0.01x + 4.56 | 0.38 | p < 0.01 | vs. ECS: p = 0.83 vs. EOF: p = 0.86 vs. EOS: p = 0.25 | |
| ECS Non-adapted | y = −0.009x + 5.32 | 0.02 | p = 0.52 | p = 0.73 | vs. ECF: p = 0.63 vs. EOF: p = 0.54 vs. EOS: p = 0.86 |
| ECS Adapted | y = −0.017x + 5.9 | 0.05 | p = 0.29 | vs. ECF: p = 0.83 vs. EOF: p = 0.97 vs. EOS: p = 0.46 | |
| EOF Non-adapted | y = −0.019x + 5.28 | 0.49 | p < 0.001 | p = 0.86 | vs. ECF: p = 0.77 vs. ECS: p = 0.54 vs. EOS: p = 0.69 |
| EOF Adapted | y = −0.016x + 5.02 | 0.12 | p = 0.09 | vs. ECF: p = 0.86 vs. ECS: p = 0.97 vs. EOS: p = 0.61 | |
| EOS Non-adapted | y = −0.014x + 5.61 | 0.04 | p = 0.35 | p = 0.48 | vs. ECF: p = 0.79 vs. ECS: p = 0.86 vs. EOF: p = 0.69 |
| EOS Adapted | y = −0.026x + 6.6 | 0.24 | p < 0.05 | vs. ECF: p = 0.25 vs. ECS: p = 0.46 vs. EOF: p = 0.61 |
| Conditions | Equation | R2 | Slope Different from Zero | Difference between Slope of Non-Adapted and Adapted Trials | Difference in Slope between Conditions |
|---|---|---|---|---|---|
| ECF Non-adapted | y = 0.0018x − 0.03 | 0.26 | p < 0.05 | p = 0.60 | vs. ECS: p < 0.05 vs. EOF: p = 0.16 vs. EOS: p < 0.05 |
| ECF Adapted | y = 0.001x − 0.048 | 0.41 | p < 0.001 | vs. ECS: p < 0.001 vs. EOF: p = 0.24 vs. EOS: p < 0.001 | |
| ECS Non-adapted | y = 0.0001x − 0.005 | 0.38 | p < 0.01 | p = 0.37 | vs. ECF: p < 0.05 vs. EOF: p < 0.001 vs. EOS: p = 0.12 |
| ECS Adapted | y = 0.00007x − 0.003 | 0.26 | p < 0.05 | vs. ECF: p < 0.001 vs. EOF: p < 0.001 vs. EOS: p = 0.81 | |
| EOF Non-adapted | y = 0.0008x − 0.03 | 0.49 | p < 0.001 | p = 0.66 | vs. ECF: p = 0.16 vs. ECS: p < 0.001 vs. EOS: p < 0.001 |
| EOF Adapted | y = 0.0009x − 0.04 | 0.57 | p < 0.001 | vs. ECF: p = 0.24 vs. ECS: p < 0.001 vs. EOS: p < 0.001 | |
| EOS Non-adapted | y = 0.00004x − 0.0007 | 0.09 | p = 0.13 | p = 0.68 | vs. ECF: p < 0.05 vs. ECS: p = 0.12 vs. EOF: p < 0.001 |
| EOS Adapted | y = 0.00006x − 0.002 | 0.26 | p < 0.01 | vs. ECF: p < 0.001 vs. ECS: p = 0.81 vs. EOF: p < 0.001 |
| Condition | Non-Adapted | Adapted | ||
|---|---|---|---|---|
| ML | AP | ML | AP | |
| ECF | 0.34 | 0.36 | 0.24 | 0.24 |
| ECS | 0.26 | 0.21 | 0.26 | 0.16 |
| EOF | 0.23 | 0.19 | 0.2 | 0.18 |
| EOS | 0.22 | 0.18 | 0.19 | 0.17 |
| Conditions | Non-Adapted | Adapted | ||
|---|---|---|---|---|
| ML | AP | ML | AP | |
| ECF | −0.76 ± 0.08 | −0.74 ± 0.07 | −0.64 ± 0.06 | −0.60 ± 0.09 |
| ECS | −0.55 ± 0.14 | −0.49 ± 0.15 | −0.50 ± 0.14 | −0.47 ± 0.13 |
| EOF | −0.59 ± 0.08 | −0.54 ± 0.08 | −0.52 ± 0.10 | −0.51 ± 0.10 |
| EOS | −0.51 ± 0.10 | −0.43 ± 0.11 | −0.45 ± 0.16 | −0.43 ± 0.12 |
| Condition | Equation | R2 | Slope Different from Zero | Difference Between Slope of Non-Adapted and Adapted Trials | Difference in Slope Between Conditions |
|---|---|---|---|---|---|
| ECF Non-adapted | y = 0.705x + 1.50 | 0.77 | p < 0.001 | p = 0.67 | vs. ECS: p = 0.83 vs. EOF: p = 0.17 vs. EOS: p = 0.59 |
| ECF Adapted | y = 0.65x + 1.19 | 0.76 | p < 0.001 | vs. ECS: p < 0.05 vs. EOF: p = 0.97 vs. EOS: p = 0.26 | |
| ECS Non-adapted | y = 0.77x + 0.214 | 0.58 | p < 0.001 | p = 0.06 | vs. ECF: p = 0.83 vs. EOF: p = 0.21 vs. EOS: p = 0.68 |
| ECS Adapted | y = 1.19x − 0.207 | 0.71 | p < 0.001 | vs. ECF: p < 0.05 vs. EOF: p < 0.05 vs. EOS: p < 0.01 | |
| EOF Non-adapted | y = 1.06x + 0.03 | 0.64 | p < 0.001 | p = 0.09 | vs. ECF: p = 0.17 vs. ECS: p = 0.21 vs. EOS: p = 0.51 |
| EOF Adapted | y = 0.66x + 0.55 | 0.45 | p < 0.001 | vs. ECF: p = 0.97 vs. ECS: p < 0.05 vs. EOS: p = 0.35 | |
| EOS Non-adapted | y = 0.88x + 0.036 | 0.41 | p < 0.001 | p = 0.12 | vs. ECF: p = 0.59 vs. ECS: p = 0.68 vs. EOF: p = 0.51 |
| EOS Adapted | y = 0.44x + 0.5 | 0.24 | p < 0.05 | vs. ECF: p = 0.26 vs. ECS: p < 0.01 vs. EOF: p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sozzi, S.; Ghai, S.; Schieppati, M. The ‘Postural Rhythm’ of the Ground Reaction Force during Upright Stance and Its Conversion to Body Sway—The Effect of Vision, Support Surface and Adaptation to Repeated Trials. Brain Sci. 2023, 13, 978. https://doi.org/10.3390/brainsci13070978
Sozzi S, Ghai S, Schieppati M. The ‘Postural Rhythm’ of the Ground Reaction Force during Upright Stance and Its Conversion to Body Sway—The Effect of Vision, Support Surface and Adaptation to Repeated Trials. Brain Sciences. 2023; 13(7):978. https://doi.org/10.3390/brainsci13070978
Chicago/Turabian StyleSozzi, Stefania, Shashank Ghai, and Marco Schieppati. 2023. "The ‘Postural Rhythm’ of the Ground Reaction Force during Upright Stance and Its Conversion to Body Sway—The Effect of Vision, Support Surface and Adaptation to Repeated Trials" Brain Sciences 13, no. 7: 978. https://doi.org/10.3390/brainsci13070978