

Mu Rhythm Desynchronization while Observing Rubber Hand Movement in the Mirror: The Interaction of Body Representation with Visuo-Tactile Stimulation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
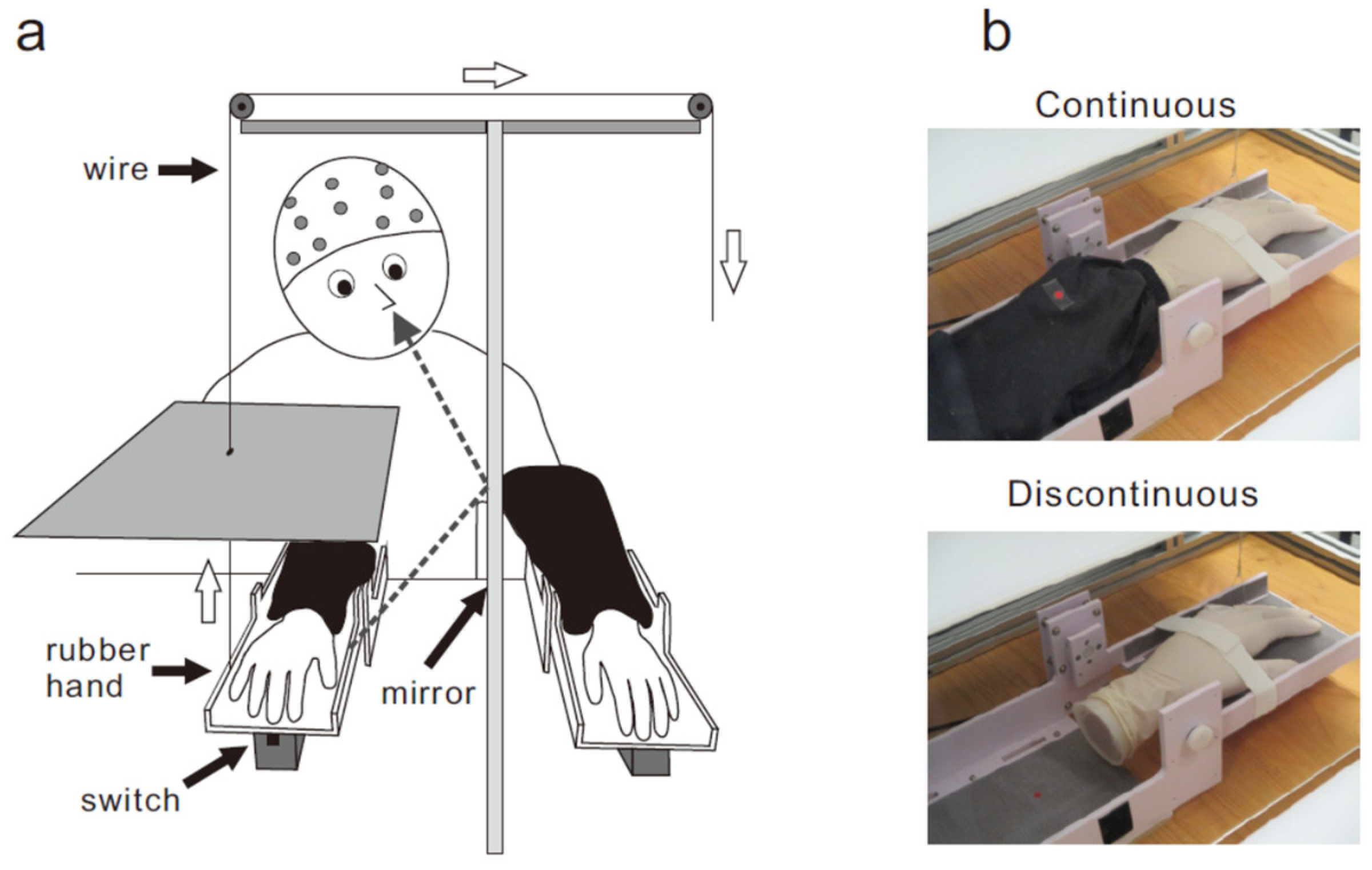
2.2. Apparatus
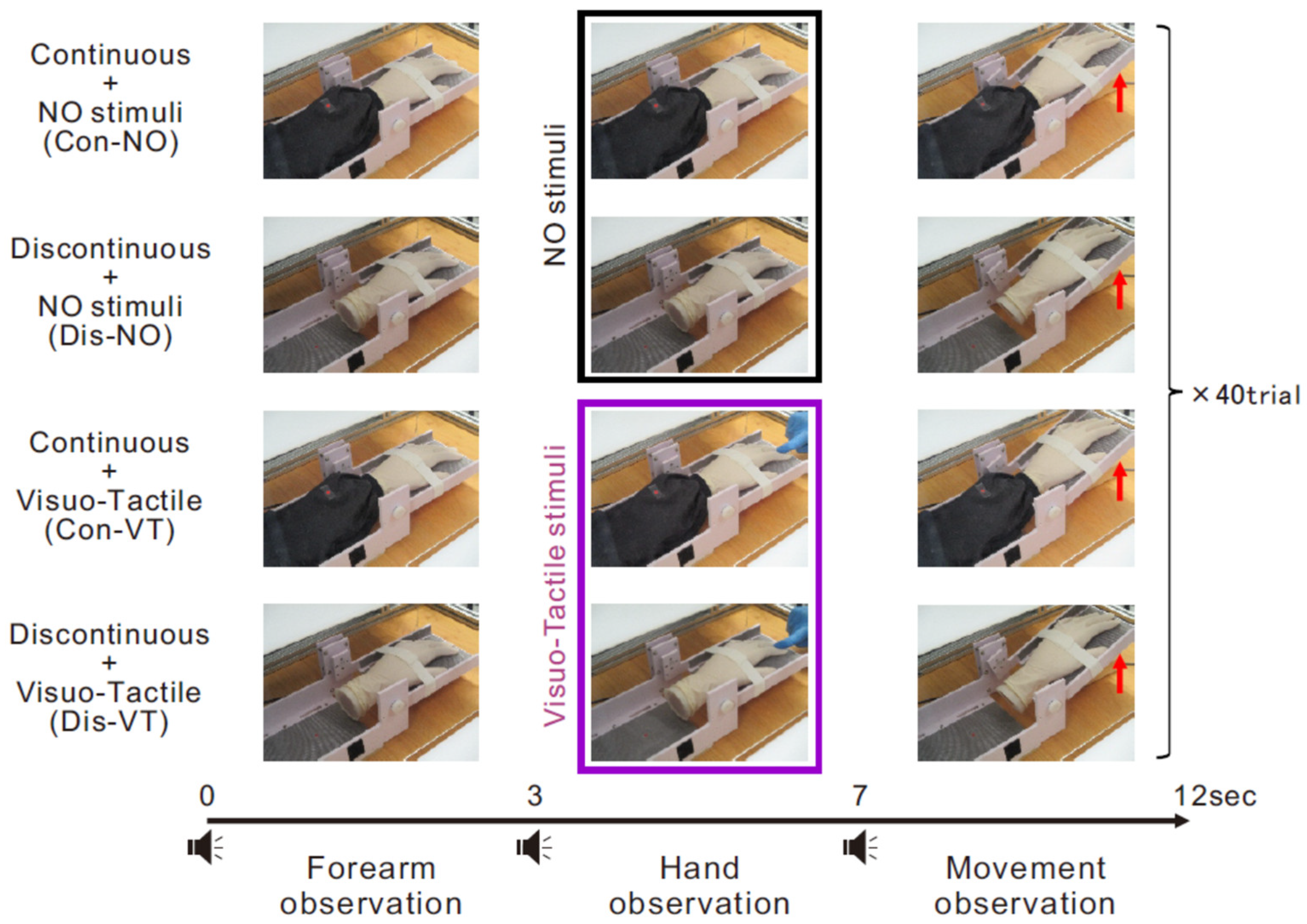
2.3. Procedure
2.4. Acquisition and Analysis of EEG Data
3. Results
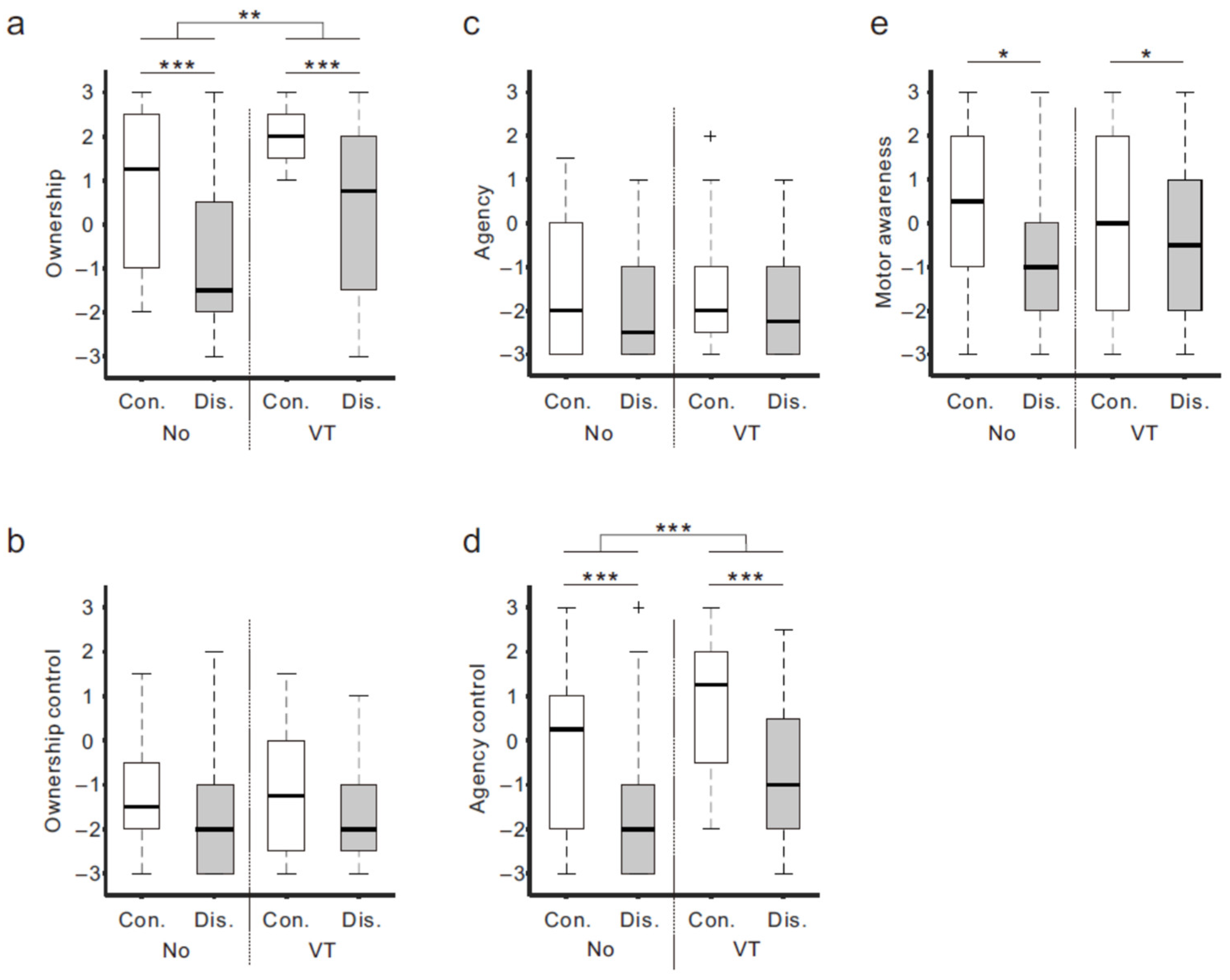
3.1. Questionnaire Rating
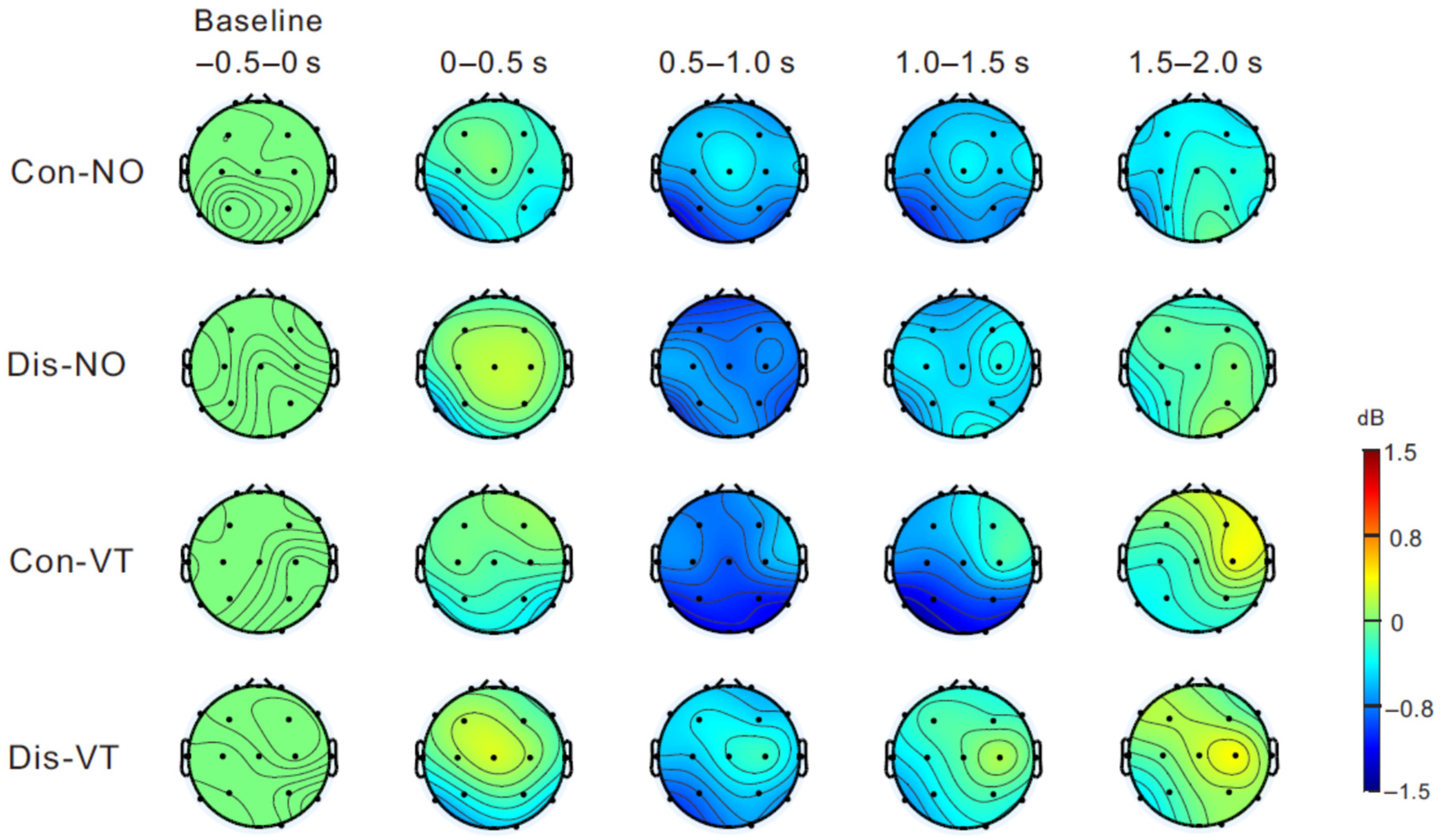
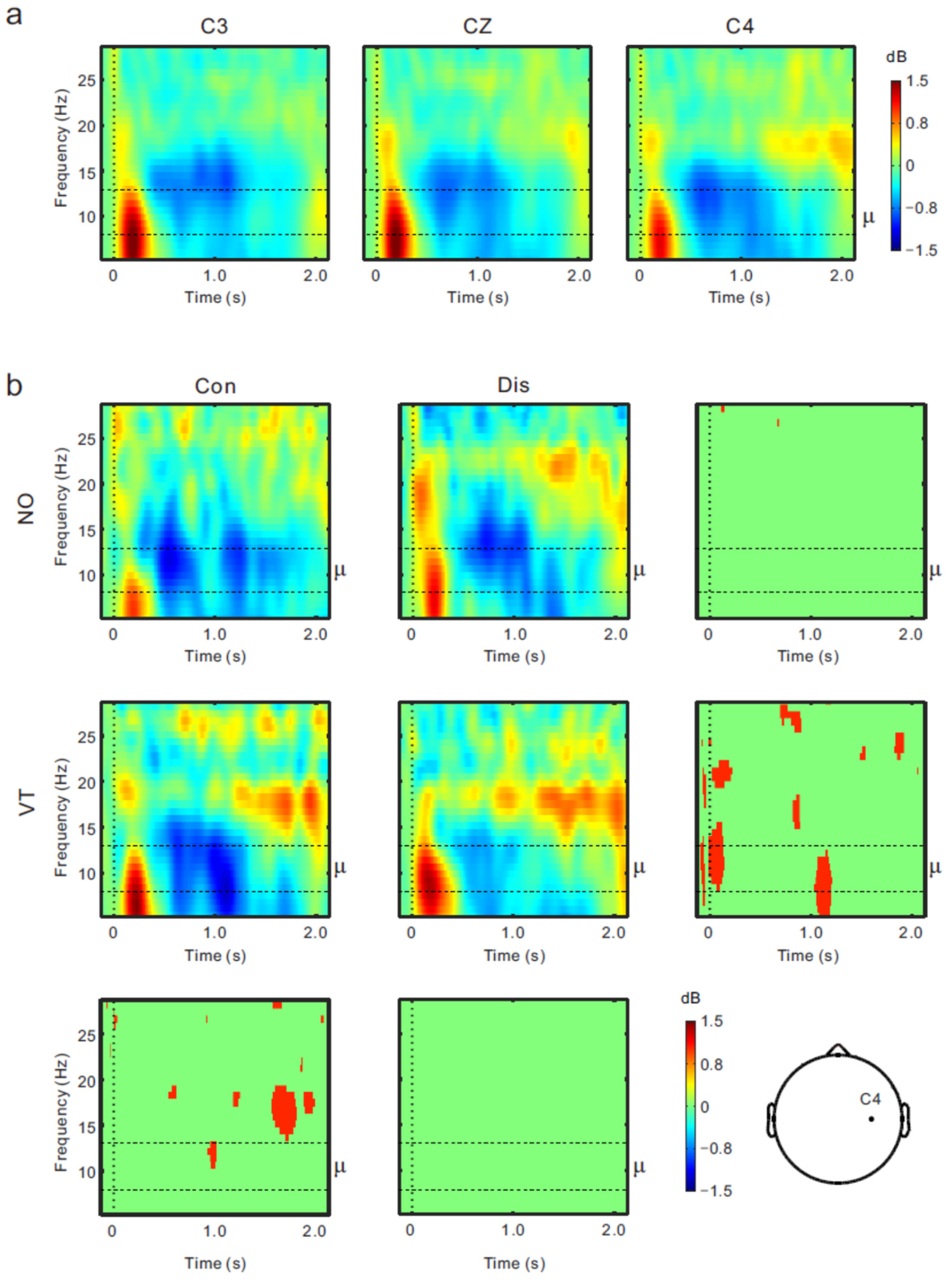
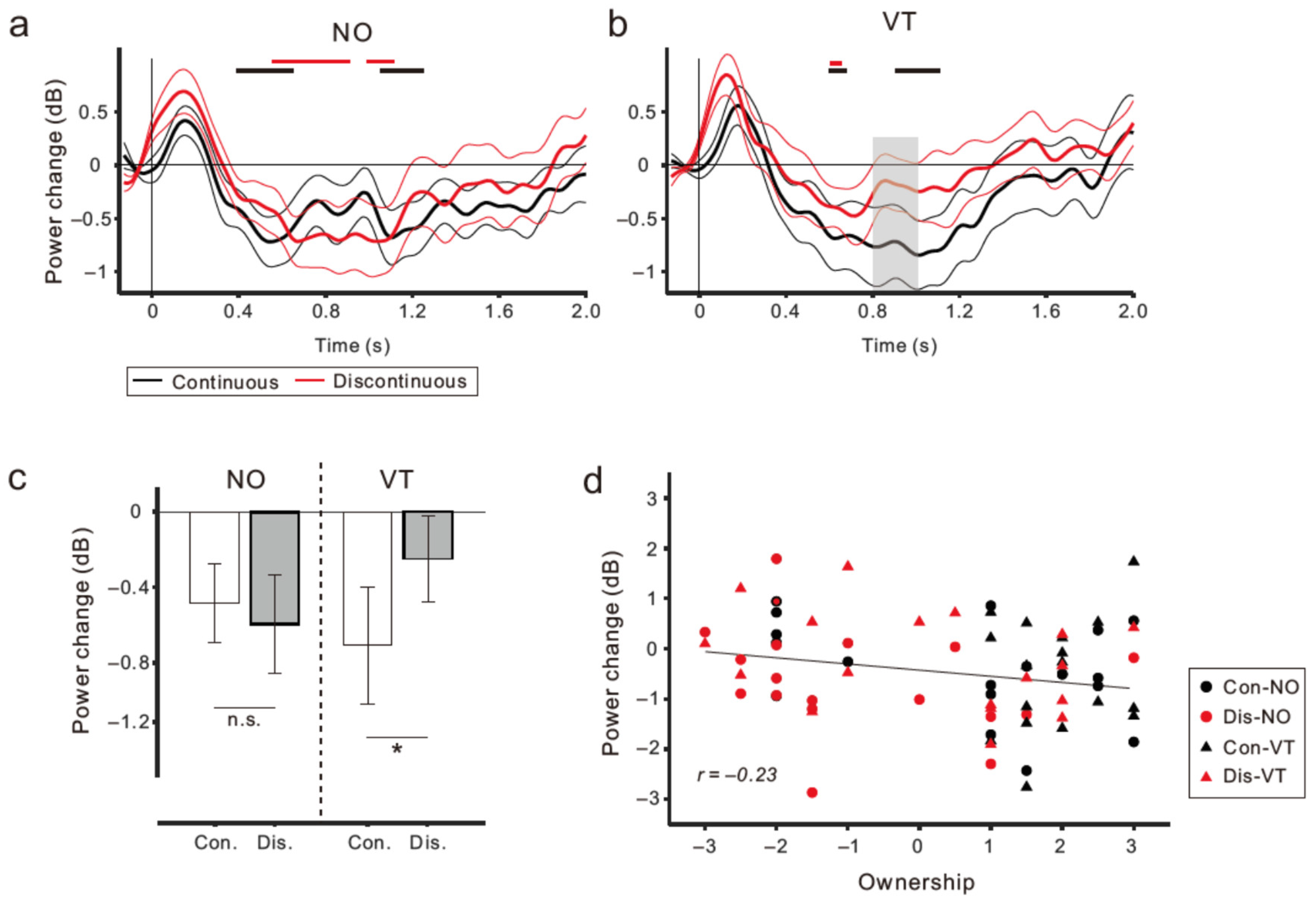
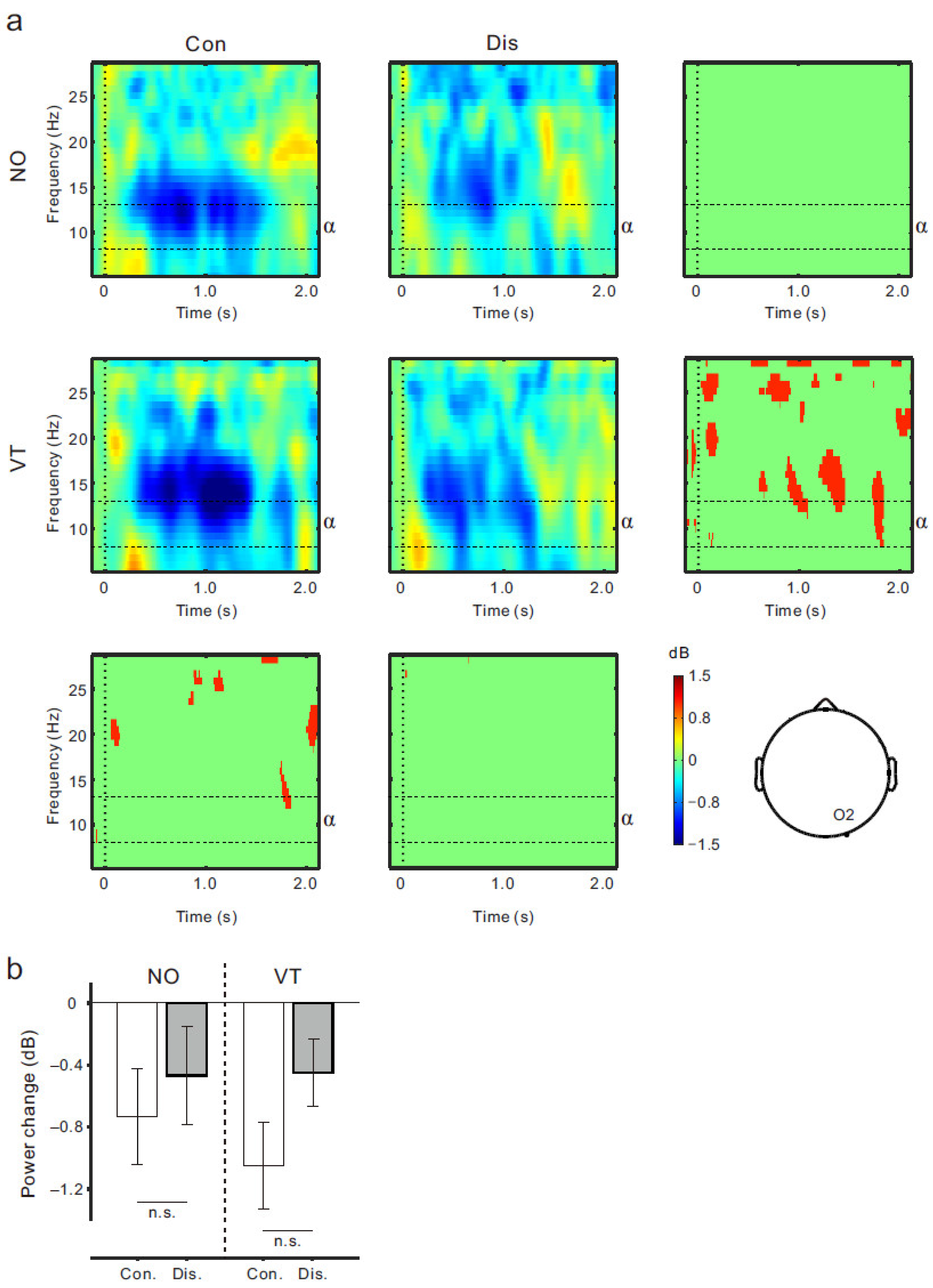
3.2. EEG Results
4. Discussion
4.1. Μu Rhythm ERD and Sense of Body Ownership
4.2. Sense of Agency and Motor Awareness during Movement Observation
4.3. Limitation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gallagher, S. Philosophical conceptions of the self: Implications for cognitive science. Trends Cogn. Sci. 2000, 4, 14–21. [Google Scholar] [CrossRef]
- Botvinick, M.; Cohen, J. Rubber hands ‘feel’ touch that eyes see. Nature 1998, 391, 756. [Google Scholar] [CrossRef]
- Zeller, D.; Litvak, V.; Friston, K.J.; Classen, J. Sensory processing and the rubber hand illusion-an evoked potentials study. J. Cogn. Neurosci. 2015, 27, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Tsakiris, M.; Haggard, P. The rubber hand illusion revisited: Visuotactile integration and self-attribution. J. Exp. Psychol. Hum. Percept. Perform. 2005, 31, 80–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsakiris, M. My body in the brain: A neurocognitive model of body-ownership. Neuropsychologia 2010, 48, 703–712. [Google Scholar] [CrossRef] [PubMed]
- de Vignemont, F. Embodiment, ownership and disownership. Conscious. Cogn. 2011, 20, 82–93. [Google Scholar] [CrossRef] [Green Version]
- Longo, M.R.; Schuur, F.; Kammers, M.P.; Tsakiris, M.; Haggard, P. What is embodiment? A psychometric approach. Cognition 2008, 107, 978–998. [Google Scholar] [CrossRef] [Green Version]
- Kilteni, K.; Normand, J.M.; Sanchez-Vives, M.V.; Slater, M. Extending body space in immersive virtual reality: A very long arm illusion. PLoS ONE 2012, 7, e40867. [Google Scholar] [CrossRef] [Green Version]
- Slater, M.; Perez-Marcos, D.; Ehrsson, H.H.; Sanchez-Vives, M.V. Towards a digital body: The virtual arm illusion. Front. Hum. Neurosci. 2008, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Vives, M.V.; Spanlang, B.; Frisoli, A.; Bergamasco, M.; Slater, M. Virtual hand illusion induced by visuomotor correlations. PLoS ONE 2010, 5, e10381. [Google Scholar] [CrossRef] [Green Version]
- Pyasik, M.; Tieri, G.; Pia, L. Visual appearance of the virtual hand affects embodiment in the virtual hand illusion. Sci. Rep. 2020, 10, 5412. [Google Scholar] [CrossRef] [Green Version]
- Tieri, G.; Tidoni, E.; Pavone, E.F.; Aglioti, S.M. Mere observation of body discontinuity affects perceived ownership and vicarious agency over a virtual hand. Exp. Brain Res. 2015, 233, 1247–1259. [Google Scholar] [CrossRef]
- Tieri, G.; Tidoni, E.; Pavone, E.F.; Aglioti, S.M. Body visual discontinuity affects feeling of ownership and skin conductance responses. Sci. Rep. 2015, 5, 17139. [Google Scholar] [CrossRef] [Green Version]
- Casula, E.P.; Tieri, G.; Rocchi, L.; Pezzetta, R.; Maiella, M.; Pavone, E.F.; Aglioti, S.M.; Koch, G. Feeling of Ownership over an Embodied Avatar’s Hand Brings about Fast Changes of Fronto-Parietal Cortical Dynamics. J. Neurosci. 2022, 42, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Metral, M.; Guerraz, M. Fake hand in movement: Visual motion cues from the rubber hand are processed for kinesthesia. Conscious. Cogn. 2019, 73, 102761. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, S.; Unenaka, S.; Shimada, S.; Ohki, Y. Distinct modulation of mu and beta rhythm desynchronization during observation of embodied fake hand rotation. Neuropsychologia 2021, 159, 107952. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, S.; Unenaka, S.; Zama, T.; Shimada, S.; Ohki, Y. Sensorimotor and Posterior Brain Activations During the Observation of Illusory Embodied Fake Hand Movement. Front. Hum. Neurosci. 2019, 13, 367. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, S.; Unenaka, S.; Zama, T.; Shimada, S.; Ohki, Y. Spontaneous imitative movements induced by an illusory embodied fake hand. Neuropsychologia 2018, 111, 77–84. [Google Scholar] [CrossRef]
- Raz, G.; Gurevitch, G.; Vaknin, T.; Aazamy, A.; Gefen, I.; Grunstein, S.; Azouri, G.; Goldway, N. Electroencephalographic evidence for the involvement of mirror-neuron and error-monitoring related processes in virtual body ownership. Neuroimage 2020, 207, 116351. [Google Scholar] [CrossRef]
- Fox, N.A.; Bakermans-Kranenburg, M.J.; Yoo, K.H.; Bowman, L.C.; Cannon, E.N.; Vanderwert, R.E.; Ferrari, P.F.; van Ijzendoorn, M.H. Assessing human mirror activity with EEG mu rhythm: A meta-analysis. Psychol. Bull. 2016, 142, 291–313. [Google Scholar] [CrossRef]
- Pineda, J.A. The functional significance of mu rhythms: Translating “seeing” and “hearing” into “doing”. Brain Res. Rev. 2005, 50, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, F.; Yasojima, T.; Kizuka, T. Kinesthetic illusory feeling induced by a finger movement movie effects on corticomotor excitability. Neuroscience 2007, 149, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, F.; Blanchard, C.; Lebar, N.; Nazarian, B.; Kavounoudias, A.; Romaiguere, P. Brain Regions Associated to a Kinesthetic Illusion Evoked by Watching a Video of One’s Own Moving Hand. PLoS ONE 2015, 10, e0131970. [Google Scholar] [CrossRef]
- Shibata, E.; Kaneko, F. Event-related desynchronization possibly discriminates the kinesthetic illusion induced by visual stimulation from movement observation. Exp. Brain Res. 2019, 237, 3233–3240. [Google Scholar] [CrossRef] [PubMed]
- Honma, M.; Yoshiike, T.; Ikeda, H.; Kim, Y.; Kuriyama, K. Sleep dissolves illusion: Sleep withstands learning of visuo-tactile-proprioceptive integration induced by repeated days of rubber hand illusion training. PLoS ONE 2014, 9, e85734. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Kalckert, A.; Ehrsson, H.H. Moving a Rubber Hand that Feels Like Your Own: A Dissociation of Ownership and Agency. Front. Hum. Neurosci. 2012, 6, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.J.; Sejnowski, T.J. An information-maximization approach to blind separation and blind deconvolution. Neural Comput. 1995, 7, 1129–1159. [Google Scholar] [CrossRef]
- Winkler, I.; Haufe, S.; Tangermann, M. Automatic Classification of Artifactual ICA-Components for Artifact Removal in EEG Signals. Behav. Brain Funct. 2011, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Makeig, S. Auditory event-related dynamics of the EEG spectrum and effects of exposure to tones. Electroencephalogr. Clin. Neurophysiol. 1993, 86, 283–293. [Google Scholar] [CrossRef]
- Asai, T. Illusory body-ownership entails automatic compensative movement: For the unified representation between body and action. Exp. Brain Res. 2015, 233, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Lanillos, P.; Franklin, S.; Maselli, A.; Franklin, D.W. Active strategies for multisensory conflict suppression in the virtual hand illusion. Sci. Rep. 2021, 11, 22844. [Google Scholar] [CrossRef] [PubMed]
- Blakemore, S.J.; Wolpert, D.M.; Frith, C.D. Abnormalities in the awareness of action. Trends Cogn. Sci. 2002, 6, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Frith, C.D.; Blakemore, S.J.; Wolpert, D.M. Abnormalities in the awareness and control of action. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1771–1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, A.; Bentin, S. Does focusing on hand-grasping intentions modulate electroencephalogram mu and alpha suppressions? Neuroreport 2010, 21, 1050–1054. [Google Scholar] [CrossRef]
- Hobson, H.M.; Bishop, D.V. Mu suppression—A good measure of the human mirror neuron system? Cortex 2016, 82, 290–310. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Question |
|---|---|
| Ownership | 1. I felt as if I were looking at my own hand. |
| 2. I felt as if the rubber hand was my hand. | |
| Ownership control | 3. It seemed as if I might have more than one left hand or arm. |
| 4. It felt as if I no longer had a left hand, as if my left hand had disappeared. | |
| Agency | 5. I felt as if I was controlling the movements of the rubber hand. |
| 6. I felt as if I was causing the movements of the rubber hand. | |
| Agency control | 7. I felt as if the rubber hand was controlling my hand movements. |
| 8. I felt as if the rubber hand was controlling my will. | |
| Motor awareness | 9. I felt as if my hand was moving against my will when observing the rubber hand movement. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shibuya, S.; Ohki, Y. Mu Rhythm Desynchronization while Observing Rubber Hand Movement in the Mirror: The Interaction of Body Representation with Visuo-Tactile Stimulation. Brain Sci. 2023, 13, 969. https://doi.org/10.3390/brainsci13060969
Shibuya S, Ohki Y. Mu Rhythm Desynchronization while Observing Rubber Hand Movement in the Mirror: The Interaction of Body Representation with Visuo-Tactile Stimulation. Brain Sciences. 2023; 13(6):969. https://doi.org/10.3390/brainsci13060969
Chicago/Turabian StyleShibuya, Satoshi, and Yukari Ohki. 2023. "Mu Rhythm Desynchronization while Observing Rubber Hand Movement in the Mirror: The Interaction of Body Representation with Visuo-Tactile Stimulation" Brain Sciences 13, no. 6: 969. https://doi.org/10.3390/brainsci13060969