
Ischemic Preconditioning Provides Neuroprotection by Inhibiting NLRP3 Inflammasome Activation and Cell Pyroptosis
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Middle Cerebral Artery Occulusion Model (MCAO) Establishment
2.3. Neurobehavioral Evaluation
2.4. Infarct Volume Assessment
2.5. Cell Cultures
2.6. Oxygen-Glucose Deprivation (OGD) and IPC Induction
2.7. Cell Viability Analysis
2.8. Western Blotting (WB)
2.9. Enzyme-Linked Immunosorbent Assay (ELISA)
2.10. Immunofluorescence Staining
2.11. Statistical Analysis
3. Results
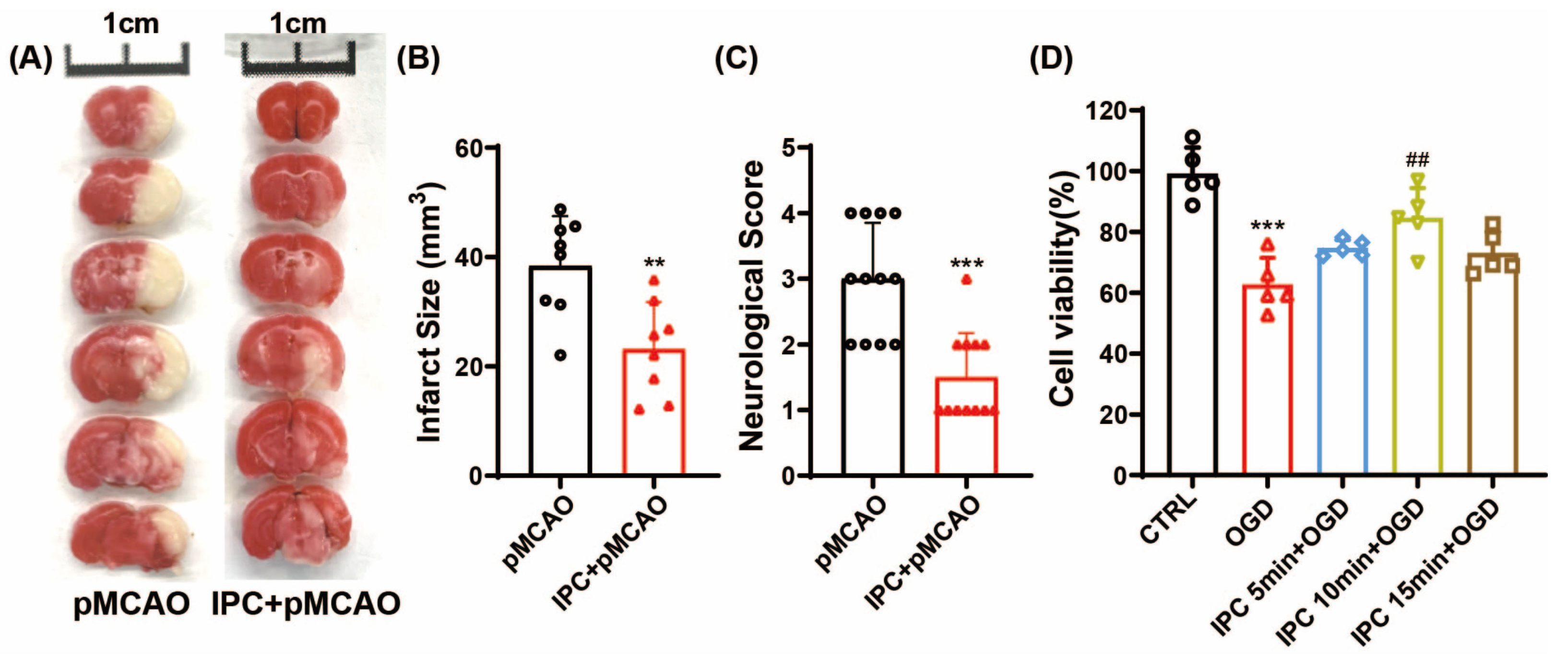
3.1. IPC Could Attenuate Acute Ischemic Injury In Vivo and In Vitro
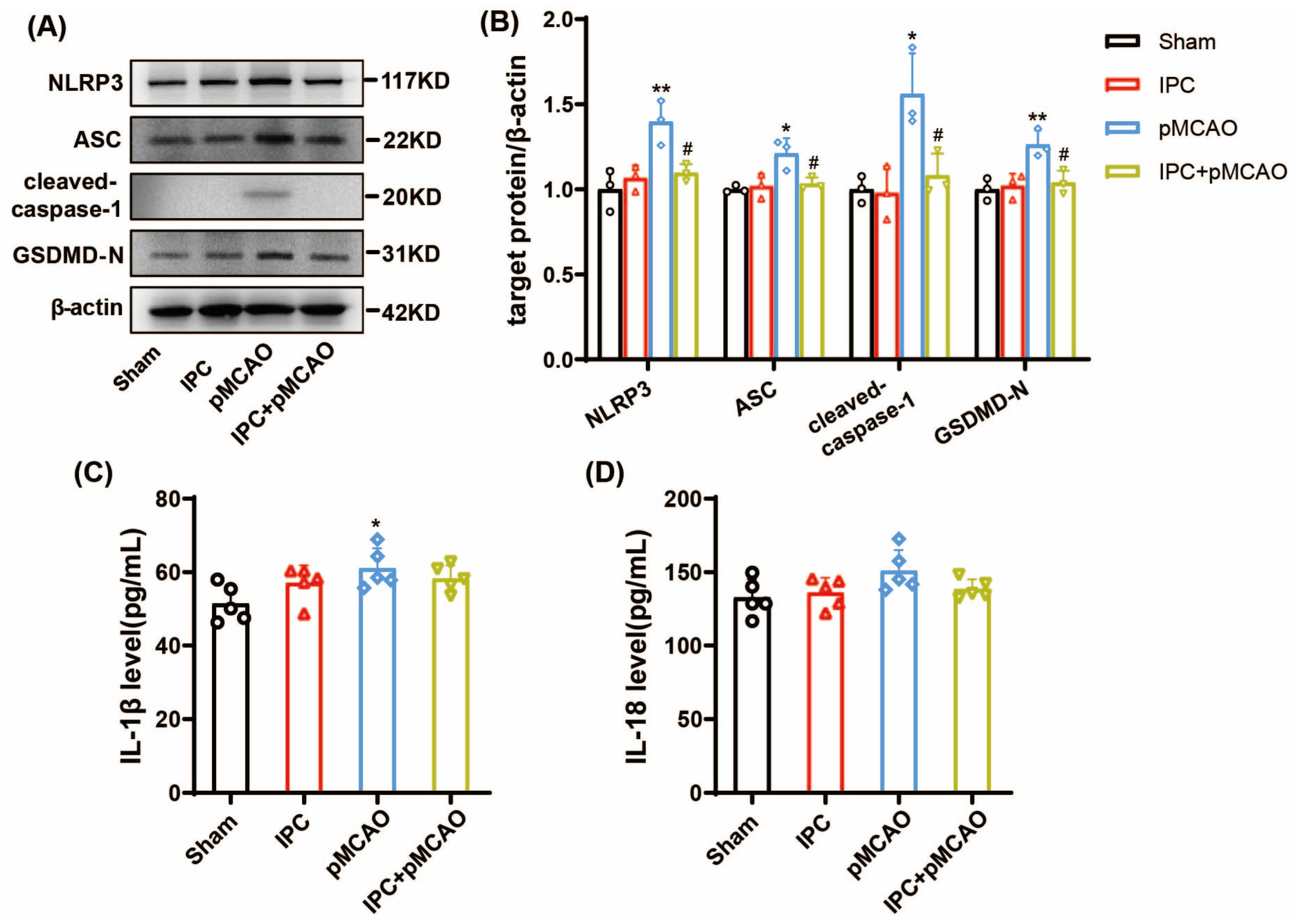
3.2. IPC Inhibited the Activated NLRP3 Inflammasome and Attenuated Inflammation in MCAO Mice
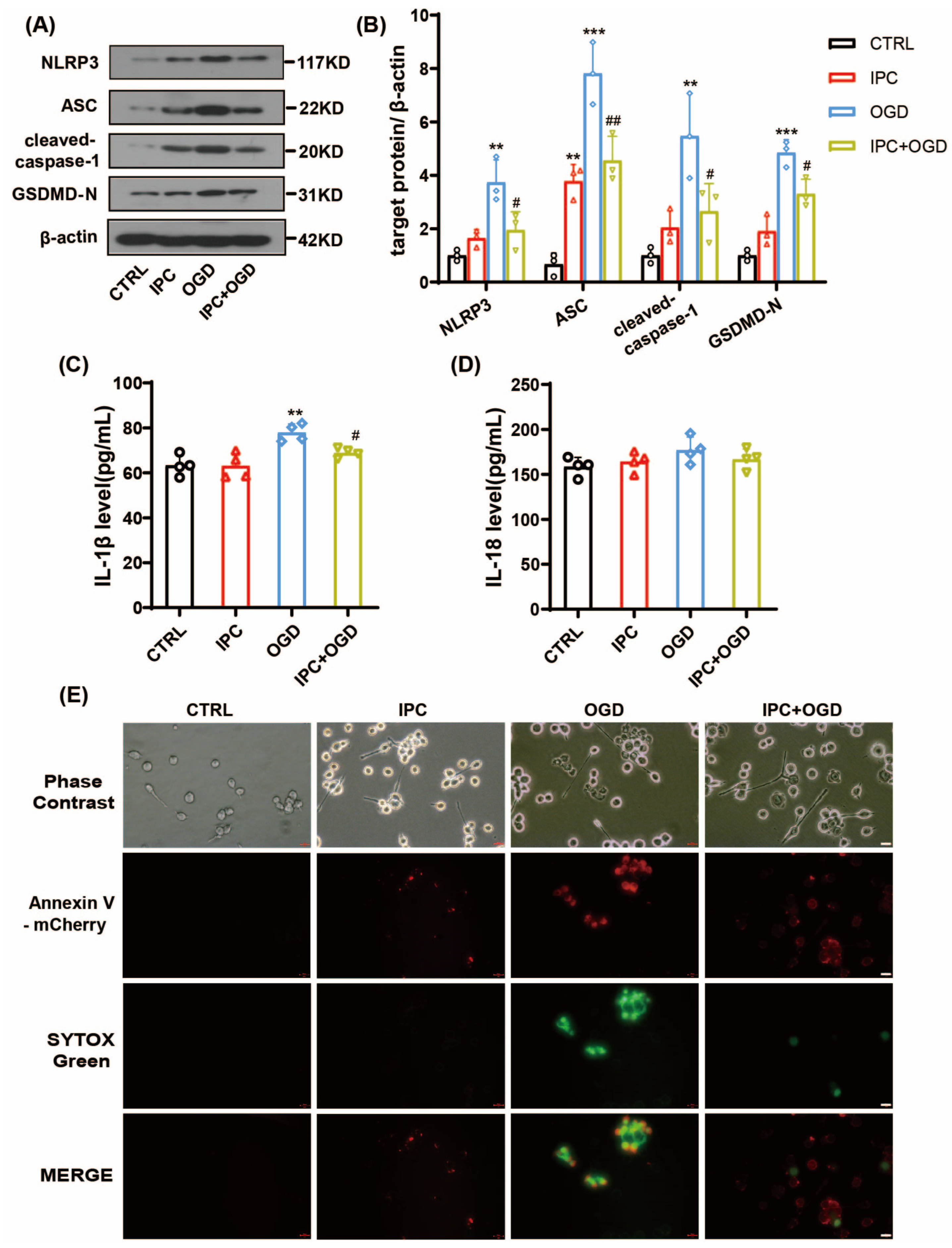
3.3. IPC Could Effectively Reduce NLRP3 Inflammasome Activation after OGD
3.4. IPC Attenuated Cell Pyroptosis after Ischemic Stroke In Vivo and Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gidday, J.M. Cerebral preconditioning and ischaemic tolerance. Nat. Rev. Neurosci. 2006, 7, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Sun, Y.; Li, Q.; Li, S.; Shi, Y.; Leak, R.K.; Chen, J.; Zhang, F. Ischemic preconditioning provides long-lasting neuroprotection against ischemic stroke: The role of Nrf2. Exp. Neurol. 2020, 325, 113142. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.-N.; Guo, W.-T.; Liu, J.; Chang, J.; Ma, H.; Zhang, P.; Zhang, F.-L.; Han, K.; Hu, H.-H.; Jin, H.; et al. Changes in cerebral autoregulation and blood biomarkers after remote ischemic preconditioning. Neurology 2019, 93, e8–e19. [Google Scholar] [CrossRef] [PubMed]
- Yung, L.M.; Wei, Y.; Qin, T.; Wang, Y.; Smith, C.D.; Waeber, C. Sphingosine kinase 2 mediates cerebral preconditioning and protects the mouse brain against ischemic injury. Stroke 2012, 43, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.G.; Muruve, D.A.; Power, C. Inflammasomes in the CNS. Nat. Rev. Neurosci. 2014, 15, 84–97. [Google Scholar] [CrossRef]
- Shi, J.; Gao, W.; Shao, F. Pyroptosis: Gasdermin-mediated programmed necrotic cell death. Trends. Biochem. Sci. 2017, 42, 245–254. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, W.; Li, W.; Zhao, Y. NLRP3 inflammasome: Checkpoint connecting innate and adaptive immunity in autoimmune diseases. Front. Immunol. 2021, 12, 732933. [Google Scholar] [CrossRef]
- Toldo, S.; Mezzaroma, E.; Buckley, L.F.; Potere, N.; Di Nisio, M.; Biondi-Zoccai, G.; Van Tassell, B.W.; Abbate, A. Targeting the NLRP3 inflammasome in cardiovascular diseases. Pharmacol. Ther. 2021, 236, 108053. [Google Scholar] [CrossRef]
- Wang, S.Y.; Fu, X.X.; Duan, R.; Wei, B.; Cao, H.M.; Yan, E.; Chen, S.Y.; Zhang, Y.D.; Jiang, T. The Alzheimer’s disease-associated gene TREML2 modulates inflammation by regulating microglia polarization and NLRP3 inflammasome activation. Neural. Regen. Res. 2023, 18, 434–438. [Google Scholar]
- Gustin, A.; Kirchmeyer, M.; Koncina, E.; Felten, P.; Losciuto, S.; Heurtaux, T.; Tardivel, A.; Heuschling, P.; Dostert, C. NLRP3 inflammasome is expressed and functional in mouse brainmicroglia but not in astrocytes. PLoS ONE 2015, 10, e0130624. [Google Scholar] [CrossRef]
- Fann, D.Y.; Lee, S.Y.; Manzanero, S.; Tang, S.C.; Gelderblom, M.; Chunduri, P.; Bernreuther, C.; Glatzel, M.; Cheng, Y.L.; Thundyil, J.; et al. Intravenous immunoglobulin suppresses NLRP1 and NLRP3 inflammasome-mediated neuronal death in ischemic stroke. Cell Death Dis. 2013, 4, e790. [Google Scholar] [CrossRef] [PubMed]
- Franke, M.; Bieber, M.; Kraft, P.; Weber, A.N.R.; Stoll, G.; Schuhmann, M.K. The NLRP3 inflammasome drives inflammation in ischemia/reperfusion injury after transient middle cerebral artery occlusion in mice. Brain Behav. Immun. 2021, 92, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Yan, J.; Geng, C.; Wang, D.; Li, C.; Feng, S.; Wang, H. A polymorphism within the 3’UTR of NLRP3 is associated with susceptibility for ischemic stroke in Chinese population. Cell. Mol. Neurobiol. 2016, 36, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Dong, Q.; Song, Z.; Shen, F.; Shi, J.; Li, Y. NLRP3 inflammasome: A promising target in ischemic stroke. Inflamm. Res. 2017, 66, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Jiang, W.; Cui, P.; Zheng, K.; Dang, C.; Wang, J.; Li, H.; Chen, L.; Zhang, R.; Wang, Q.M.; et al. Microglial PGC-1α protects against ischemic brain injury by suppressing neuroinflammation. Genome Med. 2021, 13, 47. [Google Scholar] [CrossRef]
- Jiang, T.; Yu, J.-T.; Zhu, X.-C.; Zhang, Q.-Q.; Tan, M.-S.; Cao, L.; Wang, H.-F.; Shi, J.-Q.; Gao, L.; Qin, H.; et al. Ischemic preconditioning provides neuroprotection by induction of AMP-activated protein kinase-dependent autophagy in a rat model of ischemic stroke. Mol. Neurobiol. 2015, 51, 220–229. [Google Scholar] [CrossRef]
- Zhao, A.-P.; Dong, Y.-F.; Liu, W.; Gu, J.; Sun, X.-L. Nicorandil inhibits inflammasome activation and Toll-like receptor-4 signal transduction to protect against oxygen-glucose deprivation-induced inflammation in BV-2 cells. CNS Neurosci. Ther. 2014, 20, 147–153. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Y.; Xia, S.; Kong, Q.; Li, S.; Liu, X.; Junqueira, C.; Meza-Sosa, K.F.; Mok, T.M.Y.; Ansara, J.; et al. Gasdermin E suppresses tumour growth by activating anti-tumour immunity. Nature 2020, 579, 415–420. [Google Scholar] [CrossRef]
- Stetler, R.A.; Leak, R.K.; Gan, Y.; Li, P.; Zhang, F.; Hu, X.; Jing, Z.; Chen, J.; Zigmond, M.J.; Gao, Y. Preconditioning provides neuroprotection in models of CNS disease: Paradigms and clinical significance. Prog. Neurobiol. 2014, 114, 58–83. [Google Scholar] [CrossRef]
- McDonough, A.; Weinstein, J.R. Neuroimmune response in ischemic preconditioning. Neurotherapeutics 2016, 13, 748–761. [Google Scholar] [CrossRef]
- Shi, S.; Yang, W.; Tu, X.; Chen, C.; Wang, C. Ischemic preconditioning reduces ischemic brain injury by suppressing nuclear factor kappa B expression and neuronal apoptosis. Neural Regen. Res. 2013, 8, 633–638. [Google Scholar]
- Liang, J.; Luan, Y.; Lu, B.; Zhang, H.; Luo, Y.; Ge, P. Protection of ischemic postconditioning against neuronal apoptosis induced by transient focal ischemia is associated with attenuation of NF-κB/p65 activation. PLoS ONE 2014, 9, e96734. [Google Scholar] [CrossRef] [PubMed]
- Vartanian, K.; Stenzel-Poore, M. Toll-like receptor tolerance as a mechanism for neuroprotection. Transl. Stroke Res. 2010, 1, 252–260. [Google Scholar] [CrossRef]
- Wang, P.-F.; Xiong, X.-Y.; Chen, J.; Wang, Y.-C.; Duan, W.; Yang, Q.-W. Function and mechanism of toll-like receptors in cerebral ischemic tolerance: From preconditioning to treatment. J. Neuroinflamm. 2015, 12, 80. [Google Scholar] [CrossRef]
- Du, X.; Yang, J.; Liu, C.; Wang, S.; Zhang, C.; Zhao, H.; Du, H.; Geng, X. Hypoxia-inducible factor 1α and 2α have beneficial effects in remote ischemic preconditioning against stroke by modulating inflammatory responses in aged rats. Front. Aging Neurosci. 2020, 12, 54. [Google Scholar] [CrossRef]
- Bellut, M.; Papp, L.; Bieber, M.; Kraft, P.; Stoll, G.; Schuhmann, M.K. NLPR3 inflammasome inhibition alleviates hypoxic endothelial cell death in vitro and protects blood-brain barrier integrity in murine stroke. Cell Death Dis. 2021, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Qin, J.-X.; Shi, J.-Q.; Jiang, T.; Wang, F.; Xie, C.; Gao, Q.; Zhi, N.; Dong, Q.; Guan, Y.-T. Fine particulate matter exposure aggravates ischemic injury via NLRP3 inflammasome activation and pyroptosis. CNS Neurosci. Ther. 2022, 28, 1045–1058. [Google Scholar] [CrossRef] [PubMed]
- Zuurbier, C.J.; Jong, W.M.C.; Eerbeek, O.; Koeman, A.; Pulskens, W.P.; Butter, L.M.; Leemans, J.C.; Hollmann, M.W. Deletion of the innate immune NLRP3 receptor abolishes cardiac ischemic preconditioning and is associated with decreased Il-6/STAT3 signaling. PLoS ONE 2012, 7, e40643. [Google Scholar] [CrossRef]
- Pearce, L.; Davidson, S.M.; Yellon, D.M. Does remote ischaemic conditioning reduce inflammation? A focus on innate immunity and cytokine response. Basic Res. Cardiol. 2021, 116, 12. [Google Scholar] [CrossRef]
- Pang, Y.-Q.; Yang, J.; Jia, C.-M.; Zhang, R.; Pang, Q. Hypoxic preconditioning reduces NLRP3 inflammasome expression and protects against cerebral ischemia/reperfusion injury. Neural Regen. Res. 2022, 17, 395–400. [Google Scholar] [CrossRef]
- Zhaolin, Z.; Guohua, L.; Shiyuan, W.; Zuo, W. Role of pyroptosis in cardiovascular disease. Cell Prolif. 2019, 52, e12563. [Google Scholar] [CrossRef]
- Chen, S.; Mei, S.; Luo, Y.; Wu, H.; Zhang, J.; Zhu, J. Gasdermin family: A promising therapeutic target for stroke. Transl. Stroke Res. 2018, 9, 555–563. [Google Scholar] [CrossRef]
- Liu, J.; Zheng, J.; Xu, Y.; Cao, W.; Wang, J.; Wang, B.; Zhao, L.; Zhang, X.; Liao, W. Enriched environment attenuates pyroptosis to improve functional recovery after cerebral ischemia/reperfusion injury. Front. Aging Neurosci. 2021, 13, 717644. [Google Scholar] [CrossRef]
- Gou, X.; Xu, D.; Li, F.; Hou, K.; Fang, W.; Li, Y. Pyroptosis in stroke-new insights into disease mechanisms and therapeutic strategies. J. Physiol. Biochem. 2021, 77, 511–529. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hao, J.-H.; Yao, D.; Li, R.; Li, X.-F.; Yu, Z.-Y.; Luo, X.; Liu, X.-H.; Wang, M.-H.; Wang, W. Caspase-1 inhibition prevents neuronal death by targeting the canonical inflammasome pathway of pyroptosis in a murine model of cerebral ischemia. CNS Neurosci. Ther. 2020, 26, 925–939. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Y.; Chen, W.; Li, W.; Huang, Y.; Duan, D.; Ge, L.; He, J.; Liu, J.; Hu, Z.; Lu, M. Ischemic-hypoxic preconditioning enhances the mitochondrial function recovery of transplanted olfactory mucosa mesenchymal stem cells via miR-181a signaling in ischemic stroke. Aging 2021, 13, 11234–11256. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, L.; Sun, X.; Pan, M.; Zhang, W.; Zhu, D.; Lu, Z.; Wang, K.; Dong, Y.; Guan, Y. Ischemic Preconditioning Provides Neuroprotection by Inhibiting NLRP3 Inflammasome Activation and Cell Pyroptosis. Brain Sci. 2023, 13, 897. https://doi.org/10.3390/brainsci13060897
Gao L, Sun X, Pan M, Zhang W, Zhu D, Lu Z, Wang K, Dong Y, Guan Y. Ischemic Preconditioning Provides Neuroprotection by Inhibiting NLRP3 Inflammasome Activation and Cell Pyroptosis. Brain Sciences. 2023; 13(6):897. https://doi.org/10.3390/brainsci13060897
Chicago/Turabian StyleGao, Li, Xin Sun, Meibo Pan, Wenrui Zhang, Desheng Zhu, Zhongjiao Lu, Kan Wang, Yinfeng Dong, and Yangtai Guan. 2023. "Ischemic Preconditioning Provides Neuroprotection by Inhibiting NLRP3 Inflammasome Activation and Cell Pyroptosis" Brain Sciences 13, no. 6: 897. https://doi.org/10.3390/brainsci13060897