

Glabridin Therapy Reduces Chronic Allodynia, Spinal Microgliosis, and Dendritic Spine Generation by Inhibiting Fractalkine-CX3CR1 Signaling in a Mouse Model of Tibial Fractures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs and Administration
2.3. Mouse Model of Chronic Fracture Pain
2.4. Behavioral Testing
2.5. ELISA Analysis
2.6. Golgi Staining
2.7. Immunofluorescence
2.8. Statistical Analysis
3. Results
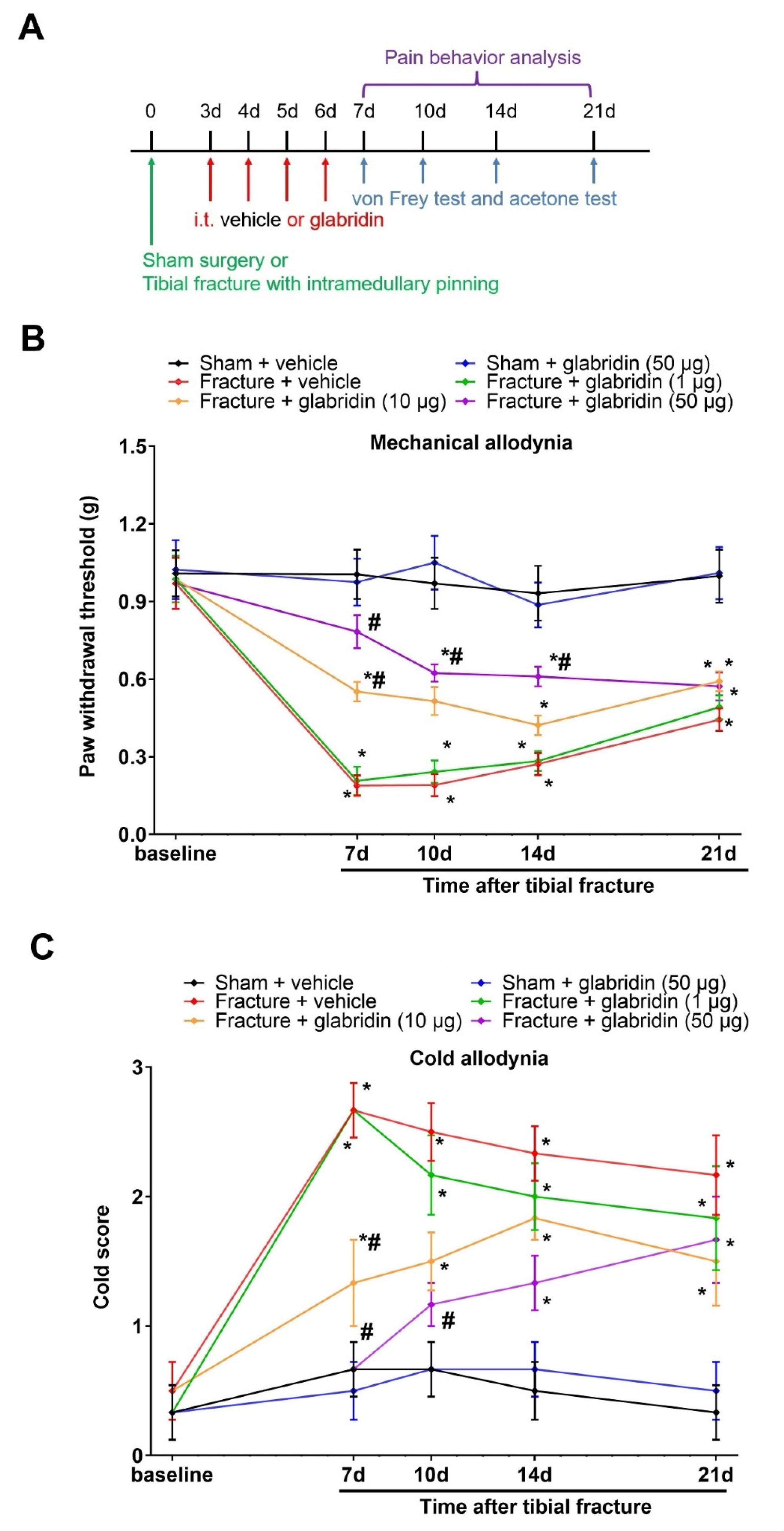
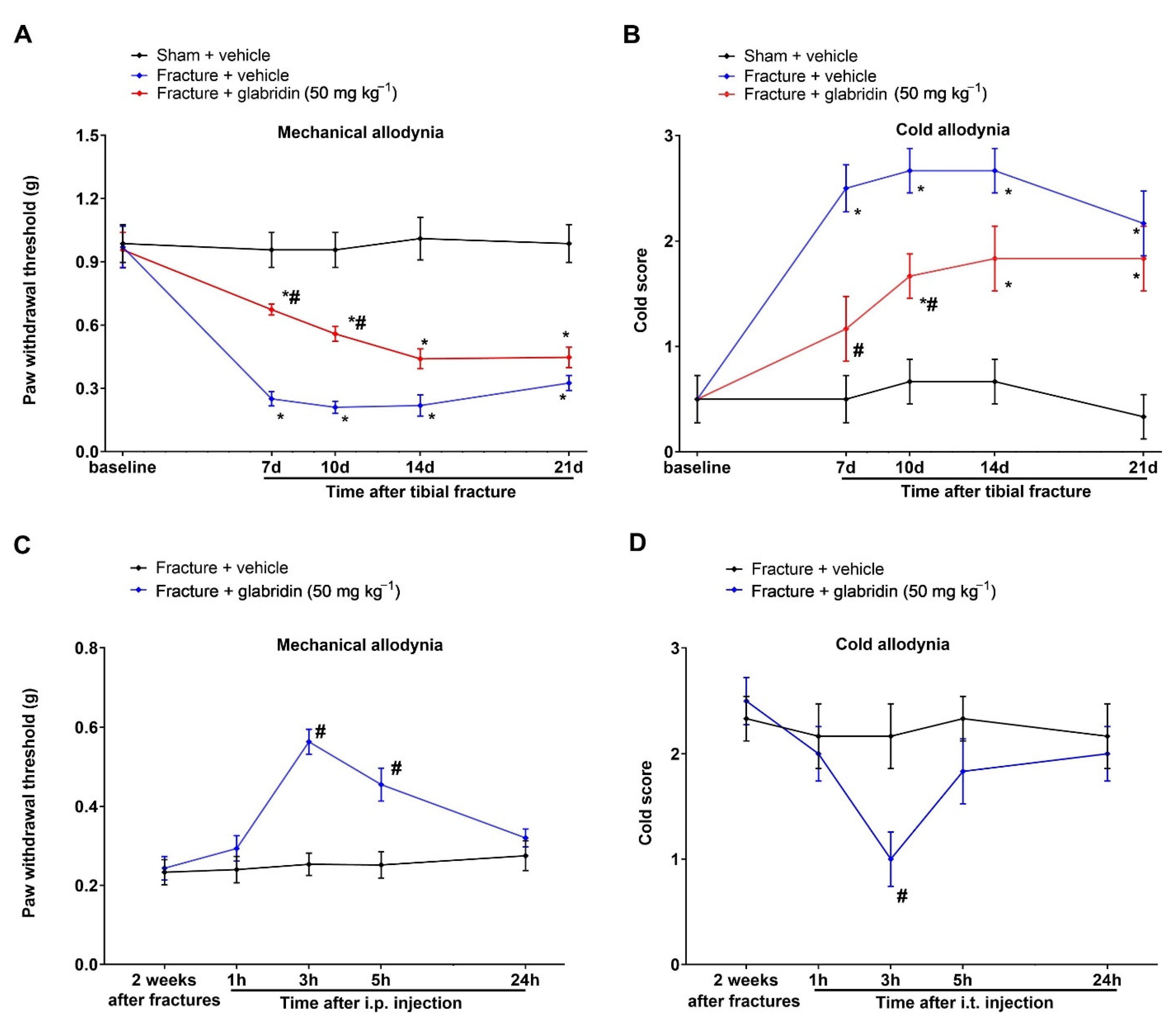
3.1. Intrathecal Pre-Administration of Glabridin Prevents Tibial Fracture-Associated Chronic Allodynia following Orthopedic Surgeries
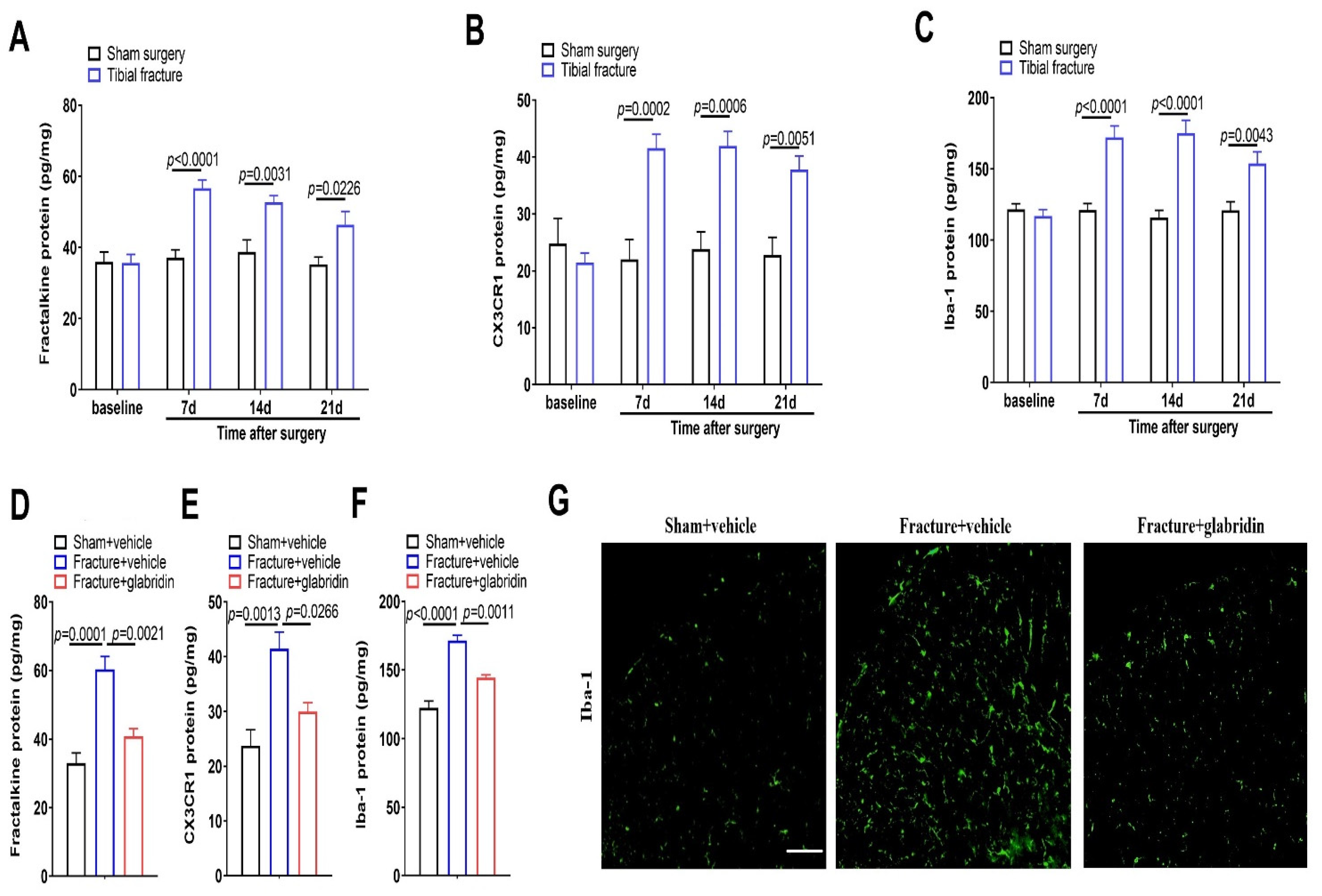
3.2. Intrathecal Glabridin Reduces Spinal Fractalkine/CX3CR1 Expression and Microgliosis following Tibial Fractures and Orthopedic Surgeries
3.3. Post-Treatment with Glabridin via Intrathecal Route Relieved the Existing Prolonged Allodynia following Orthopedic Surgeries with Tibial Fractures
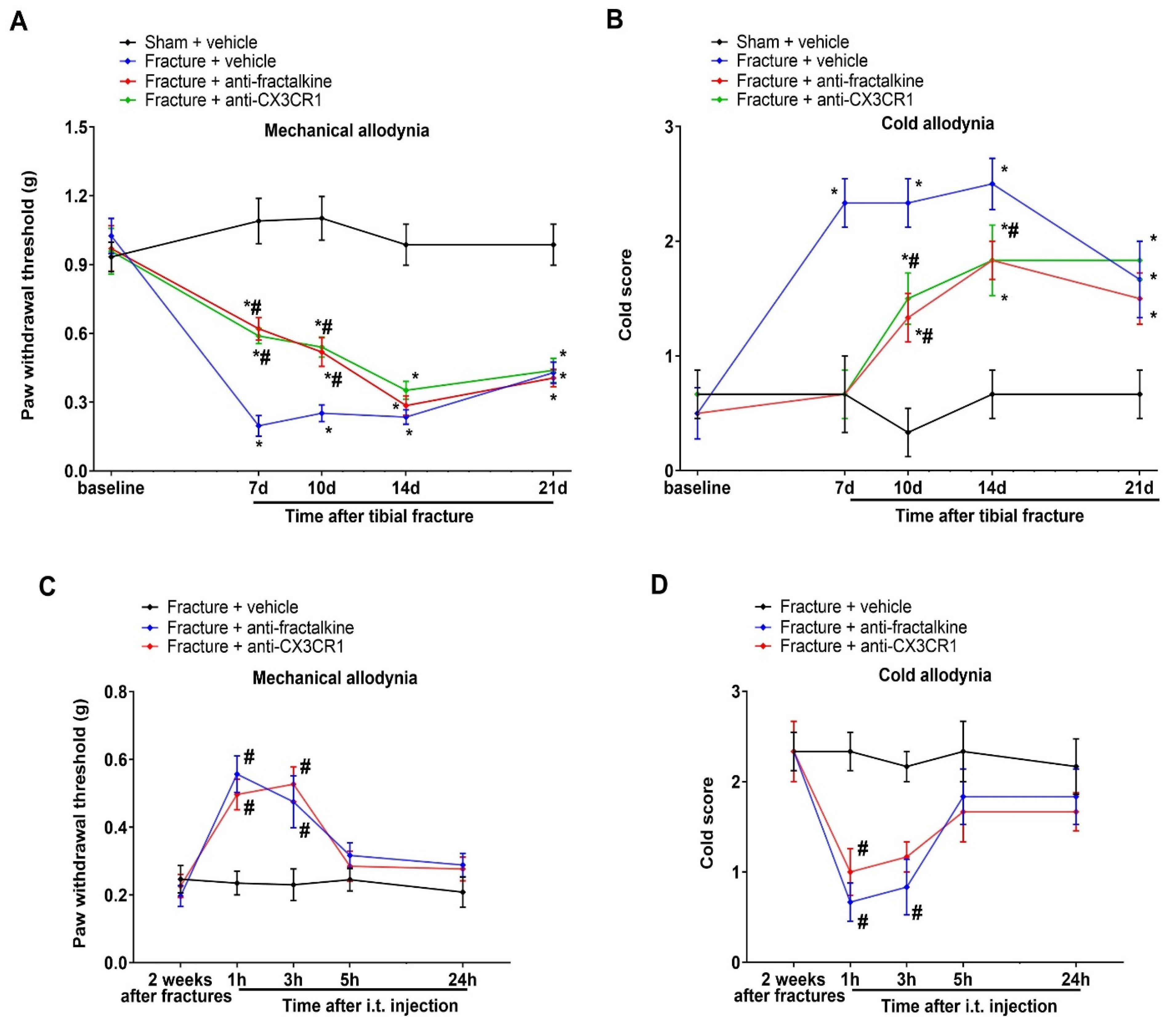
3.4. Spinal Suppression of Fractalkine/CX3CR1 Signaling Protects against Tibial Fracture-Associated Chronic Allodynia
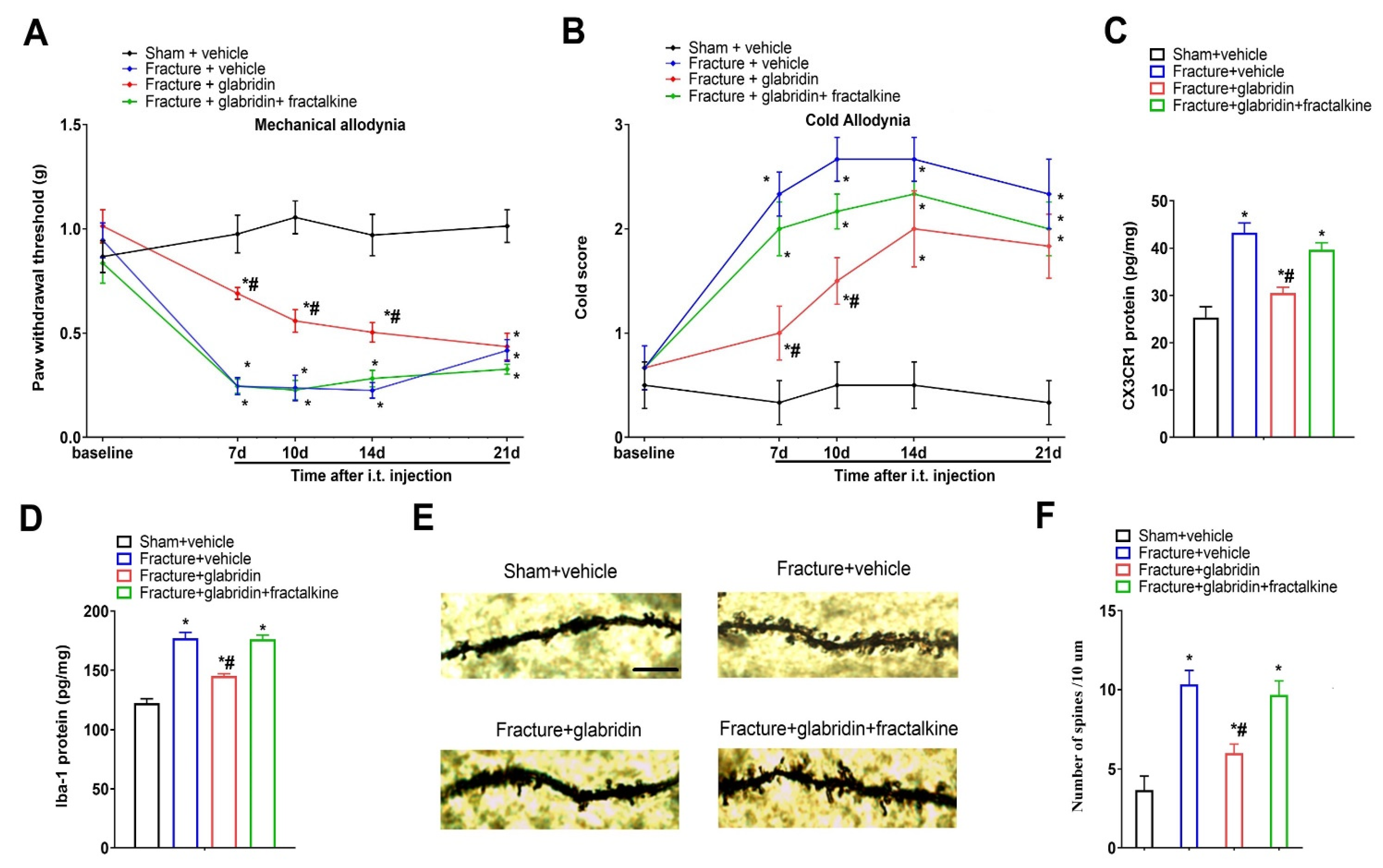
3.5. Recombinant Fractalkine Abolished Glabridin-Induced Anti-Nociception, Spinal Inhibition of CX3CR1 Expression, and Microgliosis
3.6. Glabridin Reduces Fracture-Induced Spinal Dendritic Spine Generation, Which Is Compensated by Co-Administration of Recombinant Fractalkine
3.7. Intrathecal Injection of Recombinant Fractalkine Evokes Acute Allodynia, Which Is Compensated by Co-Administration of Minocycline
3.8. Systemic Therapy of Glabridin Ameliorates Fracture-Associated Chronic Allodynia Behaviors following Orthopedic Surgeries
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, W.; Lv, H.; Liu, S.; Liu, B.; Zhu, Y.; Chen, X.; Yang, G.; Liu, L.; Zhang, T.; Wang, H.; et al. National incidence of traumatic fractures in China: A retrospective survey of 512,187 individuals. Lancet Glob. Health 2017, 5, e807–e817. [Google Scholar] [CrossRef]
- Khan, J.S.; Devereaux, P.; LeManach, Y.; Busse, J.W. Patient coping and expectations about recovery predict the development of chronic post-surgical pain after traumatic tibial fracture repair. Br. J. Anaesth. 2016, 117, 365–370. [Google Scholar] [CrossRef]
- McVeigh, L.G.; Perugini, A.J.; Fehrenbacher, J.C.; White, F.A.; Kacena, M.A. Assessment, Quantification, and Management of Fracture Pain: From Animals to the Clinic. Curr. Osteoporos. Rep. 2020, 18, 460–470. [Google Scholar] [CrossRef]
- Tajerian, M.; Leu, D.; Zou, Y.; Sahbaie, P.; Li, W.; Khan, H.; Hsu, V.; Kingery, W.; Huang, T.T.; Becerra, L.; et al. Brain Neuroplastic Changes Accompany Anxiety and Memory Deficits in a Model of Complex Regional Pain Syndrome. Anesthesiology 2014, 121, 852–865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.-D.; Barde, S.; Yang, T.; Lei, B.; Eriksson, L.I.; Mathew, J.P.; Andreska, T.; Akassoglou, K.; Harkany, T.; Hökfelt, T.G.M.; et al. Orthopedic surgery modulates neuropeptides and BDNF expression at the spinal and hippocampal levels. Proc. Natl. Acad. Sci. USA 2016, 113, E6686–E6695. [Google Scholar] [CrossRef] [PubMed]
- Baral, P.; Udit, S.; Chiu, I.M. Pain and immunity: Implications for host defence. Nat. Rev. Immunol. 2019, 19, 433–447. [Google Scholar] [CrossRef]
- Li, W.-W.; Yang, Y.; Guo, T.-Z.; Sahbaie, P.; Shi, X.-Y.; Guang, Q.; Kingery, W.S.; Herzenberg, L.A.; Clark, J.D. IL-6 signaling mediates the germinal center response, IgM production and nociceptive sensitization in male mice after tibia fracture. Brain Behav. Immun. 2021, 94, 148–158. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, H.; Li, N.; Li, J.; Zhang, L. Chronic Pain after Bone Fracture: Current Insights into Molecular Mechanisms and Therapeutic Strategies. Brain Sci. 2022, 12, 1056. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhang, Y.Q.; Qadri, Y.J.; Serhan, C.N.; Ji, R.R. Microglia in pain: Detrimental and protective roles in pathogenesis and res-olution of pain. Neuron 2018, 100, 1292–1311. [Google Scholar] [CrossRef]
- Ji, R.-R.; Nackley, A.; Huh, B.Y.; Terrando, N.; Maixner, D.W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef]
- Qiang, Z.; Yu, W. Chemokine CCL7 regulates spinal phosphorylation of GluA1-containing AMPA receptor via interleukin-18 in remifentanil-induced hyperalgesia in rats. Neurosci. Lett. 2019, 711, 134440. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Hu, L.; Qin, F.; Yin, L.; Yi, X.; Yuan, L.; Wu, W. Spinal WNT pathway contributes to remifentanil induced hyperalgesia through regulating fractalkine and CX3CR1 in rats. Neurosci. Lett. 2016, 633, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Sessler, K.; Blechschmidt, V.; Hoheisel, U.; Mense, S.; Schirmer, L.; Treede, R.-D. Spinal cord fractalkine (CX3CL1) signaling is critical for neuronal sensitization in experimental nonspecific, myofascial low back pain. J. Neurophysiol. 2021, 125, 1598–1611. [Google Scholar] [CrossRef]
- Wang, Z.; Li, L.; Bian, C.; Yang, L.; Lv, N.; Zhang, Y. Involvement of NF-κB and the CX3CR1 Signaling Network in Mechanical Al-lodynia Induced by Tetanic Sciatic Stimulation. Neurosci. Bull. 2018, 34, 64–73. [Google Scholar] [CrossRef]
- Borghi, S.M.; Fattori, V.; Pinho-Ribeiro, F.A.; Domiciano, T.P.; Miranda-Sapla, M.M.; Zaninelli, T.H.; Casagrande, R.; Pinge-Filho, P.; Pa-vanelli, W.R.; Alves-Filho, J.C.; et al. Contribution of spinal cord glial cells to L. amazonensis ex-perimental infection-induced pain in BALB/c mice. J. Neuroinflam. 2019, 16, 113. [Google Scholar] [CrossRef]
- Cui, W.; Li, Y.; Wang, Z.M.; Song, C.; Yu, Y.; Wang, G.M.; Li, J.; Wang, C.; Zhang, L. Spinal caspase-6 regulates AMPA receptor trafficking and dendritic spine plasticity through netrin-1 in postoperative pain after orthopedic surgery for tibial fracture in mice. Pain 2021, 162, 124–134. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharm. 2020, 180, 114147. [Google Scholar] [CrossRef]
- Colvin, L.A.; Bull, F.; Hales, T.G. Perioperative opioid analgesia—When is enough too much? A review of opioid-induced tolerance and hyperalgesia. Lancet 2019, 393, 1558–1568. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Brand, E.; Wang, W.; Zhao, Z. Licorice: Resources, applications in ancient and modern times. J. Ethnopharmacol. 2022, 298, 115594. [Google Scholar] [CrossRef]
- Yang, R.; Yuan, B.-C.; Ma, Y.-S.; Zhou, S.; Liu, Y. The anti-inflammatory activity of licorice, a widely used Chinese herb. Pharm. Biol. 2016, 55, 5–18. [Google Scholar] [CrossRef]
- Li, S.; Wang, Y.; Wu, M.; Younis, M.H.; Olson, A.P.; Barnhart, T.E.; Engle, J.W.; Zhu, X.; Cai, W. Spleen-Targeted Glabridin-Loaded Na-noparticles Regulate Polarization of Monocyte/Macrophage (Mo/Mφ) for the Treatment of Cerebral Ischemia-Reperfusion Injury. Adv. Mater. 2022, 34, e2204976. [Google Scholar] [CrossRef]
- Park, S.H.; Kang, J.S.; Yoon, Y.D.; Lee, K.; Kim, K.J.; Lee, K.H.; Lee, C.W.; Moon, E.Y.; Han, S.B.; Kim, B.H.; et al. Glabridin inhib-its lipopolysaccharide-induced activation of a microglial cell line, BV-2, by blocking NF-kappaB and AP-1. Phytother. Res. 2010, 24, S29–S34. [Google Scholar] [CrossRef] [PubMed]
- Parlar, A.; Arslan, S.O.; Çam, S.A. Glabridin Alleviates Inflammation and Nociception in Rodents by Activating BKCa Channels and Reducing NO Levels. Biol. Pharm. Bull. 2020, 43, 884–897. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, C.R.; Jiang, C.; Andriessen, A.S.; Wang, K.; Wang, Z.; Ding, H.; Zhao, J.; Luo, X.; Lee, M.S.; Lei, Y.L.; et al. STING controls nociception via type I interferon signalling in sensory neurons. Nature 2021, 591, 275–280. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, P.; Liu, C.; Chen, W.; Wang, P.; Jiang, L. Hydrogen-Rich Saline Attenuates Chronic Allodynia after Bone Fractures via Reducing Spinal CXCL1/CXCR2-Mediated Iron Accumulation in Mice. Brain Sci. 2022, 12, 1610. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Z.; Song, C.; Liu, H.; Li, Y.; Li, J.; Yu, Y.; Wang, G.; Cui, W. Spinal NR2B phosphorylation at Tyr1472 regulates IRE (-) DMT1-mediated iron accumulation and spine morphogenesis via kalirin-7 in tibial fracture-associated postoperative pain af-ter orthopedic surgery in female mice. Reg. Anesth. Pain Med. 2021, 46, 363–373. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, S.; Zhao, Q.; Li, Y.; Song, C.; Wang, C.; Yu, Y.; Wang, G. Spinal Protein Kinase Mζ Regulates α-Amino-3-hydroxy-5-methyl-4-isoxazolepropionic Acid Receptor Trafficking and Dendritic Spine Plasticity via Kalirin-7 in the Pathogenesis of Remifentanil-induced Postincisional Hyperalgesia in Rats. Anesthesiology 2018, 129, 173–186. [Google Scholar] [CrossRef]
- Bell, R.; Moreira, V.; Kalso, E.; Yli-Kauhaluoma, J. Liquorice for pain? Ther. Adv. Psychopharmacol. 2021, 11, 20451253211024873. [Google Scholar] [CrossRef]
- Wang, C.; Xu, R.; Wang, X.; Li, Q.; Li, Y.; Jiao, Y.; Zhao, Q.; Guo, S.; Su, L.; Yu, Y.; et al. Spinal CCL1/CCR8 regulates phosphorylation of GluA1-containing AMPA receptor in postoperative pain after tibial fracture and orthopedic surgery in mice. Neurosci. Res. 2020, 154, 20–26. [Google Scholar] [CrossRef]
- Zhang, L.; Li, N.; Zhang, H.; Wang, Y.; Gao, T.; Zhao, Y.; Wang, G.; Yu, Y.; Wang, C.; Li, Y. Artesunate therapy alleviates frac-ture-associated chronic pain after orthopedic surgery by suppressing CCL21-dependent TREM2/DAP12 inflammatory sig-naling in mice. Front. Pharmacol. 2022, 13, 894963. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Luo, C.; Bali, K.K.; Xie, R.G.; Mains, R.E.; Eipper, B.A.; Kuner, R. A role for Kalirin-7 in nociceptive sensitization via activi-ty-dependent modulation of spinal synapses. Nat. Commun. 2015, 6, 6820. [Google Scholar] [CrossRef] [PubMed]
- Stratton, H.J.; Khanna, R. Sculpting Dendritic Spines during Initiation and Maintenance of Neuropathic Pain. J. Neurosci. 2020, 40, 7578–7589. [Google Scholar] [CrossRef] [PubMed]
- Grace, P.M.; Gaudet, A.D.; Staikopoulos, V.; Maier, S.F.; Hutchinson, M.R.; Salvemini, D.; Watkins, L.R. Nitroxidative signaling mecha-nisms in pathological pain. Trends Neurosci. 2016, 39, 862–879. [Google Scholar] [CrossRef]
- Squillace, S.; Salvemini, D. Nitroxidative stress in pain and opioid-induced adverse effects: Therapeutic opportunities. Pain 2021, 163, 205–213. [Google Scholar] [CrossRef]
- Bhatt, S.; Sharma, A.; Dogra, A.; Sharma, P.; Kumar, A.; Kotwal, P.; Bag, S.; Misra, P.; Singh, G.; Kumar, A.; et al. Glabridin attenuates paracetamol-induced liver injury in mice via CYP2E1-mediated inhibition of oxidative stress. Drug Chem. Toxicol. 2022, 45, 2352–2360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, X.; Zhong, B.; Liao, Q.; Wang, X.; Xie, Y.; He, X. Review on the Diverse Biological Effects of Glabridin. Drug Des. Dev. Ther. 2023, 17, 15–37. [Google Scholar] [CrossRef]
- Li, T.; Chen, X.; Zhang, C.; Zhang, Y.; Yao, W. An update on reactive astrocytes in chronic pain. J. Neuroinflam. 2019, 16, 140. [Google Scholar] [CrossRef]
- Liu, X.; Bae, C.; Gelman, B.B.; Chung, J.M.; Tang, S.-J. A neuron-to-astrocyte Wnt5a signal governs astrogliosis during HIV-associated pain pathogenesis. Brain 2022, 145, 4108–4123. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, J.; Liu, H.; Qiu, Z.; Xiao, Z.; Lu, Z. Glabridin Therapy Reduces Chronic Allodynia, Spinal Microgliosis, and Dendritic Spine Generation by Inhibiting Fractalkine-CX3CR1 Signaling in a Mouse Model of Tibial Fractures. Brain Sci. 2023, 13, 739. https://doi.org/10.3390/brainsci13050739
Long J, Liu H, Qiu Z, Xiao Z, Lu Z. Glabridin Therapy Reduces Chronic Allodynia, Spinal Microgliosis, and Dendritic Spine Generation by Inhibiting Fractalkine-CX3CR1 Signaling in a Mouse Model of Tibial Fractures. Brain Sciences. 2023; 13(5):739. https://doi.org/10.3390/brainsci13050739
Chicago/Turabian StyleLong, Juan, Hongbing Liu, Zhimin Qiu, Zhong Xiao, and Zhongqiu Lu. 2023. "Glabridin Therapy Reduces Chronic Allodynia, Spinal Microgliosis, and Dendritic Spine Generation by Inhibiting Fractalkine-CX3CR1 Signaling in a Mouse Model of Tibial Fractures" Brain Sciences 13, no. 5: 739. https://doi.org/10.3390/brainsci13050739