
Cortical Layer Markers Expression and Increased Synaptic Density in Interstitial Neurons of the White Matter from Drug-Resistant Epilepsy Patients
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Sample Collection
2.2. Immunohistochemical Staining
2.3. Image Acquisition and Data Analysis
3. Results
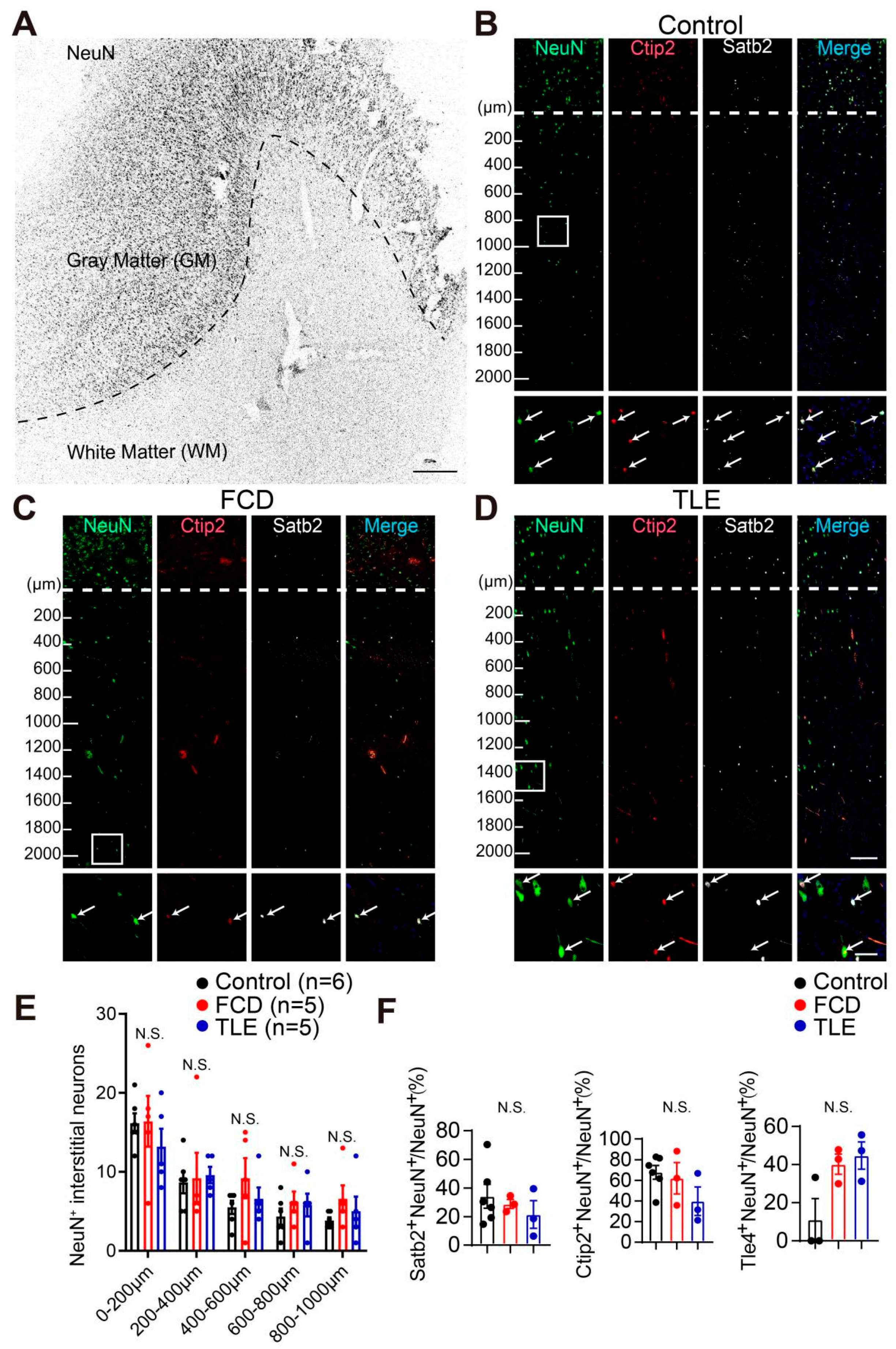
3.1. Cortical Layer Marker Expression in Interstitial Neurons of the White Matter from the Adult Human Brain
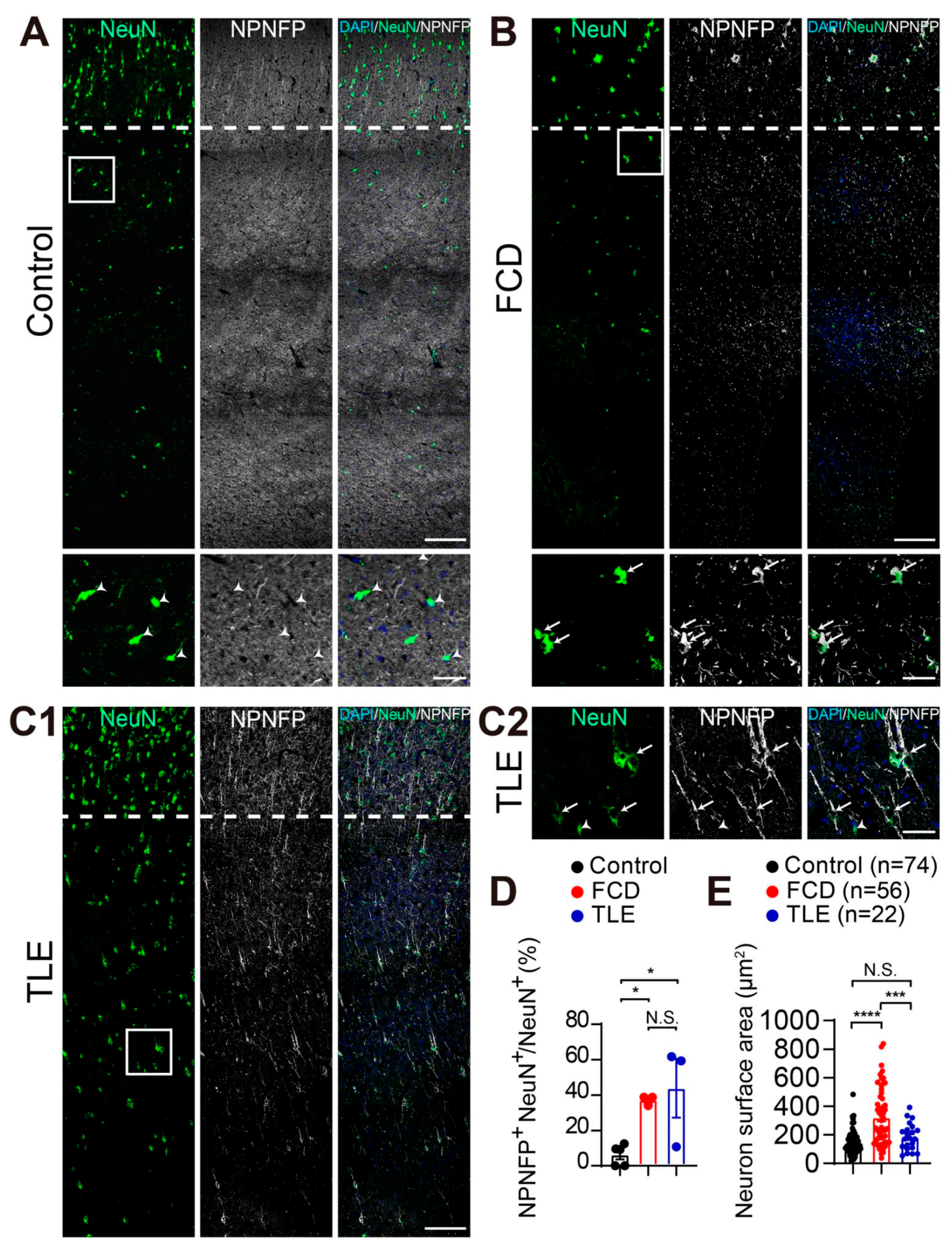
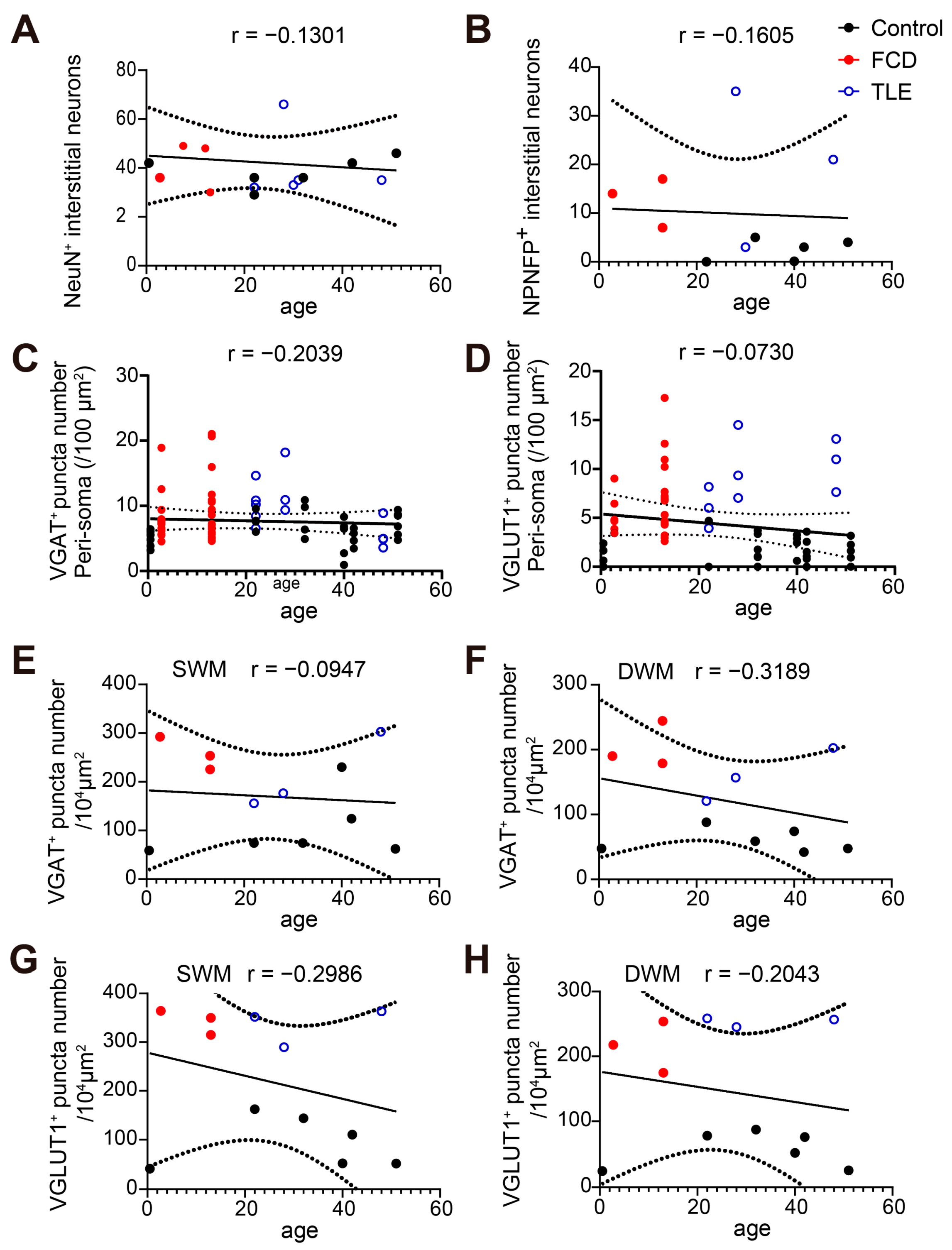
3.2. Increased Non-Phosphorylated Neurofilament Protein (NPNFP) Expression in Interstitial Neurons in the White Matter of Drug-Resistant Epilepsy Patients
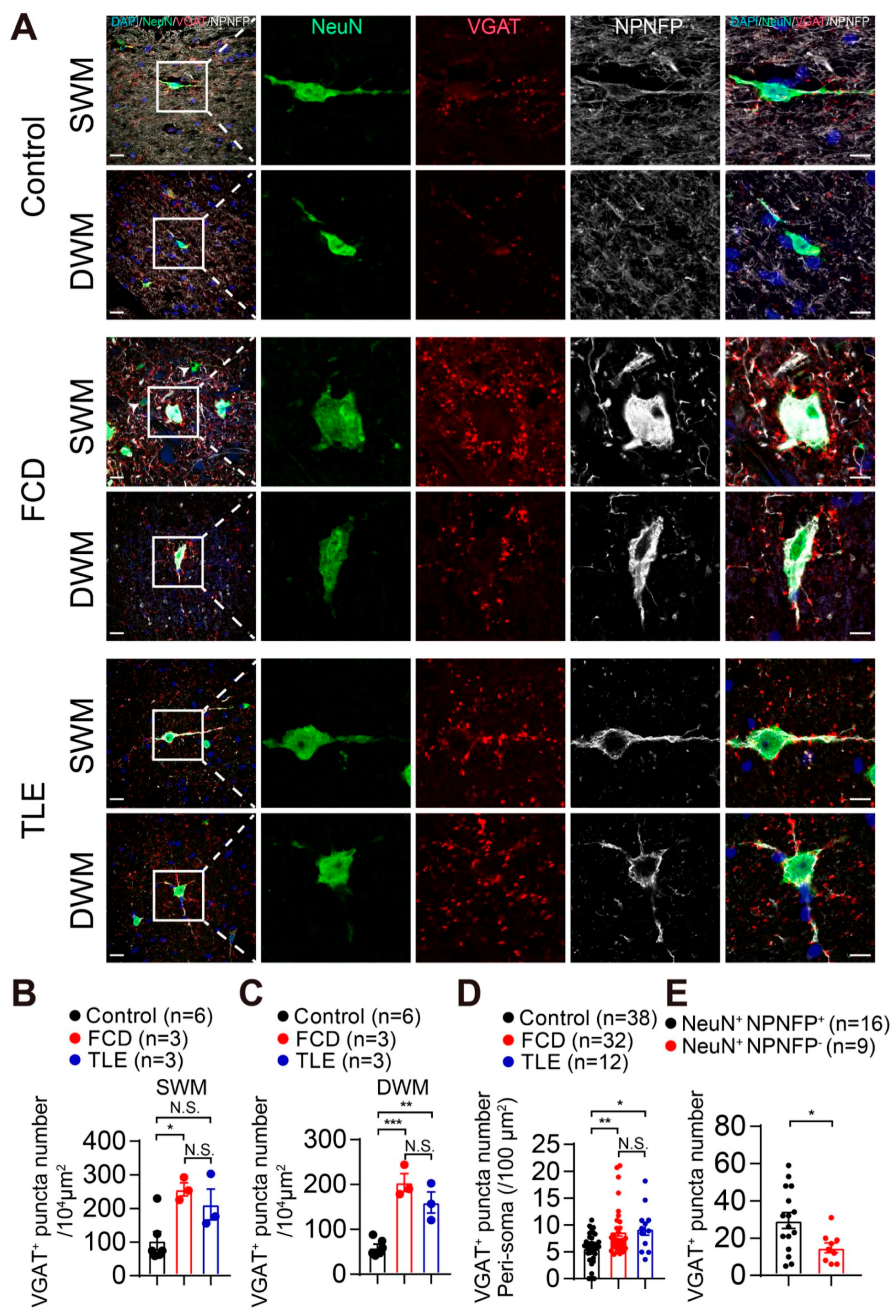
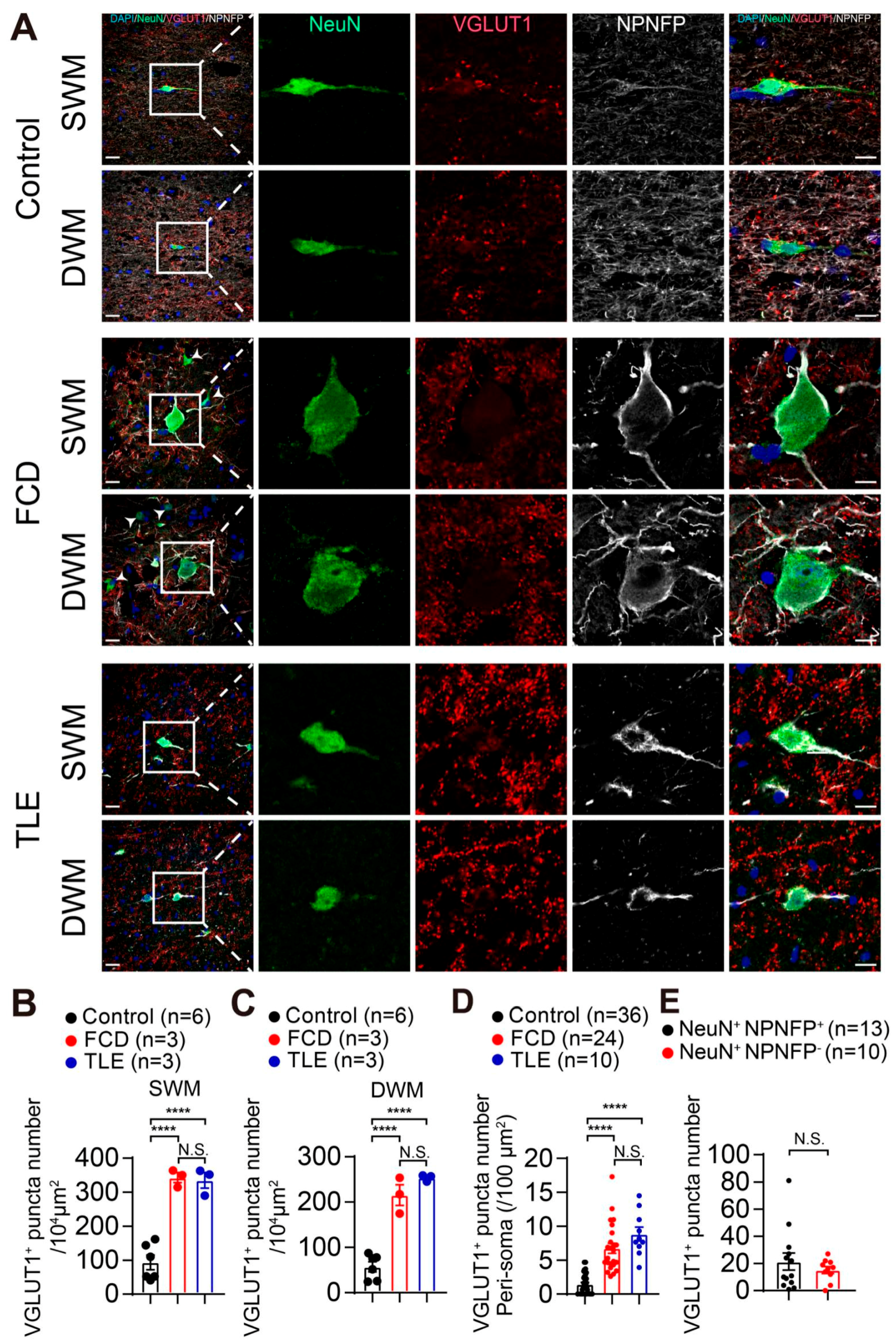
3.3. Increased GABAergic and Glutamatergic Synaptic Density in the White Matter of Drug-Resistant Epilepsy Patients
4. Discussion
5. Conclusions
6. Limitations and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Satb2 | special AT-rich sequence-binding protein 2 |
| Ctip2 | COUP-TF-interacting protein 2 |
| Tle4 | transducin-like enhancer protein 4 |
| NPNFP | non-phosphorylated neurofilament protein |
| GABA | γ-aminobutyric acid |
| MAP2 | microtubule associated protein 2 |
| SST | somatostatin |
| NPY | neuropeptide Y |
| PBS | phosphate-buffered saline |
| NOS | nitric oxide synthase |
| AChE | acetylcholine esterase |
| NeuN | neuronal nuclei antigen |
| VGAT | vesicular GABA transporter |
| VGLUT1 | vesicular glutamate transporter 1 |
| DAPI | 4′, 6-diamidino-2-phenylindole |
| MRI | magnetic resonance imaging |
| FCD | focal cortical dysplasia |
| TLE | temporal lobe epilepsy |
| SWM | superficial white matter |
| DWM | deep white matter |
| ROI | region of interest |
| SEM | standard error of the mean |
References
- Cajal, S.R. Beitrag zum Studium der Medulla oblongata, des Kleinhirns und des Ursprungs der Gehirnnerven; Deutsche vom Verfasser erweiterte Ausgabe besorgt von Johannes Bresler, mit einem Vorwort von E. Mendel. ch. III: Kleinhirnrinde; Verlag von Johann Ambrosius Barth: Leipzig, German, 1896; pp. 21–25. [Google Scholar]
- Kostovic, I.; Rakic, P. Cytology and time of origin of interstitial neurons in the white matter in infant and adult human and monkey telencephalon. J. Neurocytol. 1980, 9, 219–242. [Google Scholar] [CrossRef]
- Judas, M.; Sedmak, G.; Pletikos, M. Early history of subplate and interstitial neurons: From Theodor Meynert (1867) to the discovery of the subplate zone (1974). J. Anat. 2010, 217, 344–367. [Google Scholar] [CrossRef] [PubMed]
- Sedmak, G.; Judas, M. White Matter Interstitial Neurons in the Adult Human Brain: 3% of Cortical Neurons in Quest for Recognition. Cells 2021, 10, 190. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Sola, M.L.; GonzalezDelgado, F.J.; Pueyo-Morlans, M.; Medina-Bolivar, C.; HernandezAcosta, N.C.; Gonzalez-Gomez, M.; Meyer, G. Neurons in the white matter of the adult human neocortex. Front. Neuroanat. 2009, 3, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedmak, G.; Judas, M. The total number of white matter interstitial neurons in the human brain. J. Anat. 2019, 235, 626–636. [Google Scholar] [CrossRef]
- Garcia-Marin, V.; Blazquez-Llorca, L.; Rodriguez, J.-R.; Gonzalez-Soriano, J.; DeFelipe, J. Differential distribution of neurons in the gyral white matter of the human cerebral cortex. J. Comp. Neurol. 2010, 518, 4740–4759. [Google Scholar] [CrossRef] [PubMed]
- Judas, M.; Sedmak, G.; Pletikos, M.; Jovanov-Milosevic, N. Populations of subplate and interstitial neurons in fetal and adult human telencephalon. J. Anat. 2010, 217, 381–399. [Google Scholar] [CrossRef]
- Chun, J.J.; Shatz, C.J. Interstitial cells of the adult neocortical white matter are the remnant of the early generated subplate neuron population. J. Comp. Neurol. 1989, 282, 555–569. [Google Scholar] [CrossRef]
- Hoerder-Suabedissen, A.; Wang, W.Z.; Lee, S.; Davies, K.E.; Goffinet, A.M.; Rakić, S.; Parnavelas, J.; Reim, K.; Nicolić, M.; Paulsen, O.; et al. Novel markers reveal subpopulations of subplate neurons in the murine cerebral cortex. Cereb. Cortex 2009, 19, 1738–1750. [Google Scholar] [CrossRef] [PubMed]
- Hoerder-Suabedissen, A.; Molnár, Z. Molecular diversity of early-born subplate neurons. Cereb. Cortex 2013, 23, 1473–1483. [Google Scholar] [CrossRef] [Green Version]
- Banovac, I.; Sedmak, D.; Esclapez, M.; Petanjek, Z. The Distinct Characteristics of Somatostatin Neurons in the Human Brain. Mol. Neurobiol. 2022, 59, 4953–4965. [Google Scholar] [CrossRef] [PubMed]
- Meyer, G.; Wahle, P.; Castañeyra-Perdomo, A.; Ferres-Torres, R. Morphology of neurons in the white matter of the adult human neocortex. Exp. Brain Res. 1992, 88, 204–212. [Google Scholar] [CrossRef]
- Soki, N.; Richter, Z.; Karadi, K.; Lorincz, K.; Horvath, R.; Gyimesi, C.; Szekeres-Paraczky, C.; Horvath, Z.; Janszky, J.; Doczi, T.; et al. Investigation of synapses in the cortical white matter in human temporal lobe epilepsy. Brain Res. 2022, 1779, 147787. [Google Scholar] [CrossRef]
- Iffland, P.H.; Crino, P.B. Focal Cortical Dysplasia: Gene Mutations, Cell Signaling, and Therapeutic Implications. Annu. Rev. Pathol. 2017, 12, 547–571. [Google Scholar] [CrossRef]
- Zeng, H.; Shen, E.H.; Hohmann, J.G.; Oh, S.W.; Bernard, A.; Royall, J.J.; Glattfelder, K.J.; Sunkin, S.M.; Morris, J.A.; Guillozet-Bongaarts, A.L.; et al. Large-scale cellular-resolution gene profiling in human neocortex reveals species-specific molecular signatures. Cell 2012, 149, 483–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britanova, O.; Akopov, S.; Lukyanov, S.; Gruss, P.; Tarabykin, V. Novel transcription factor Satb2 interacts with matrix attachment region DNA elements in a tissue-specific manner and demonstrates cell-type-dependent expression in the developing mouse CNS. Eur. J. Neurosci. 2005, 21, 658–668. [Google Scholar] [CrossRef]
- Molyneaux, B.J.; Arlotta, P.; Menezes, J.R.; Macklis, J.D. Neuronal subtype specification in the cerebral cortex. Nat. Rev. Neurosci. 2007, 8, 427–437. [Google Scholar] [CrossRef]
- Hevner, R.F. Layer-specific markers as probes for neuron type identity in human neocortex and malformations of cortical development. J. Neuropathol. Exp. Neurol. 2007, 66, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, T.; Hanai, S.; Takashima, S.; Nakagawa, E.; Okazaki, S.; Inoue, T.; Miyata, R.; Hoshino, K.; Akashi, T.; Sasaki, M.; et al. Neocortical layer formation of human developing brains and lissencephalies: Consideration of layer-specific marker expression. Cereb. Cortex 2011, 21, 588–596. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Ge, Q.; Yang, J.; Wang, M.; Zhou, Y.; Guo, J.-X.; Zhu, M.; Shi, J.; Hu, Y.; Shen, L.; et al. Pathological Networks Involving Dysmorphic Neurons in Type II Focal Cortical Dysplasia. Neurosci. Bull. 2022, 38, 1007–1024. [Google Scholar] [CrossRef]
- Ljungberg, M.C.; Bhattacharjee, M.B.; Lu, Y.; Armstrong, D.L.; Yoshor, D.; Swann, J.W.; Sheldon, M.; D’Arcangelo, G. Activation of Mammalian Target of Rapamycin in Cytomegalic Neurons of Human Cortical Dysplasia. Ann. Neurol. 2006, 60, 420–429. [Google Scholar] [CrossRef]
- Rossini, L.; Villani, F.; Granata, T.; Tassi, L.; Tringali, G.; Cardinale, F.; Aronica, E.; Spreafico, R.; Garbelli, R. FCD Type II and mTOR pathway: Evidence for different mechanisms involved in the pathogenesis of dysmorphic neurons. Epilepsy Res. 2017, 129, 146–156. [Google Scholar] [CrossRef]
- Rossini, L.; Medici, V.; Tassi, L.; Cardinale, F.; Tringali, G.; Bramerio, M.; Villani, F.; Spreafico, R.; Garbelli, R. Layer-specific gene expression in epileptogenic type II focal cortical dysplasia: Normal-looking neurons reveal the presence of a hidden laminar organization. Acta Neuropathol. Commun. 2014, 2, 45. [Google Scholar] [CrossRef] [PubMed]
- Cajal, S.R. Histologie du Systeme Nerveux de l’Homme & des vertébrés; Maloine: Paris, France, 1911; (Reprinted in 2 volumes by Consejo Superior de Investigaciones Cientificas, Madrid, 1955). [Google Scholar]
- Kostovic, I.; Rakic, P. Developmental history of the transient subplate zone in the visual and somatosensory cortex of the macaque monkey and human brain. J. Comp. Neurol. 1990, 297, 441–470. [Google Scholar] [CrossRef] [PubMed]
- Allendoerfer, K.L.; Shatz, C.J. The subplate, a transient neocortical structure: Its role in the development of connections between thalamus and cortex. Annu. Rev. Neurosci. 1994, 17, 185–218. [Google Scholar] [CrossRef]
- Judas, M.; Sestan, N.; Kostovic, I. Nitrinergic neurons in the developing and adult human telencephalon: Transient and permanent patterns of expression in comparison to other mammals. Microsc. Res. Tech. 1999, 45, 401–419. [Google Scholar] [CrossRef]
- Kostovic, I.; Judas, M.; Sedmak, G. Developmental history of the subplate zone, subplate neurons and interstitial white matter neurons: Relevance for schizophrenia. Int. J. Dev. Neurosci. 2011, 29, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Miškić, T.; Kostović, I.; Rašin, M.R.; Krsnik, Ž. Adult Upper Cortical Layer Specific Transcription Factor CUX2 Is Expressed in Transient Subplate and Marginal Zone Neurons of the Developing Human Brain. Cells 2021, 10, 415. [Google Scholar] [CrossRef]
- Kubo, K.I.; Deguchi, K.; Nagai, T.; Ito, Y.; Yoshida, K.; Endo, T.; Benner, S.; Shan, W.; Kitazawa, A.; Aramaki, M.; et al. Association of impaired neuronal migration with cognitive deficits in extremely preterm infants. JCI Insight 2017, 2, e88609. [Google Scholar] [CrossRef]
- Rojiani, A.M.; Emery, J.A.; Anderson, K.J.; Massey, J.K. Distribution of heterotopic neurons in normal hemispheric white matter: A morphometric analysis. J. Neuropathol. Exp. Neurol. 1996, 55, 178–183. [Google Scholar] [CrossRef] [Green Version]
- Mischel, P.S.; Nguyen, L.P.; Vinters, H.V. Cerebral cortical dysplasia associated with pediatric epilepsy. Review of neuropathologic features and proposal for a grading system. J. Neuropathol. Exp. Neurol. 1995, 54, 137–153. [Google Scholar] [CrossRef]
- Taylor, D.C.; Falconer, M.A.; Bruton, C.J.; Corsellis, J.A. Focal dysplasia of the cerebral cortex in epilepsy. J. Neurol. Neurosurg. Psychiatry 1971, 34, 369–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, H.K.; Zentner, J.; Hufnagel, A.; Campos, M.G.; Schramm, J.; Elger, C.E.; Wiestler, O.D. Surgical pathology of chronic epileptic seizure disorders: Experience with 63 specimens from extratemporal corticectomies, lobectomies and functional hemispherectomies. Acta Neuropathol. 1993, 86, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Akbarian, S.; Kim, J.J.; Potkin, S.G.; Hetrick, W.P.; Bunney, W.E.; Jones, E.G. Maldistribution of interstitial neurons in prefrontal white matter of the brains of schizophrenic patients. Arch. Gen. Psychiatry 1996, 53, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Beasley, C.L.; Cotter, D.R.; Everall, I.P. Density and distribution of white matter neurons in schizophrenia, bipolar disorder and major depressive disorder: No evidence for abnormalities of neuronal migration. Mol. Psychiatry 2002, 7, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Kanold, P.O.; Shatz, C.J. Subplate neurons regulate maturation of cortical inhibition and outcome of ocular dominance plasticity. Neuron 2006, 51, 627–638. [Google Scholar] [CrossRef] [Green Version]
- Torres-Reveron, J.; Friedlander, M.J. Properties of persistent postnatal cortical subplate neurons. J. Neurosci. 2007, 27, 9962–9974. [Google Scholar] [CrossRef] [Green Version]
- Clancy, B.; Silva-Filho, M.; Friedlander, M.J. Structure and projections of white matter neurons in the postnatal rat visual cortex. J. Comp. Neurol. 2001, 434, 233–252. [Google Scholar] [CrossRef]
- von Engelhardt, J.; Khrulev, S.; Eliava, M.; Wahlster, S.; Monyer, H. 5-HT(3A) receptor-bearing white matter interstitial GABAergic interneurons are functionally integrated into cortical and subcortical networks. J. Neurosci. 2011, 31, 16844–16854. [Google Scholar] [CrossRef] [Green Version]
- Cepeda, C.; André, V.M.; Hauptman, J.S.; Yamazaki, I.; Huynh, M.N.; Chang, J.W.; Chen, J.Y.; Fisher, R.S.; Vinters, H.V.; Levine, M.S.; et al. Enhanced GABAergic network and receptor function in pediatric cortical dysplasia Type IIB compared with Tuberous Sclerosis Complex. Neurobiol. Dis. 2012, 45, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Alfano, V.; Romagnolo, A.; Mills, J.D.; Cifelli, P.; Gaeta, A.; Morano, A.; Mühlebner, A.; Aronica, E.; Palma, E.; Ruffolo, G. Unexpected Effect of IL-1β on the Function of GABAA Receptors in Pediatric Focal Cortical Dysplasia. Brain Sci. 2022, 12, 807. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Diagnosis | Age at Epilepsy Onset | Epilepsy Duration | Age at Surgery | Monthly Seizure Frequency | Site of Surgery |
|---|---|---|---|---|---|---|
| FCD 15 | FCD IIB | 9.25 | 10 | 13.75 | * | R.gfs |
| FCD 21 | FCD IIB | 6.75 | 0.75 | 7.5 | 45 | L.gt |
| FCD 24 | FCD IIB | 0.75 | 2 | 2.75 | * | R.gf |
| FCD 25 | FCD IIB | 3 | 10 | 13 | 90 | R.gfs |
| FCD 27 | FCD IIB | 11 | 5 | 16 | 45 | L.gfs |
| TLE 31 | Crypto | 10 | 18 | 28 | 3 | R.gt |
| TLE 32 | Crypto | 12 | 10 | 22 | 15.3 | R.gt |
| TLE 36 | Crypto | 44 | 4 | 48 | 6 | L.gt |
| TLE 37 | HS | 9 | 7 | 16 | 4 | L.gt |
| TLE 40 | HS | 7.25 | 6 | 13.25 | 4.5 | R.gt |
| TLE 41 | HS | 13.25 | 0.75 | 14 | 1.5 | L.gt |
| Autopsy | Age at Death | Postmortem Delay (min) | ||||
| 2017CBB047 | Autopsy | 0.5 | 253 | / | / | R.gfs 3 |
| 2018CBB037 | Autopsy | 42 | 768 | / | / | R.gfs 3 |
| 2019CBB013 | Autopsy | 51 | 473 | / | / | R.gfs 3 |
| 2022CBB015 | Autopsy | 22 | 805 | / | / | R.gfs 3 |
| 2020CBB017 | Autopsy | 40 | 621 | / | / | R.gfs 3 |
| 2021CBB052 | Autopsy | 32 | 220 | / | / | R.gfs 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Wang, M.; Lv, Y.; Chen, J. Cortical Layer Markers Expression and Increased Synaptic Density in Interstitial Neurons of the White Matter from Drug-Resistant Epilepsy Patients. Brain Sci. 2023, 13, 626. https://doi.org/10.3390/brainsci13040626
Yang J, Wang M, Lv Y, Chen J. Cortical Layer Markers Expression and Increased Synaptic Density in Interstitial Neurons of the White Matter from Drug-Resistant Epilepsy Patients. Brain Sciences. 2023; 13(4):626. https://doi.org/10.3390/brainsci13040626
Chicago/Turabian StyleYang, Jiachao, Mi Wang, Yang Lv, and Jiadong Chen. 2023. "Cortical Layer Markers Expression and Increased Synaptic Density in Interstitial Neurons of the White Matter from Drug-Resistant Epilepsy Patients" Brain Sciences 13, no. 4: 626. https://doi.org/10.3390/brainsci13040626