
Rapamycin Affects the Hippocampal SNARE Complex to Alleviate Cognitive Dysfunction Induced by Surgery in Aged Rats
 , , and
, , and 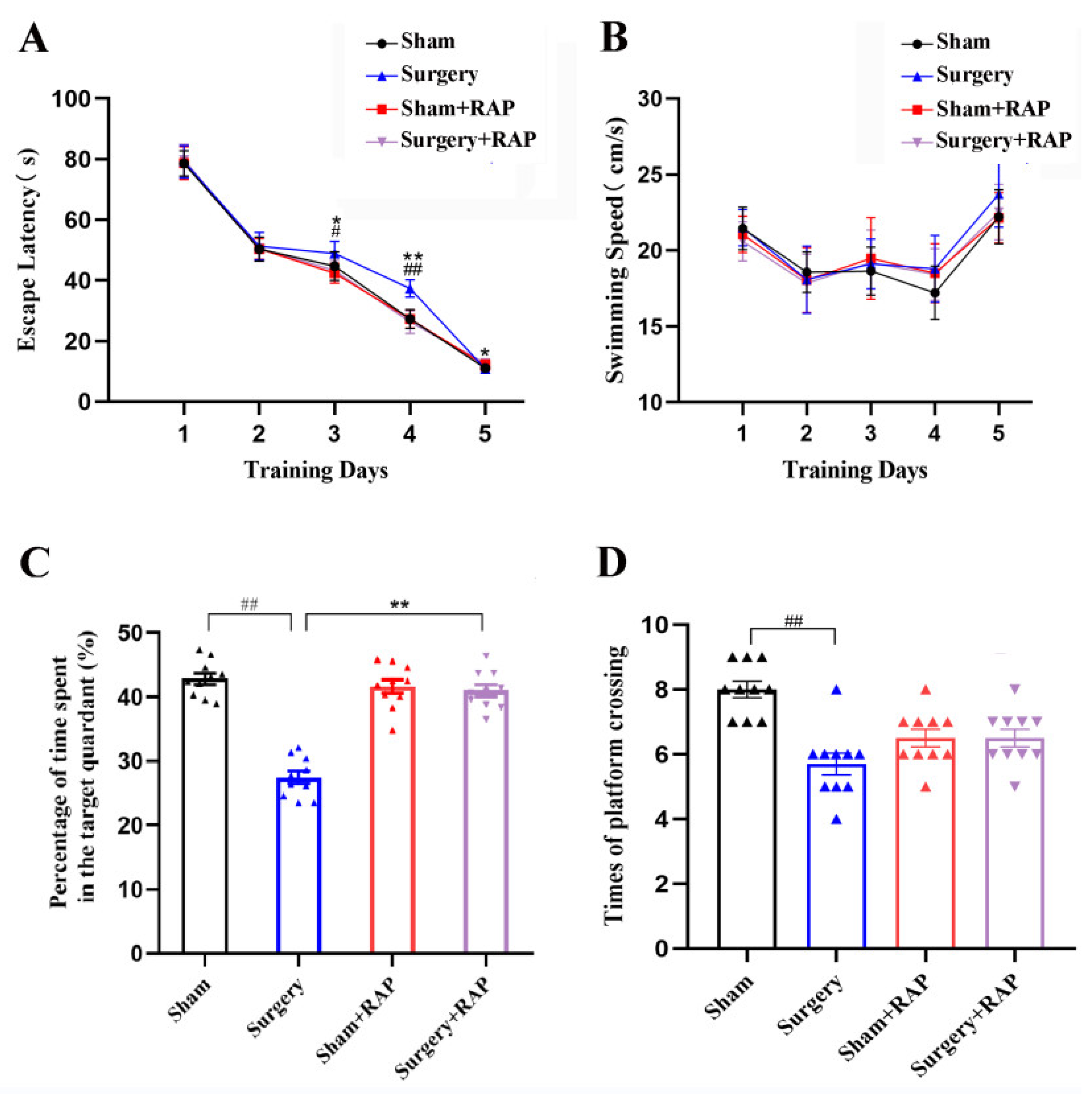{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethics
2.2. Anesthesia plus Surgery Protocol
2.3. Experimental Grouping and Treatment Scheme
2.4. Arterial Blood Gas and Blood Glucose Analysis
2.5. Morris Water Maze Test
2.6. Euthanasia and Sample Collection
2.7. Synaptosome Extraction
2.8. Western Blotting
2.9. Evaluation of Neurotransmitter Levels
2.10. Statistical Analysis
3. Results
3.1. Rapamycin Mitigated Neurobehavioral Deficits Induced by Surgery in Older Rats
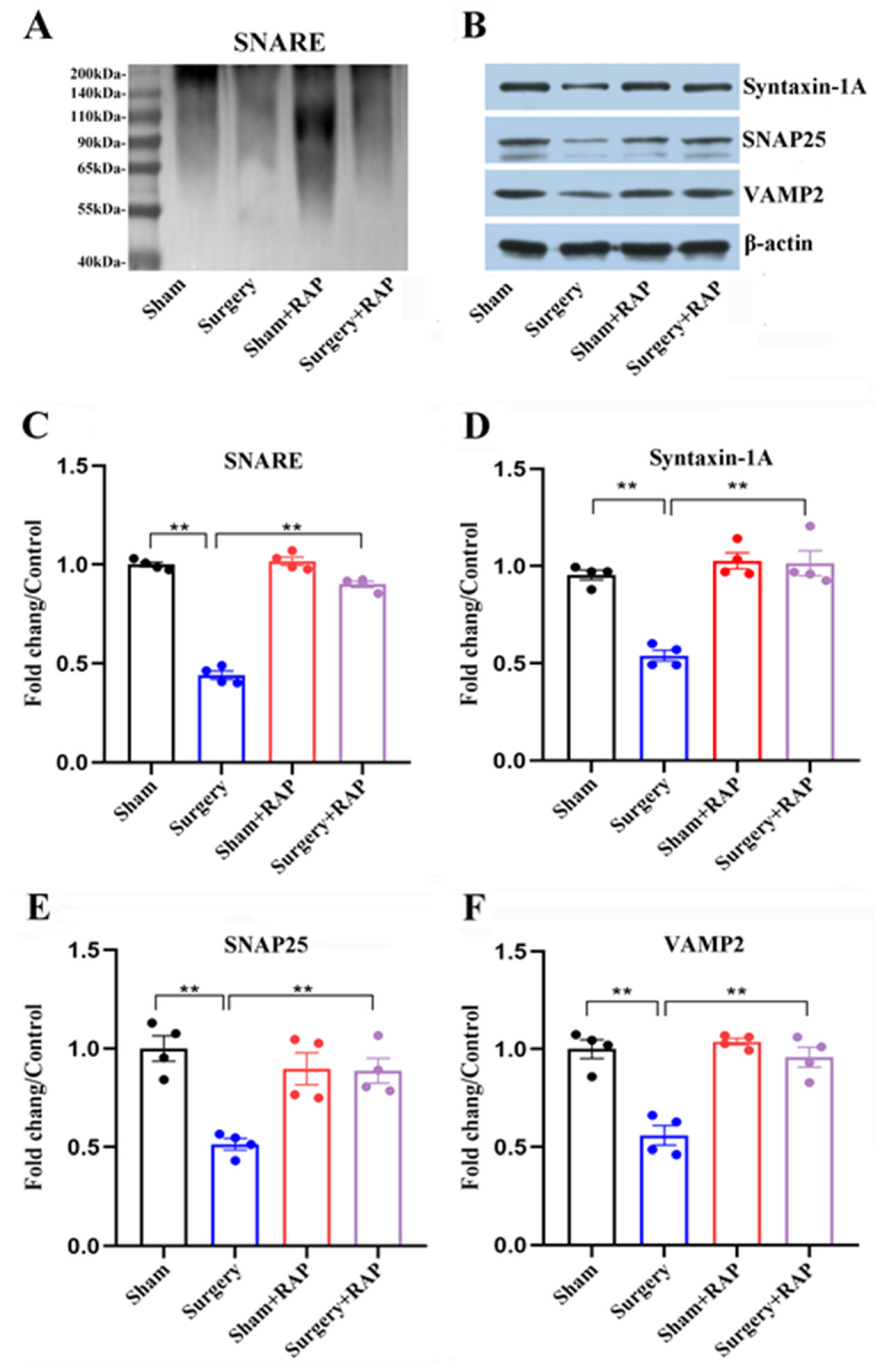
3.2. Rapamycin Ameliorated Dysregulation of SNARE and SNARE-Core Proteins Induced by Surgery
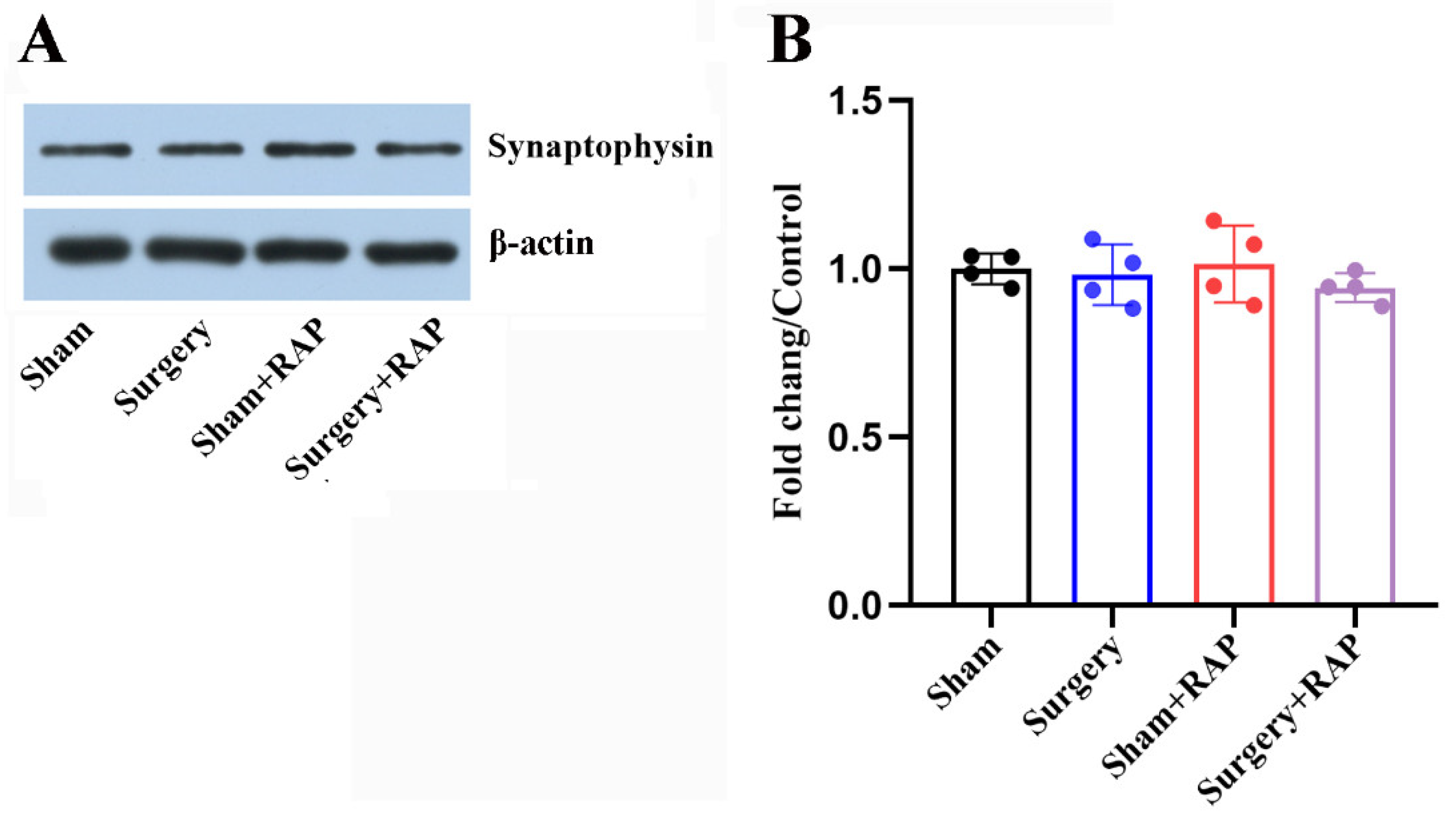
3.3. Exploratory Laparotomy Had no Effect on the Levels of Synaptic Vesicle-Associated Proteins
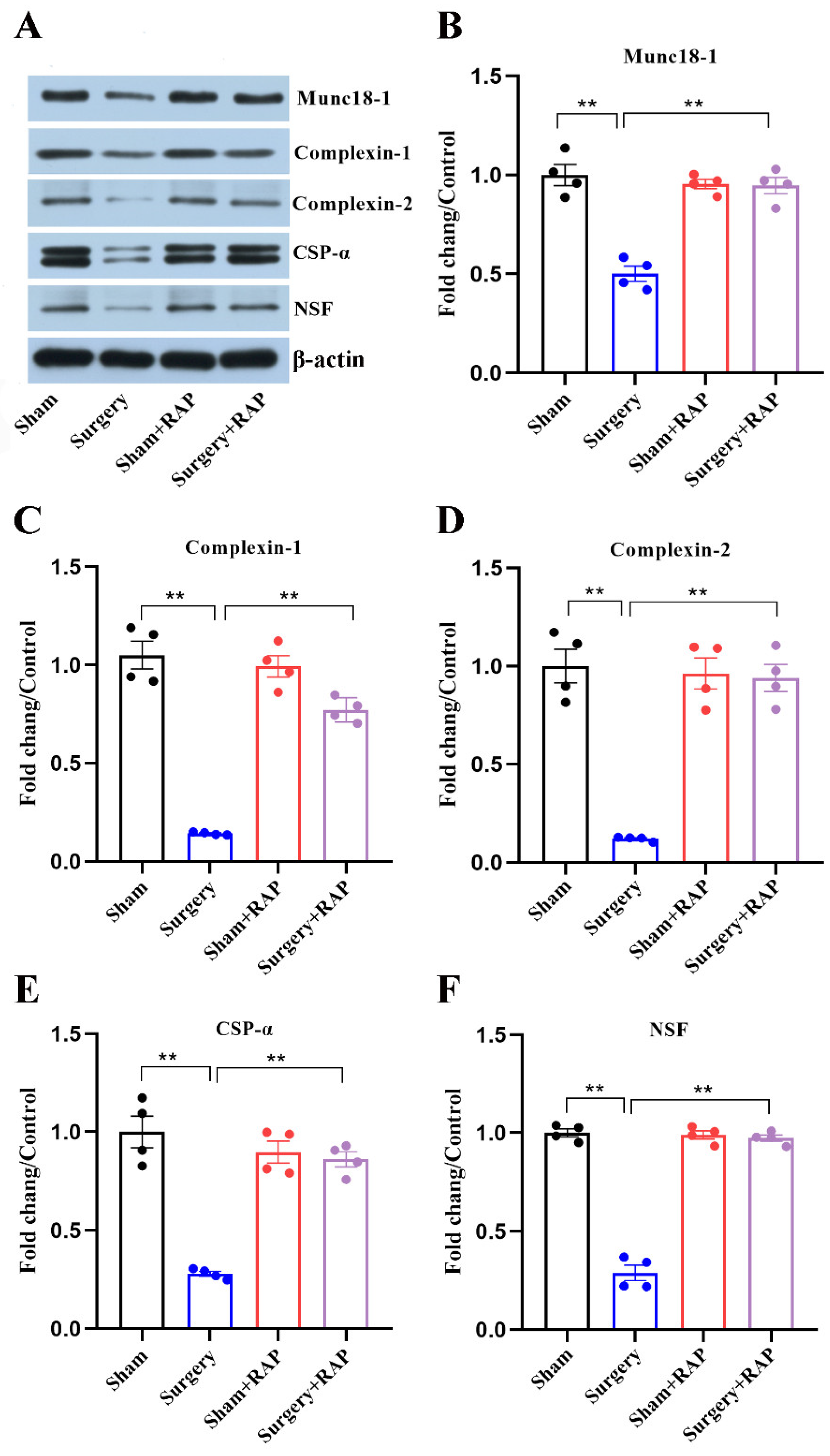
3.4. Rapamycin Rescued Disruption of SNARE Chaperones Caused by Surgery
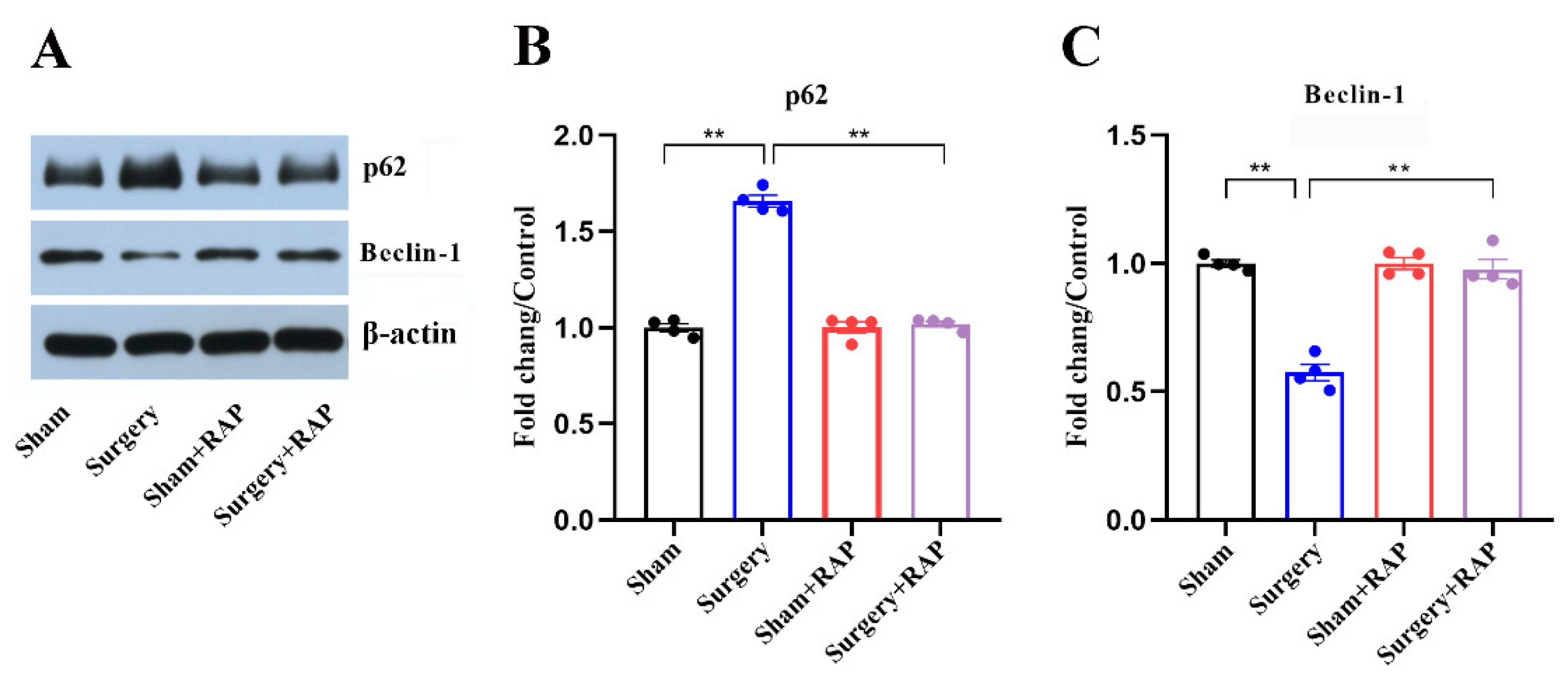
3.5. Rapamycin Ameliorated Suppressed Autophagy in the Hippocampus of Aged Rats
3.6. Rapamycin Reverses Surgical Stress-Induced Imbalances of Neurotransmitters
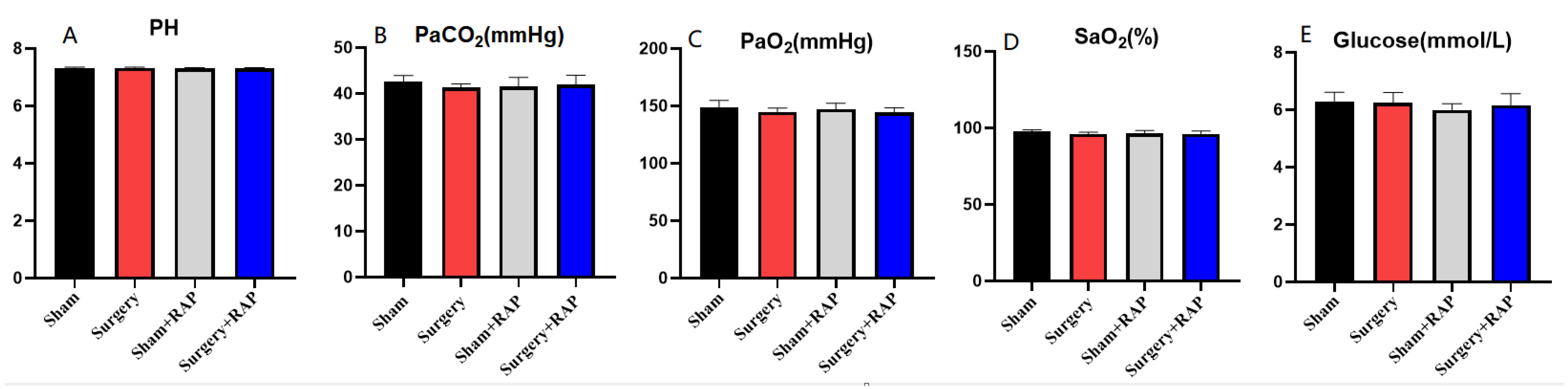
3.7. Arterial Gas and Glucose Analysis
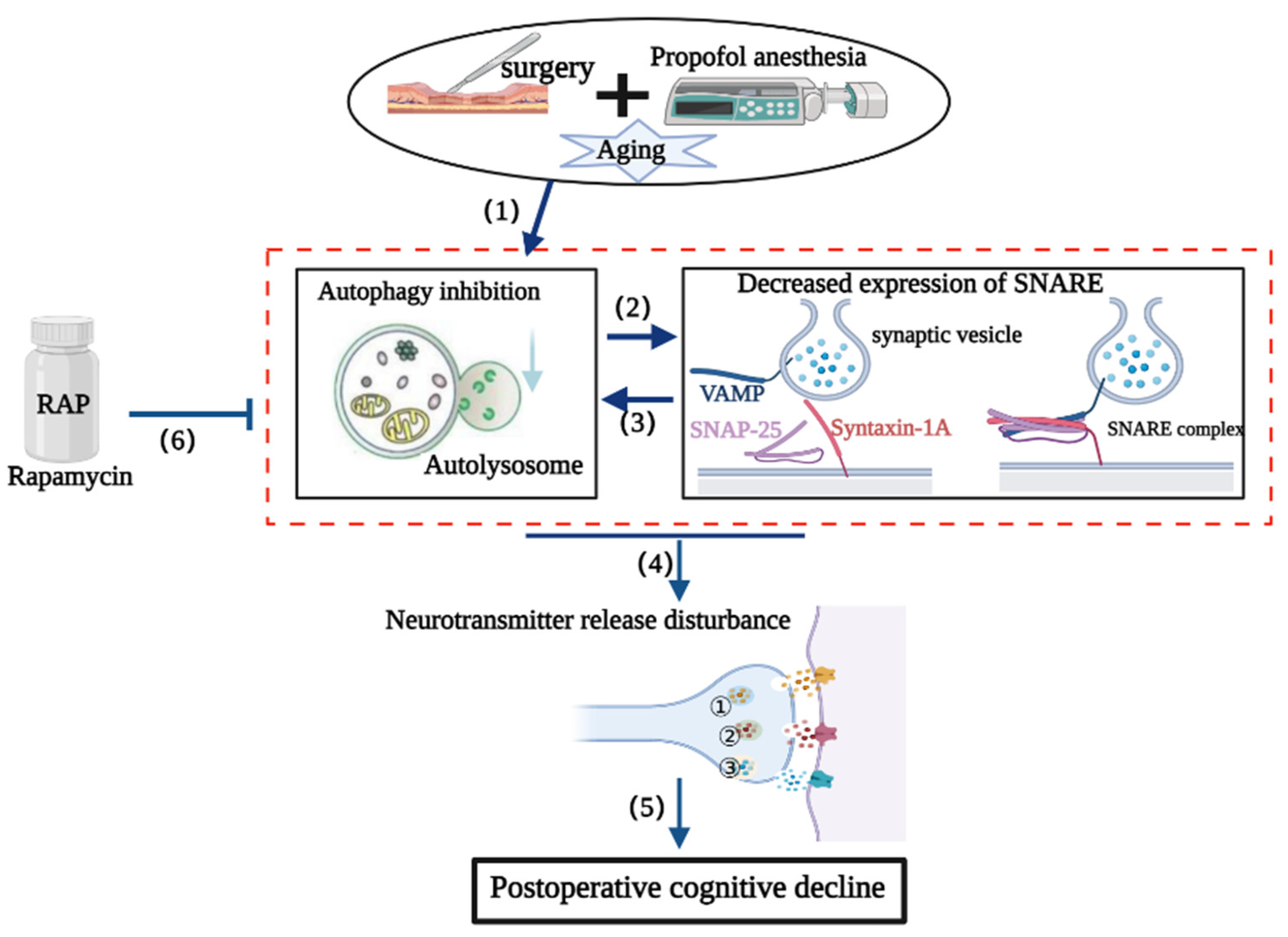
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Evered, L.; Silbert, B.; Knopman, D.S.; Scott, D.A.; DeKosky, S.T.; Rasmussen, L.S.; Oh, E.S.; Crosby, G.; Berger, M.; Eckenhoff, R.G.; et al. Recommendations for the nomenclature of cognitive change associated with anaesthesia and surgery-2018. Br. J. Anaesth. 2018, 121, 1005–1012. [Google Scholar] [CrossRef] [Green Version]
- Holmgaard, F.; Vedel, A.G.; Rasmussen, L.S.; Paulson, O.B.; Nilsson, J.C.; Ravn, H.B. The association between postoperative cognitive dysfunction and cerebral oximetry during cardiac surgery: A secondary analysis of a randomised trial. Br. J. Anaesth. 2019, 123, 196–205. [Google Scholar] [CrossRef]
- Evered, L.A.; Silbert, B.S. Postoperative Cognitive Dysfunction and Noncardiac Surgery. Anesth. Analg. 2018, 127, 496–505. [Google Scholar] [CrossRef]
- Vutskits, L.; Xie, Z. Lasting impact of general anaesthesia on the brain: Mechanisms and relevance. Nat. Rev. Neurosci. 2016, 17, 705–717. [Google Scholar] [CrossRef]
- Needham, M.J.; Webb, C.E.; Bryden, D.C. Postoperative cognitive dysfunction and dementia: What we need to know and do. Br. J. Anaesth. 2017, 119, i115–i125. [Google Scholar] [CrossRef] [Green Version]
- Kahl, U.; Callsen, S.; Beck, S.; Pinnschmidt, H.; von Breunig, F.; Haese, A.; Graefen, M.; Zöllner, C.; Fischer, M. Health-related quality of life and self-reported cognitive function in patients with delayed neurocognitive recovery after radical prostatectomy: A prospective follow-up study. Health Qual. Life Outcomes 2021, 19, 64. [Google Scholar] [CrossRef]
- Yang, N.; Li, Z.; Han, D.; Mi, X.; Tian, M.; Liu, T.; Li, Y.; He, J.; Kuang, C.; Cao, Y.; et al. Autophagy prevents hippocampal alpha-synuclein oligomerization and early cognitive dysfunction after anesthesia/surgery in aged rats. Aging 2020, 12, 7262–7281. [Google Scholar] [CrossRef]
- Sudhof, T.C. Neurotransmitter release: The last millisecond in the life of a synaptic vesicle. Neuron 2013, 80, 675–690. [Google Scholar] [CrossRef] [Green Version]
- Yoon, T.Y.; Munson, M. SNARE complex assembly and disassembly. Curr. Biol. 2018, 28, R397–R401. [Google Scholar] [CrossRef] [Green Version]
- Sauvola, C.W.; Littleton, J.T. SNARE Regulatory Proteins in Synaptic Vesicle Fusion and Recycling. Front. Mol. Neurosci. 2021, 14, 733138. [Google Scholar] [CrossRef]
- Beeri, M.S.; Haroutunian, V.; Schmeidler, J.; Sano, M.; Fam, P.; Kavanaugh, A.; Barr, A.M.; Honer, W.G.; Katsel, P. Synaptic protein deficits are associated with dementia irrespective of extreme old age. Neurobiol. Aging 2012, 33, 1125.e1–1125.e8. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, L.; Hou, C.; Lai, Y.; Long, J.; Liu, J.; Zhong, Q.; Diao, J. SNARE-mediated membrane fusion in autophagy. Semin Cell Dev. Biol. 2016, 60, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Panigrahi, D.P.; Patil, S.; Bhutia, S.K. Autophagy in health and disease: A comprehensive review. Biomed. Pharmacother. 2018, 104, 485–495. [Google Scholar] [CrossRef]
- Mizushima, N.; Levine, B. Autophagy in Human Diseases. N. Engl. J. Med. 2020, 383, 1564–1576. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Islam, W.; Zhang, J.; Zheng, W.; Lu, G.D. Diverse Role of SNARE Protein Sec22 in Vesicle Trafficking, Membrane Fusion, and Autophagy. Cells 2019, 8, 337. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cheng, X.; Li, M.; Wang, Y.; Fu, T.; Zhou, Z.; Gong, X.; Xu, X.; Liu, J.; Pan, L. Decoding three distinct states of the Syntaxin17 SNARE motif in mediating autophagosome-lysosome fusion. Proc. Natl. Acad. Sci. USA 2020, 117, 21391–21402. [Google Scholar] [CrossRef]
- Baker, R.W.; Hughson, F.M. Chaperoning SNARE assembly and disassembly. Nat. Rev. Mol. Cell Biol. 2016, 17, 465–479. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Li, L.; Li, Z.; Ni, C.; Cao, Y.; Liu, T.; Tian, M.; Chui, D.; Guo, X. Protective effect of dapsone on cognitive impairment induced by propofol involves hippocampal autophagy. Neurosci. Lett. 2017, 649, 85–92. [Google Scholar] [CrossRef]
- Li, Y.; Chen, L.; Li, Z.; Song, Y.; Yuan, Y.; Liu, T.; Hong, J.; Wang, Q.; Chang, H.; Kuang, Z.; et al. Potential Serum Biomarkers for Postoperative Neurocognitive Disorders Based on Proteomic Analysis of Cognitive-Related Brain Regions. Front. Aging Neurosci. 2021, 13, 741263. [Google Scholar] [CrossRef]
- Cao, Y.J.; Wang, Q.; Zheng, X.X.; Cheng, Y.; Zhang, Y. Involvement of SNARE complex in the hippocampus and prefrontal cortex of offspring with depression induced by prenatal stress. J. Affect. Disord. 2018, 235, 374–383. [Google Scholar] [CrossRef]
- Li, Y.; Yuan, Y.; Li, Y.; Han, D.; Liu, T.; Yang, N.; Mi, X.; Hong, J.; Liu, K.; Song, Y.; et al. Inhibition of alpha-Synuclein Accumulation Improves Neuronal Apoptosis and Delayed Postoperative Cognitive Recovery in Aged Mice. Oxid. Med. Cell. Longev. 2021, 2021, 5572899. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Liu, T.; Li, Y.; Mi, X.; Han, D.; Yang, N.; Chen, L.; Li, Y.; Hong, J.; Kuang, C.; et al. JNK inhibition alleviates delayed neurocognitive recovery after surgery by limiting microglia pyroptosis. Int. Immunopharmacol. 2021, 99, 107962. [Google Scholar] [CrossRef]
- Mi, X.; Cao, Y.; Li, Y.; Li, Y.; Hong, J.; He, J.; Liang, Y.; Yang, N.; Liu, T.; Han, D.; et al. The Non-peptide Angiotensin-(1-7) Mimic AVE 0991 Attenuates Delayed Neurocognitive Recovery After Laparotomy by Reducing Neuroinflammation and Restoring Blood-Brain Barrier Integrity in Aged Rats. Front. Aging Neurosci. 2021, 13, 624387. [Google Scholar] [CrossRef]
- Burre, J.; Sharma, M.; Tsetsenis, T.; Buchman, V.; Etherton, M.R.; Sudhof, T.C. Alpha-synuclein promotes SNARE-complex assembly in vivo and in vitro. Science 2010, 329, 1663–1667. [Google Scholar] [CrossRef] [Green Version]
- Carlson, S.W.; Yan, H.; Ma, M.; Li, Y.; Henchir, J.; Dixon, C.E. Traumatic Brain Injury Impairs Soluble N-Ethylmaleimide-Sensitive Factor Attachment Protein Receptor Complex Formation and Alters Synaptic Vesicle Distribution in the Hippocampus. J. Neurotrauma 2016, 33, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yang, N.; Li, Y.; Li, Y.; Hong, J.; Wang, Q.; Liu, K.; Han, D.; Han, Y.; Mi, X.; et al. Cholecystokinin octapeptide improves hippocampal glutamatergic synaptogenesis and postoperative cognition by inhibiting induction of A1 reactive astrocytes in aged mice. CNS Neurosci. Ther. 2021, 27, 1374–1384. [Google Scholar] [CrossRef]
- Lian, F.; Cao, C.; Deng, F.; Liu, C.; Zhou, Z. Propofol alleviates postoperative cognitive dysfunction by inhibiting inflammation via up-regulating miR-223-3p in aged rats. Cytokine 2022, 150, 155783. [Google Scholar] [CrossRef]
- Mardini, F.; Tang, J.X.; Li, J.C.; Arroliga, M.J.; Eckenhoff, R.G.; Eckenhoff, M.F. Effects of propofol and surgery on neuropathology and cognition in the 3xTgAD Alzheimer transgenic mouse model. Br. J. Anaesth. 2017, 119, 472–480. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.H.; Culley, D.J.; Baxter, M.G.; Xie, Z.; Tanzi, R.E.; Crosby, G. Spatial memory is intact in aged rats after propofol anesthesia. Anesth. Analg. 2008, 107, 1211–1215. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Tan, H.; Jiang, W.; Zuo, Z. The choice of general anesthetics may not affect neuroinflammation and impairment of learning and memory after surgery in elderly rats. J. Neuroimmune Pharmacol. 2015, 10, 179–189. [Google Scholar] [CrossRef]
- Xu, Z.L.; Chen, G.; Liu, X.; Xie, D.; Zhang, J.; Ying, Y. Effects of ginsenosides on memory impairment in propofol-anesthetized rats. Bioengineered 2022, 13, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Chen, H.; Chen, Y.; Wei, W.; Sun, Y.; Zhang, L.; Cui, L.; Wang, Y. Dysfunction of the SNARE complex in neurological and psychiatric disorders. Pharmacol. Res. 2021, 165, 105469. [Google Scholar] [CrossRef] [PubMed]
- Melland, H.; Carr, E.M.; Gordon, S.L. Disorders of synaptic vesicle fusion machinery. J. Neurochem. 2021, 157, 130–164. [Google Scholar] [CrossRef] [PubMed]
- Margiotta, A. Role of SNAREs in Neurodegenerative Diseases. Cells 2021, 10, 991. [Google Scholar] [CrossRef]
- Pham, E.; Crews, L.; Ubhi, K.; Hansen, L.; Adame, A.; Cartier, A.; Salmon, D.; Galasko, D.; Michael, S.; Savas, J.N.; et al. Progressive accumulation of amyloid-beta oligomers in Alzheimer’s disease and in amyloid precursor protein transgenic mice is accompanied by selective alterations in synaptic scaffold proteins. FEBS J. 2010, 277, 3051–3067. [Google Scholar] [CrossRef] [Green Version]
- Deak, F.; Shin, O.H.; Kavalali, E.T.; Sudhof, T.C. Structural determinants of synaptobrevin 2 function in synaptic vesicle fusion. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 6668–6676. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wang, D.; Sun, T.; Xu, J.; Chiang, H.C.; Shin, W.; Wu, L.G. The SNARE proteins SNAP25 and synaptobrevin are involved in endocytosis at hippocampal synapses. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 9169–9175. [Google Scholar] [CrossRef] [Green Version]
- Haberman, A.; Williamson, W.R.; Epstein, D.; Wang, D.; Rina, S.; Meinertzhagen, I.A.; Hiesinger, P.R. The synaptic vesicle SNARE neuronal Synaptobrevin promotes endolysosomal degradation and prevents neurodegeneration. J. Cell Biol. 2012, 196, 261–276. [Google Scholar] [CrossRef] [Green Version]
- Cornille, F.; Deloye, F.; Fournie-Zaluski, M.C.; Roques, B.P.; Poulain, B. Inhibition of neurotransmitter release by synthetic proline-rich peptides shows that the N-terminal domain of vesicle-associated membrane protein/synaptobrevin is critical for neuro-exocytosis. J. Biol. Chem. 1995, 270, 16826–16832. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Kim, J.; Kim, H.Y.; Ryoo, N.; Lee, S.; Kim, Y.; Rhim, H.; Shin, Y.K. Amyloid-beta Oligomers May Impair SNARE-Mediated Exocytosis by Direct Binding to Syntaxin 1a. Cell Rep. 2015, 12, 1244–1251. [Google Scholar] [CrossRef] [Green Version]
- Kapila, A.K.; Watts, H.R.; Wang, T.; Ma, D. The impact of surgery and anesthesia on post-operative cognitive decline and Alzheimer’s disease development: Biomarkers and preventive strategies. J. Alzheimer’s Dis. 2014, 41, 1–13. [Google Scholar] [CrossRef]
- Dulubova, I.; Khvotchev, M.; Liu, S.; Huryeva, I.; Sudhof, T.C.; Rizo, J. Munc18-1 binds directly to the neuronal SNARE complex. Proc. Natl. Acad. Sci. USA 2007, 104, 2697–2702. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Burre, J.; Sudhof, T.C. CSPalpha promotes SNARE-complex assembly by chaperoning SNAP-25 during synaptic activity. Nat. Cell Biol. 2011, 13, 30–39. [Google Scholar] [CrossRef]
- Boutouja, F.; Stiehm, C.M.; Platta, H.W. mTOR: A Cellular Regulator Interface in Health and Disease. Cells 2019, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Kim, S.G.; Blenis, J. Rapamycin: One drug, many effects. Cell Metab. 2014, 19, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Laguna, M.P. Re: Everolimus for angiomyolipoma associated with tuberous sclerosis complex or sporadic lymphangioleiomyomatosis (EXIST-2): A multicentre, randomised, double-blind, placebo-controlled trial. J. Urol. 2014, 191, 626–627. [Google Scholar] [CrossRef]
- Parlakpinar, H.; Gunata, M. Transplantation and immunosuppression: A review of novel transplant-related immunosuppressant drugs. Immunopharmacol. Immunotoxicol. 2021, 43, 651–665. [Google Scholar] [CrossRef]
- Moreau, K.; Renna, M.; Rubinsztein, D.C. Connections between SNAREs and autophagy. Trends Biochem. Sci. 2013, 38, 57–63. [Google Scholar] [CrossRef]
- Hua, T.; Chen, B.; Lu, D.; Sakurai, K.; Zhao, S.; Han, B.X.; Kim, J.; Yin, L.; Chen, Y.; Lu, J.; et al. General anesthetics activate a potent central pain-suppression circuit in the amygdala. Nat. Neurosci. 2020, 23, 854–868. [Google Scholar] [CrossRef]
- Fechner, J.; Ihmsen, H.; Hatterscheid, D.; Schiessl, C.; Vornov, J.J.; Burak, E.; Schwilden, H.; Schuttler, J. Pharmacokinetics and clinical pharmacodynamics of the new propofol prodrug GPI 15715 in volunteers. Anesthesiology 2003, 99, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storm, H.; Stoen, R.; Klepstad, P.; Skorpen, F.; Qvigstad, E.; Raeder, J. Nociceptive stimuli responses at different levels of general anaesthesia and genetic variability. Acta Anaesthesiol. Scand. 2013, 57, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, G.; Anderson, E.M.; Bokrand-Donatelli, Y.; Neubert, J.K.; Caudle, R.M. Anti-nociceptive effect of a conjugate of substance P and light chain of botulinum neurotoxin type A. Pain 2013, 154, 2547–2553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Xu, W.; Kong, Y.; Huang, J.; Ding, Z.; Deng, M.; Guo, Q.; Zou, W. SNAP-25 Contributes to Neuropathic Pain by Regulation of VGLuT2 Expression in Rats. Neuroscience 2019, 423, 86–97. [Google Scholar] [CrossRef]
- Zhu, X.; Yue, L.; Fan, C.; Liu, Y.; Wang, Y.; Zhao, H. Mechanism of Cdk5-synaptophysin-SNARE pathway in acute and chronic inflammatory pain. Am. J. Transl. Res. 2021, 13, 1075–1084. [Google Scholar] [PubMed]
- Frank, M.; Nabb, A.T.; Gilbert, S.P.; Bentley, M. Propofol attenuates kinesin-mediated axonal vesicle transport and fusion. Mol. Biol. Cell 2022, 33, ar119. [Google Scholar] [CrossRef]
- Bademosi, A.T.; Steeves, J.; Karunanithi, S.; Zalucki, O.H.; Gormal, R.S.; Liu, S.; Lauwers, E.; Verstreken, P.; Anggono, V.; Meunier, F.A.; et al. Trapping of Syntaxin1a in Presynaptic Nanoclusters by a Clinically Relevant General Anesthetic. Cell Rep. 2018, 22, 427–440. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; McMillan, K.; Pike, C.M.; Cahill, A.L.; Herring, B.E.; Wang, Q.; Fox, A.P. Interaction of anesthetics with neurotransmitter release machinery proteins. J. Neurophysiol. 2013, 109, 758–767. [Google Scholar] [CrossRef] [Green Version]
- Herring, B.E.; McMillan, K.; Pike, C.M.; Marks, J.; Fox, A.P.; Xie, Z. Etomidate and propofol inhibit the neurotransmitter release machinery at different sites. J. Physiol. 2011, 589, 1103–1115. [Google Scholar] [CrossRef]
- Agirman, G.; Yu, K.B.; Hsiao, E.Y. Signaling inflammation across the gut-brain axis. Science 2021, 374, 1087–1092. [Google Scholar] [CrossRef]
- Han, D.; Li, Z.; Liu, T.; Yang, N.; Li, Y.; He, J.; Qian, M.; Kuang, Z.; Zhang, W.; Ni, C.; et al. Prebiotics Regulation of Intestinal Microbiota Attenuates Cognitive Dysfunction Induced by Surgery Stimulation in APP/PS1 Mice. Aging Dis. 2020, 11, 1029–1045. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, N.; Han, X.; Li, Z.; Liu, T.; Mi, X.; Li, Y.; Guo, X.; Han, D.; Yang, N. Rapamycin Affects the Hippocampal SNARE Complex to Alleviate Cognitive Dysfunction Induced by Surgery in Aged Rats. Brain Sci. 2023, 13, 598. https://doi.org/10.3390/brainsci13040598
Kang N, Han X, Li Z, Liu T, Mi X, Li Y, Guo X, Han D, Yang N. Rapamycin Affects the Hippocampal SNARE Complex to Alleviate Cognitive Dysfunction Induced by Surgery in Aged Rats. Brain Sciences. 2023; 13(4):598. https://doi.org/10.3390/brainsci13040598
Chicago/Turabian StyleKang, Ning, Xiaoguang Han, Zhengqian Li, Taotao Liu, Xinning Mi, Yue Li, Xiangyang Guo, Dengyang Han, and Ning Yang. 2023. "Rapamycin Affects the Hippocampal SNARE Complex to Alleviate Cognitive Dysfunction Induced by Surgery in Aged Rats" Brain Sciences 13, no. 4: 598. https://doi.org/10.3390/brainsci13040598