

Cannabinoid Receptor Agonist WIN55, 212-2 Attenuates Injury in the Hippocampus of Rats after Deep Hypothermic Circulatory Arrest

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Drug Administration
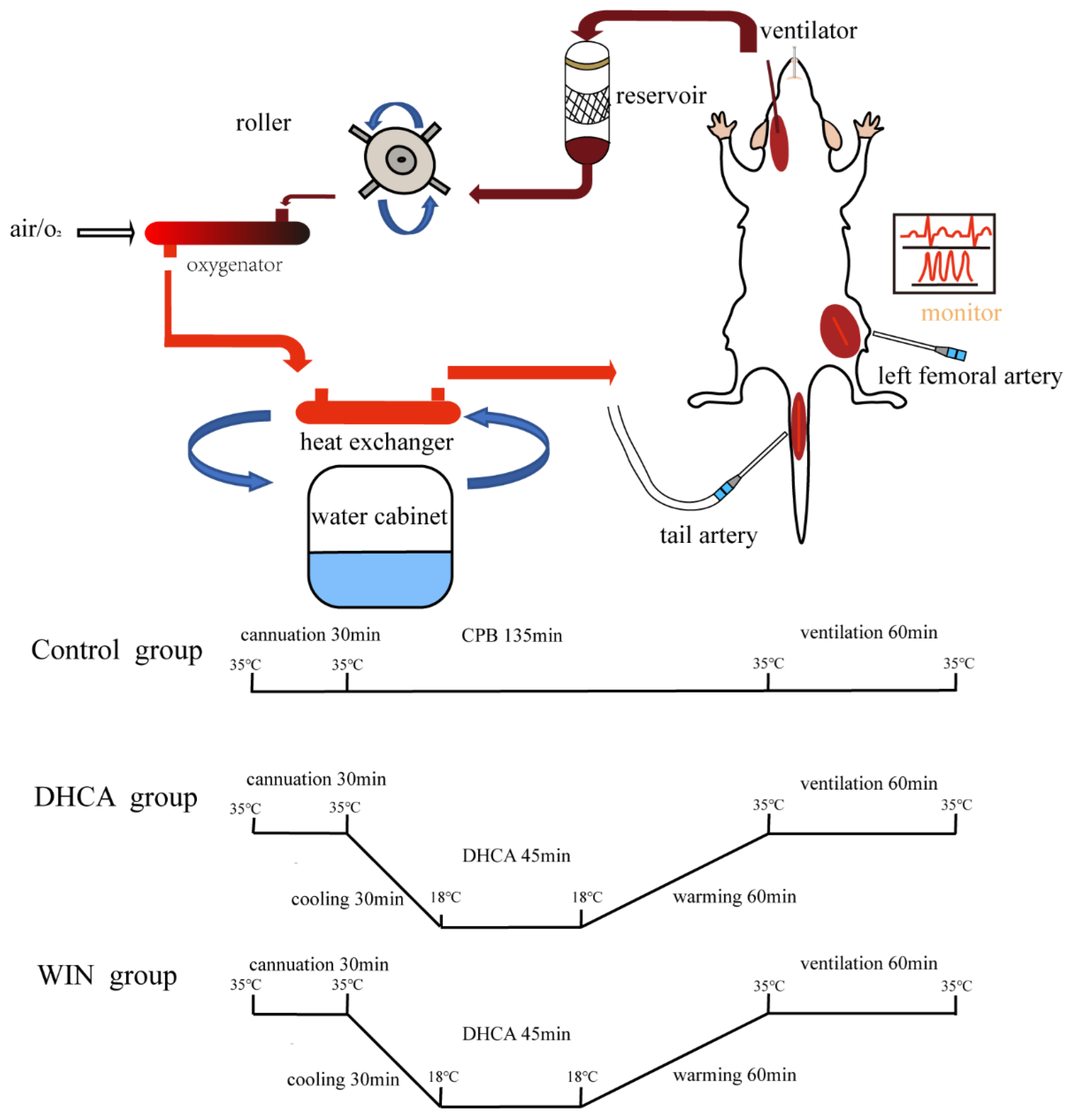
2.2. CPB and DHCA Procedures
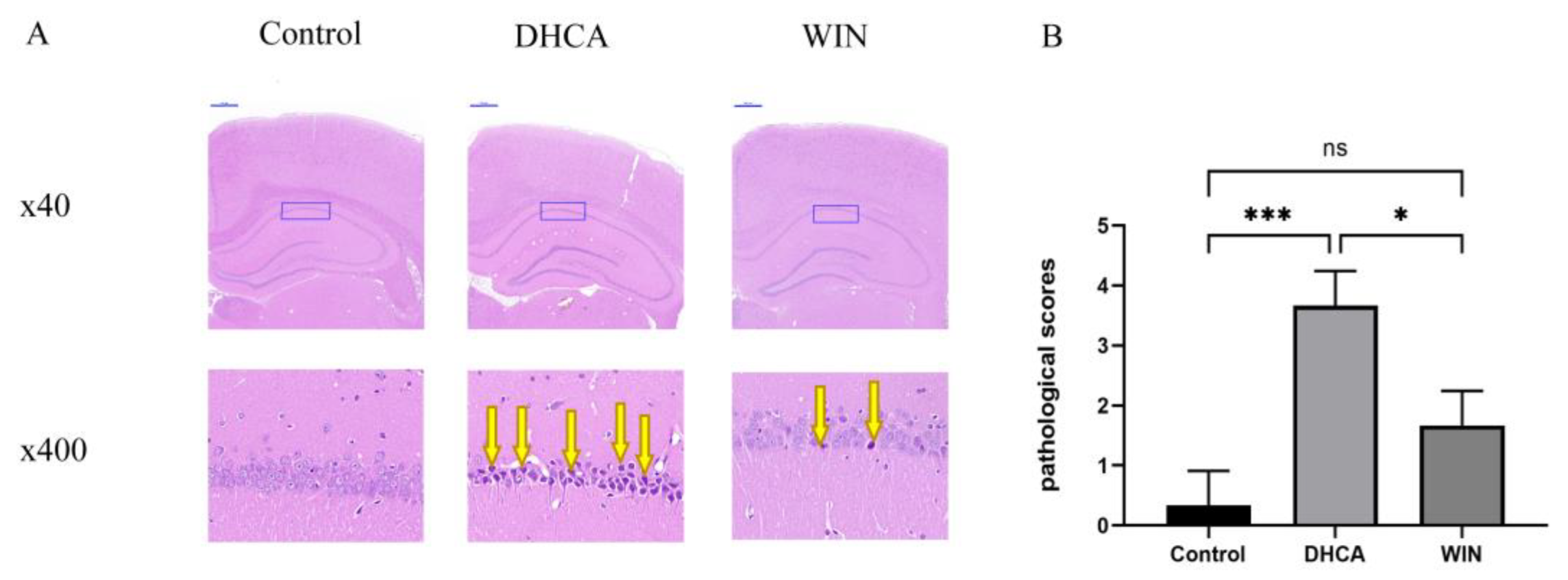
2.3. Histopathological Study
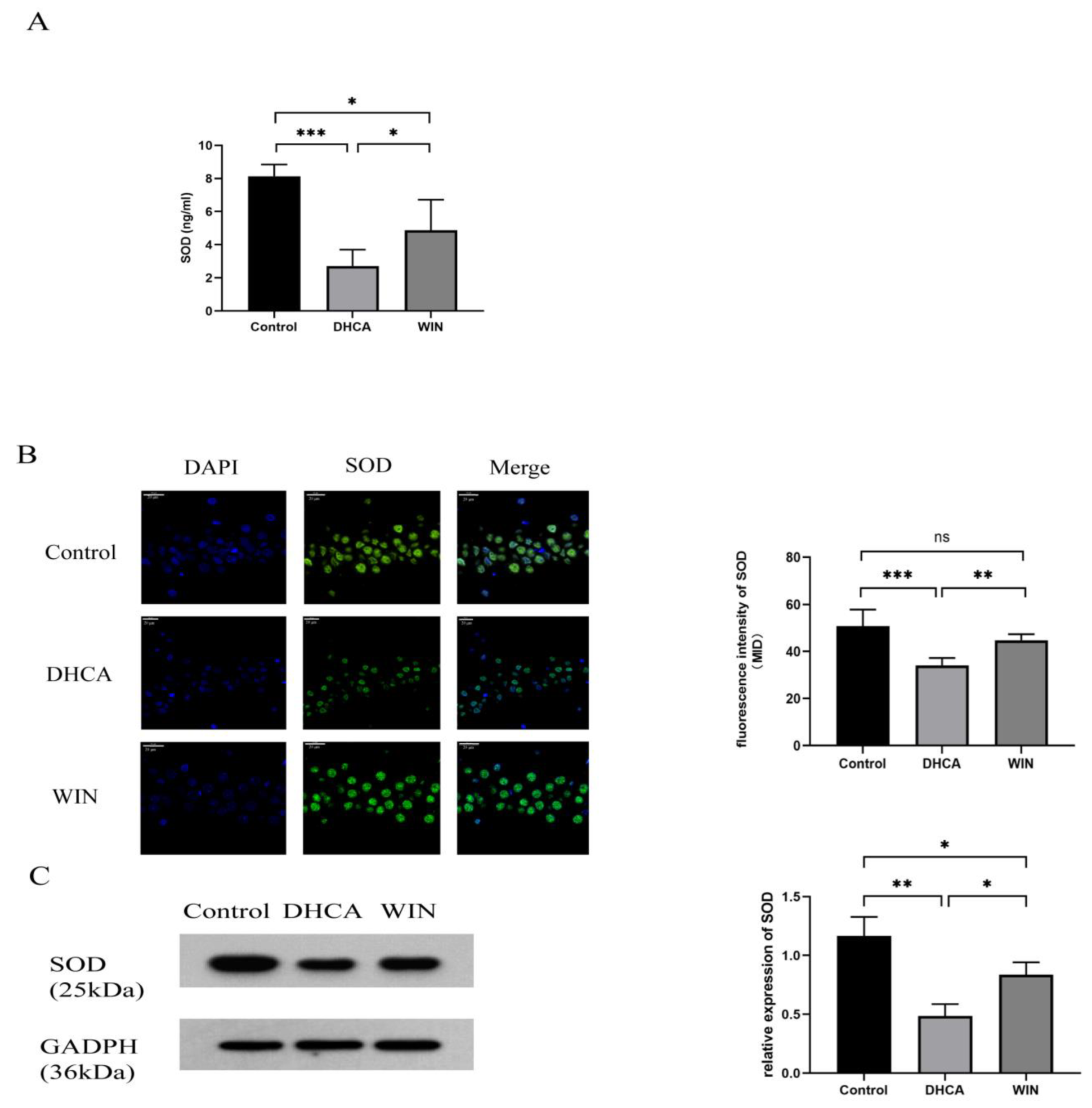
2.4. Immunofluorescence Staining of SOD
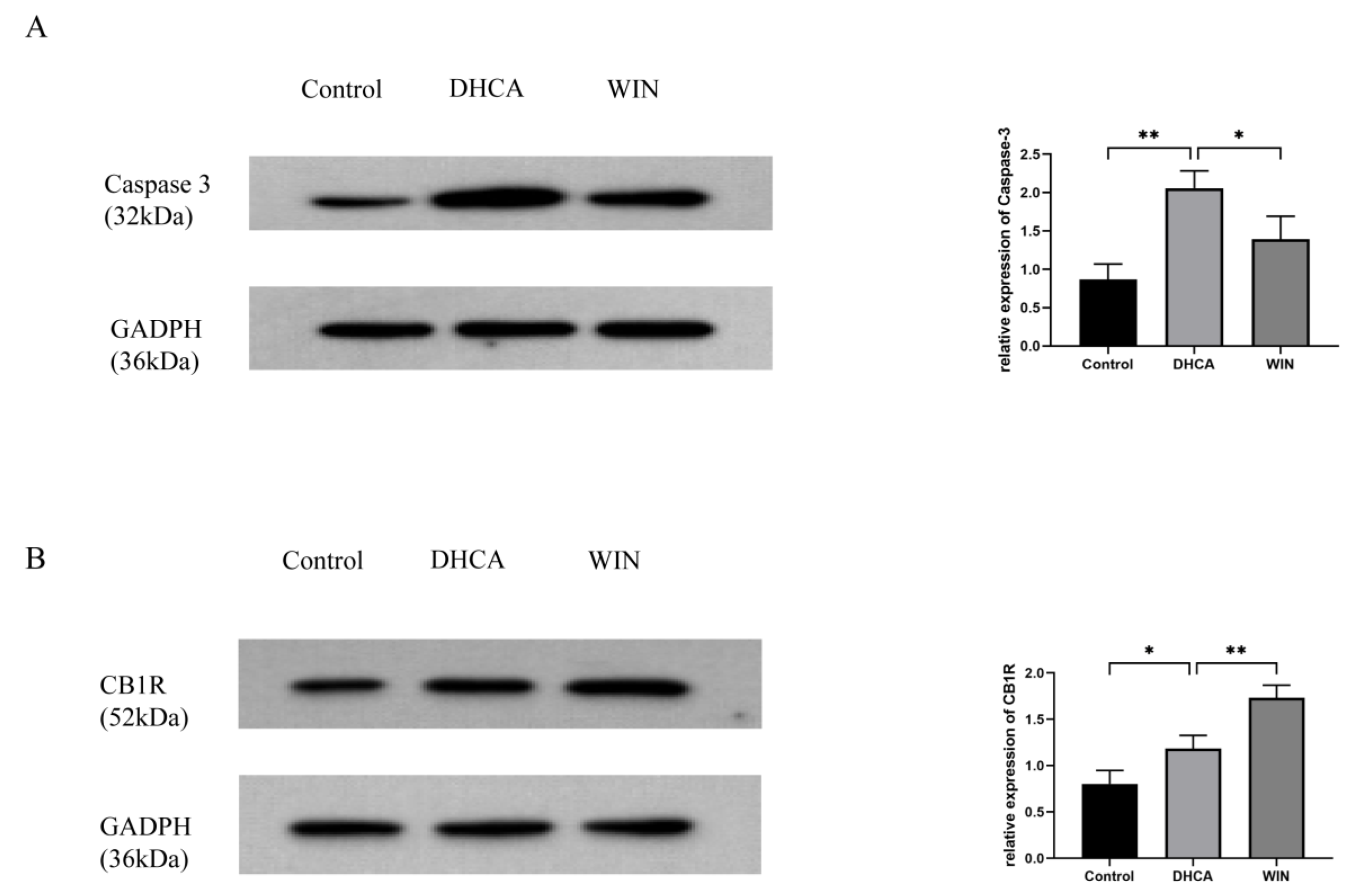
2.5. Western Blot Analysis
2.6. Enzyme-Linked Immunosorbent Assay
2.7. Statistical Analysis
3. Results
3.1. WIN55, 212-2 Alleviates Hippocampal Injury in Rats Underwent DHCA
3.2. WIN55, 212-2 Attenuates the Production of Proinflammatory Cytokines in Rats That Underwent DHCA
3.3. WIN55, 212-2 Restores SOD Level in Rats That Underwent DHCA
3.4. WIN55, 212-2 Inhibits the Expression of Apoptotic Protein and Upregulates the Expression of CB1R in the Hippocampal Tissue of Rats That Underwent DHCA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DHCA | deep hypothermic circulatory arrest |
| CB1R | type 1 cannabinoid receptor |
| CB2R | type 2 cannabinoid receptor |
| CPB | cardiopulmonary bypass |
| IL | interleukin |
| TNF-α | tumor necrosis factor-alpha |
| SOD | superoxide dismutase |
| ROS | reactive oxygen species |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| ELISA | enzyme-linked immunosorbent assay |
| MID | mean immunofluorescence density |
| CNS | central nervous system |
| AEA | anandamide |
| 2-AG | 2-arachidonoylglycerol |
| NF-κB | nuclear factor kappa-B |
| AC | adenylyl cyclase |
| MAPKs | mitogen-activated protein kinases |
| PKA | protein kinase A |
| ERK | Extracellular-regulated protein kinases |
| JNK | Jun N-terminal kinase |
References
- Stinear, C.M.; Lang, C.E.; Zeiler, S.; Byblow, W.D. Advances and challenges in stroke rehabilitation. Lancet Neurol. 2020, 19, 348–360. [Google Scholar] [CrossRef]
- Algra, S.O.; Jansen, N.J.; van der Tweel, I.; Schouten, A.N.; Groenendaal, F.; Toet, M.; van Oeveren, W.; van Haastert, I.C.; Schoof, P.H.; de Vries, L.S.; et al. Neurological injury after neonatal cardiac surgery: A randomized, controlled trial of 2 perfusion techniques. Circulation 2014, 129, 224–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziganshin, B.A.; Elefteriades, J.A. Deep hypothermic circulatory arrest. Ann. Cardiothorac. Surg. 2013, 2, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Chau, K.H.; Friedman, T.; Tranquilli, M.; Elefteriades, J.A. Deep hypothermic circulatory arrest effectively preserves neurocognitive function. Ann. Thorac. Surg. 2013, 96, 1553–1559. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Li, H.; Wang, D.; Wu, C.; Pan, Z.; Li, Y.; An, Y.; Wang, G.; Dai, J.; Wang, Q. Effects of four major brain protection strategies during proximal aortic surgery: A systematic review and network meta-analysis. Int. J. Surg. 2019, 63, 8–15. [Google Scholar] [CrossRef]
- Kellermann, K.; Gordan, M.L.; Nollert, G.; Blobner, M.; Kochs, E.F.; Jungwirth, B. Long-term assessment of NFkappaB expression in the brain and neurologic outcome following deep hypothermic circulatory arrest in rats. Perfusion 2009, 24, 429–436. [Google Scholar] [CrossRef]
- Chen, Q.; Lei, Y.Q.; Liu, J.F.; Wang, Z.C.; Cao, H. Triptolide improves neurobehavioral functions, inflammation, and oxidative stress in rats under deep hypothermic circulatory arrest. Aging 2021, 13, 3031–3044. [Google Scholar] [CrossRef]
- Zhang, Z.; Ma, Q.; Shah, B.; Mackensen, G.B.; Lo, D.C.; Mathew, J.P.; Podgoreanu, M.V.; Terrando, N. Neuroprotective Effects of Annexin A1 Tripeptide after Deep Hypothermic Circulatory Arrest in Rats. Front. Immunol. 2017, 8, 1050. [Google Scholar] [CrossRef] [Green Version]
- Kajimoto, M.; Nuri, M.; Sleasman, J.R.; Charette, K.A.; Nelson, B.R.; Portman, M.A. Inhaled nitric oxide reduces injury and microglia activation in porcine hippocampus after deep hypothermic circulatory arrest. J. Thorac. Cardiovasc. Surg. 2021, 161, e485–e498. [Google Scholar] [CrossRef]
- Shen, L.; Wang, J.; Liu, K.; Wang, C.; Wang, C.; Wu, H.; Sun, Q.; Sun, X.; Jing, H. Hydrogen-rich saline is cerebroprotective in a rat model of deep hypothermic circulatory arrest. Neurochem. Res. 2011, 36, 1501–1511. [Google Scholar] [CrossRef]
- Germano, A.F.; Dixon, C.E.; d’Avella, D.; Hayes, R.L.; Tomasello, F. Behavioral deficits following experimental subarachnoid hemorrhage in the rat. J. Neurotrauma 1994, 11, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R.; et al. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 1995, 50, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.D.; Davison, J.S.; Pittman, Q.J.; Sharkey, K.A. Cannabinoid CB(2) receptors in health and disease. Curr. Med. Chem. 2010, 17, 1393–1410. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.C.; Mackie, K. An Introduction to the Endogenous Cannabinoid System. Biol. Psychiatry 2016, 79, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Munro, S.; Thomas, K.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef]
- Herkenham, M.; Lynn, A.B.; Little, M.D.; Johnson, M.R.; Melvin, L.S.; de Costa, B.R.; Rice, K.C. Cannabinoid receptor localization in brain. Proc. Natl. Acad. Sci. USA 1990, 87, 1932–1936. [Google Scholar] [CrossRef] [Green Version]
- Tsou, K.; Brown, S.; Sanudo-Pena, M.C.; Mackie, K.; Walker, J.M. Immunohistochemical distribution of cannabinoid CB1 receptors in the rat central nervous system. Neuroscience 1998, 83, 393–411. [Google Scholar] [CrossRef]
- Zou, S.; Kumar, U. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Lopez, D.; Faustino, J.; Derugin, N.; Wendland, M.; Lizasoain, I.; Moro, M.A.; Vexler, Z.S. Reduced infarct size and accumulation of microglia in rats treated with WIN 55,212-2 after neonatal stroke. Neuroscience 2012, 207, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Won, S.J.; Mao, X.O.; Jin, K.; Greenberg, D.A. Involvement of protein kinase A in cannabinoid receptor-mediated protection from oxidative neuronal injury. J. Pharmacol. Exp. Ther. 2005, 313, 88–94. [Google Scholar] [CrossRef]
- Zoppi, S.; Perez Nievas, B.G.; Madrigal, J.L.; Manzanares, J.; Leza, J.C.; Garcia-Bueno, B. Regulatory role of cannabinoid receptor 1 in stress-induced excitotoxicity and neuroinflammation. Neuropsychopharmacology 2011, 36, 805–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Alconada, D.; Alvarez, F.J.; Goni-de-Cerio, F.; Hilario, E.; Alvarez, A. Cannabinoid-mediated Modulation of Oxidative Stress and Early Inflammatory Response after Hypoxia-Ischemia. Int. J. Mol. Sci. 2020, 21, 1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampson, A.J.; Grimaldi, M.; Axelrod, J.; Wink, D. Cannabidiol and (−)Delta9-tetrahydrocannabinol are neuroprotective antioxidants. Proc. Natl. Acad. Sci. USA 1998, 95, 8268–8273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarruk, J.G.; Fernandez-Lopez, D.; Garcia-Yebenes, I.; Garcia-Gutierrez, M.S.; Vivancos, J.; Nombela, F.; Torres, M.; Burguete, M.C.; Manzanares, J.; Lizasoain, I.; et al. Cannabinoid type 2 receptor activation downregulates stroke-induced classic and alternative brain macrophage/microglial activation concomitant to neuroprotection. Stroke 2012, 43, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Bonfils, P.K.; Reith, J.; Hasseldam, H.; Johansen, F.F. Estimation of the hypothermic component in neuroprotection provided by cannabinoids following cerebral ischemia. Neurochem. Int. 2006, 49, 508–518. [Google Scholar] [CrossRef]
- Li, Y.A.; Liu, Z.G.; Zhang, Y.P.; Hou, H.T.; He, G.W.; Xue, L.G.; Yang, Q.; Liu, X.C. Differential expression profiles of circular RNAs in the rat hippocampus after deep hypothermic circulatory arrest. Artif. Organs 2021, 45, 866–880. [Google Scholar] [CrossRef]
- Eichenbaum, H. A cortical-hippocampal system for declarative memory. Nat. Rev. Neurosci. 2000, 1, 41–50. [Google Scholar] [CrossRef]
- Squire, L.R. Memory and the hippocampus: A synthesis from findings with rats, monkeys, and humans. Psychol. Rev. 1992, 99, 195–231. [Google Scholar] [CrossRef]
- Wu, M.V.; Sahay, A.; Duman, R.S.; Hen, R. Functional differentiation of adult-born neurons along the septotemporal axis of the dentate gyrus. Cold Spring Harb. Perspect. Biol. 2015, 7, a018978. [Google Scholar] [CrossRef] [Green Version]
- Lana, D.; Ugolini, F.; Giovannini, M.G. An Overview on the Differential Interplay Among Neurons-Astrocytes-Microglia in CA1 and CA3 Hippocampus in Hypoxia/Ischemia. Front. Cell. Neurosci. 2020, 14, 585833. [Google Scholar] [CrossRef]
- Valentine, J.S.; Doucette, P.A.; Zittin Potter, S. Copper-zinc superoxide dismutase and amyotrophic lateral sclerosis. Annu. Rev. Biochem. 2005, 74, 563–593. [Google Scholar] [CrossRef] [Green Version]
- Tu, Q.; Wang, R.; Ding, B.; Zhong, W.; Cao, H. Protective and antioxidant effect of Danshen polysaccharides on cerebral ischemia/reperfusion injury in rats. Int. J. Biol. Macromol. 2013, 60, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, S.; Zhang, X.; Li, Y.; Zhao, Q.; Liu, T. Pterostilbene protects against myocardial ischemia/reperfusion injury via suppressing oxidative/nitrative stress and inflammatory response. Int. Immunopharmacol. 2017, 43, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Chen, X.; Gao, Y.; Liu, Z.; Zhai, Q.; Xiong, L.; Cai, M.; Wang, Q. Mn-SOD Upregulation by Electroacupuncture Attenuates Ischemic Oxidative Damage via CB1R-Mediated STAT3 Phosphorylation. Mol. Neurobiol. 2016, 53, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Ma, L.; Wu, M.; Zhang, L.; Zhang, X.; Zhai, Q.; Jiang, T.; Wang, Q.; Xiong, L. Anandamide protects HT22 cells exposed to hydrogen peroxide by inhibiting CB1 receptor-mediated type 2 NADPH oxidase. Oxid. Med. Cell. Longev. 2014, 2014, 893516. [Google Scholar] [CrossRef] [Green Version]
- Wiegman, C.H.; Michaeloudes, C.; Haji, G.; Narang, P.; Clarke, C.J.; Russell, K.E.; Bao, W.; Pavlidis, S.; Barnes, P.J.; Kanerva, J.; et al. Oxidative stress-induced mitochondrial dysfunction drives inflammation and airway smooth muscle remodeling in patients with chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2015, 136, 769–780. [Google Scholar] [CrossRef] [Green Version]
- Jungwirth, B.; Eckel, B.; Blobner, M.; Kellermann, K.; Kochs, E.F.; Mackensen, G.B. The impact of cardiopulmonary bypass on systemic interleukin-6 release, cerebral nuclear factor-kappa B expression, and neurocognitive outcome in rats. Anesth. Analg. 2010, 110, 312–320. [Google Scholar] [CrossRef]
- Reinsfelt, B.; Ricksten, S.E.; Zetterberg, H.; Blennow, K.; Freden-Lindqvist, J.; Westerlind, A. Cerebrospinal fluid markers of brain injury, inflammation, and blood-brain barrier dysfunction in cardiac surgery. Ann. Thorac. Surg. 2012, 94, 549–555. [Google Scholar] [CrossRef]
- Chen, L.; Lv, L.; Long, C.; Lou, S. Effects of circuit albumin coating on coagulation and inflammatory response for patients receiving aortic arch replacement: A randomized controlled trial. Perfusion 2016, 31, 576–583. [Google Scholar] [CrossRef]
- Jiang, X.; Gu, T.; Liu, Y.; Gao, S.; Shi, E.; Zhang, G. Chipmunk Brain Is Resistant to Injury from Deep Hypothermic Circulatory Arrest During Cardiopulmonary Bypass. Ther. Hypothermia Temp. Manag. 2019, 9, 118–127. [Google Scholar] [CrossRef]
- Alonso-Alconada, D.; Alvarez, A.; Alvarez, F.J.; Martinez-Orgado, J.A.; Hilario, E. The cannabinoid WIN 55212-2 mitigates apoptosis and mitochondrial dysfunction after hypoxia ischemia. Neurochem. Res. 2012, 37, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Fang, Y.Q.; Ren, H.; Chen, T.; Guo, J.J.; Yan, J.; Song, S.; Zhang, L.Y.; Liao, H. WIN55,212-2 protects oligodendrocyte precursor cells in stroke penumbra following permanent focal cerebral ischemia in rats. Acta Pharmacol. Sin. 2013, 34, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Martin, B.; Adler, M.; Razdan, R.; Ganea, D.; Tuma, R. Modulation of the balance between cannabinoid CB(1) and CB(2) receptor activation during cerebral ischemic/reperfusion injury. Neuroscience 2008, 152, 753–760. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-kappaB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, K.; Karin, M. NF-kappaB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef]
- Li, L.J.; Zhang, S.J.; Liu, P.; Wang, Y.Q.; Chen, Z.L.; Wang, Y.J.; Zhou, J.B.; Guo, Y.J.; Zhao, L. Corilagin Interferes with Toll-Like Receptor 3-Mediated Immune Response in Herpes Simplex Encephalitis. Front. Mol. Neurosci. 2019, 12, 83. [Google Scholar] [CrossRef]
- Bhargava, A.; Shukla, A.; Bunkar, N.; Shandilya, R.; Lodhi, L.; Kumari, R.; Gupta, P.K.; Rahman, A.; Chaudhury, K.; Tiwari, R.; et al. Exposure to ultrafine particulate matter induces NF-kappabeta mediated epigenetic modifications. Environ. Pollut. 2019, 252, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Ma, R.; Chai, X.; Liang, H.J.; Jiang, P.; Zhu, X.L.; Chen, X.; Su, B.X. Inhibiting a spinal cord signaling pathway protects against ischemia injury in rats. J. Thorac. Cardiovasc. Surg. 2019, 157, 494–503.e1. [Google Scholar] [CrossRef]
- Kellermann, K.; Dertinger, N.; Blobner, M.; Kees, F.; Kochs, E.F.; Jungwirth, B. Perioperative moxifloxacin treatment in rats subjected to deep hypothermic circulatory arrest: Reduction in cerebral inflammation but without improvement in cognitive performance. J. Thorac. Cardiovasc. Surg. 2011, 141, 796–802. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, N.; Ruthirakuhan, M.; Gallagher, D.; Verhoeff, N.; Kiss, A.; Black, S.E.; Lanctot, K.L. Randomized Placebo-Controlled Trial of Nabilone for Agitation in Alzheimer’s Disease. Am. J. Geriatr. Psychiatry 2019, 27, 1161–1173. [Google Scholar] [CrossRef]
- Patel, A.D.; Mazurkiewicz-Beldzinska, M.; Chin, R.F.; Gil-Nagel, A.; Gunning, B.; Halford, J.J.; Mitchell, W.; Scott Perry, M.; Thiele, E.A.; Weinstock, A.; et al. Long-term safety and efficacy of add-on cannabidiol in patients with Lennox-Gastaut syndrome: Results of a long-term open-label extension trial. Epilepsia 2021, 62, 2228–2239. [Google Scholar] [CrossRef]
- Alizadeh, S.; Djafarian, K.; Mofidi Nejad, M.; Yekaninejad, M.S.; Javanbakht, M.H. The effect of beta-caryophyllene on food addiction and its related behaviors: A randomized, double-blind, placebo-controlled trial. Appetite 2022, 178, 106160. [Google Scholar] [CrossRef] [PubMed]
- Javid, F.A.; Phillips, R.M.; Afshinjavid, S.; Verde, R.; Ligresti, A. Cannabinoid pharmacology in cancer research: A new hope for cancer patients? Eur. J. Pharmacol. 2016, 775, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Toth, K.F.; Adam, D.; Biro, T.; Olah, A. Cannabinoid Signaling in the Skin: Therapeutic Potential of the “C(ut)annabinoid” System. Molecules 2019, 24, 918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, H.; Toyang, N.; Steele, B.; Bryant, J.; Ngwa, W. The Endocannabinoid System: A Potential Target for the Treatment of Various Diseases. Int. J. Mol. Sci. 2021, 22, 9472. [Google Scholar] [CrossRef] [PubMed]
- Pagano, C.; Navarra, G.; Coppola, L.; Avilia, G.; Bifulco, M.; Laezza, C. Cannabinoids: Therapeutic Use in Clinical Practice. Int. J. Mol. Sci. 2022, 23, 3344. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, M.-H.; Yang, Q.; Zhang, Y.-P.; Wang, J.-H.; Zhang, R.-J.-Z.; Liu, Z.-G.; Liu, X.-C. Cannabinoid Receptor Agonist WIN55, 212-2 Attenuates Injury in the Hippocampus of Rats after Deep Hypothermic Circulatory Arrest. Brain Sci. 2023, 13, 525. https://doi.org/10.3390/brainsci13030525
Yu M-H, Yang Q, Zhang Y-P, Wang J-H, Zhang R-J-Z, Liu Z-G, Liu X-C. Cannabinoid Receptor Agonist WIN55, 212-2 Attenuates Injury in the Hippocampus of Rats after Deep Hypothermic Circulatory Arrest. Brain Sciences. 2023; 13(3):525. https://doi.org/10.3390/brainsci13030525
Chicago/Turabian StyleYu, Ming-Huan, Qin Yang, You-Peng Zhang, Jia-Hui Wang, Ren-Jian-Zhi Zhang, Zhi-Gang Liu, and Xiao-Cheng Liu. 2023. "Cannabinoid Receptor Agonist WIN55, 212-2 Attenuates Injury in the Hippocampus of Rats after Deep Hypothermic Circulatory Arrest" Brain Sciences 13, no. 3: 525. https://doi.org/10.3390/brainsci13030525