
The Effects of Social Processing and Role Type on Attention Networks: Insights from Team Ball Athletes

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
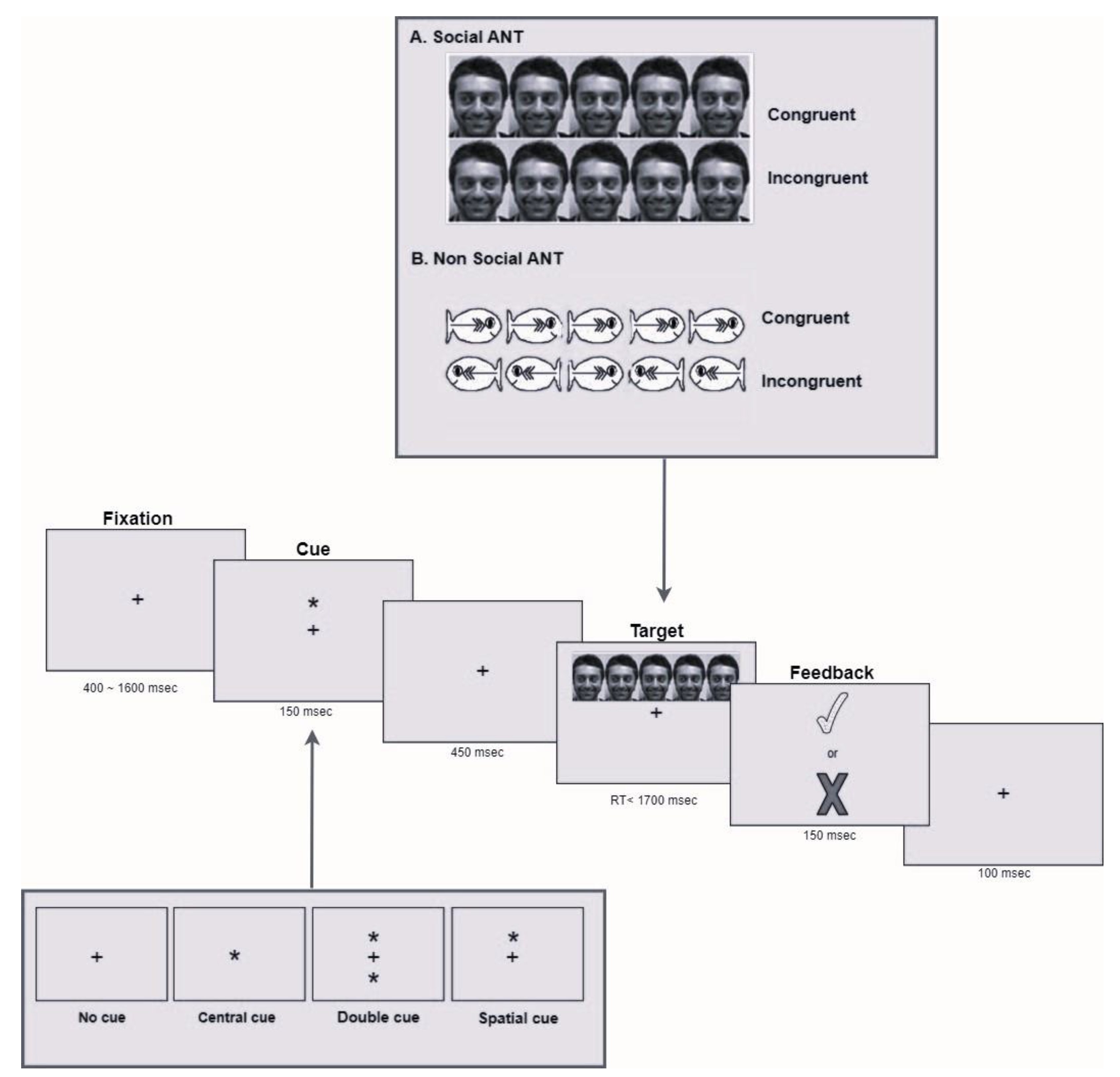
2.2. Attention Network Test (ANT)
2.2.1. Apparatus
2.2.2. Stimuli
2.2.3. Procedure
2.2.4. Data Analysis
2.3. Statistical Analysis
3. Results
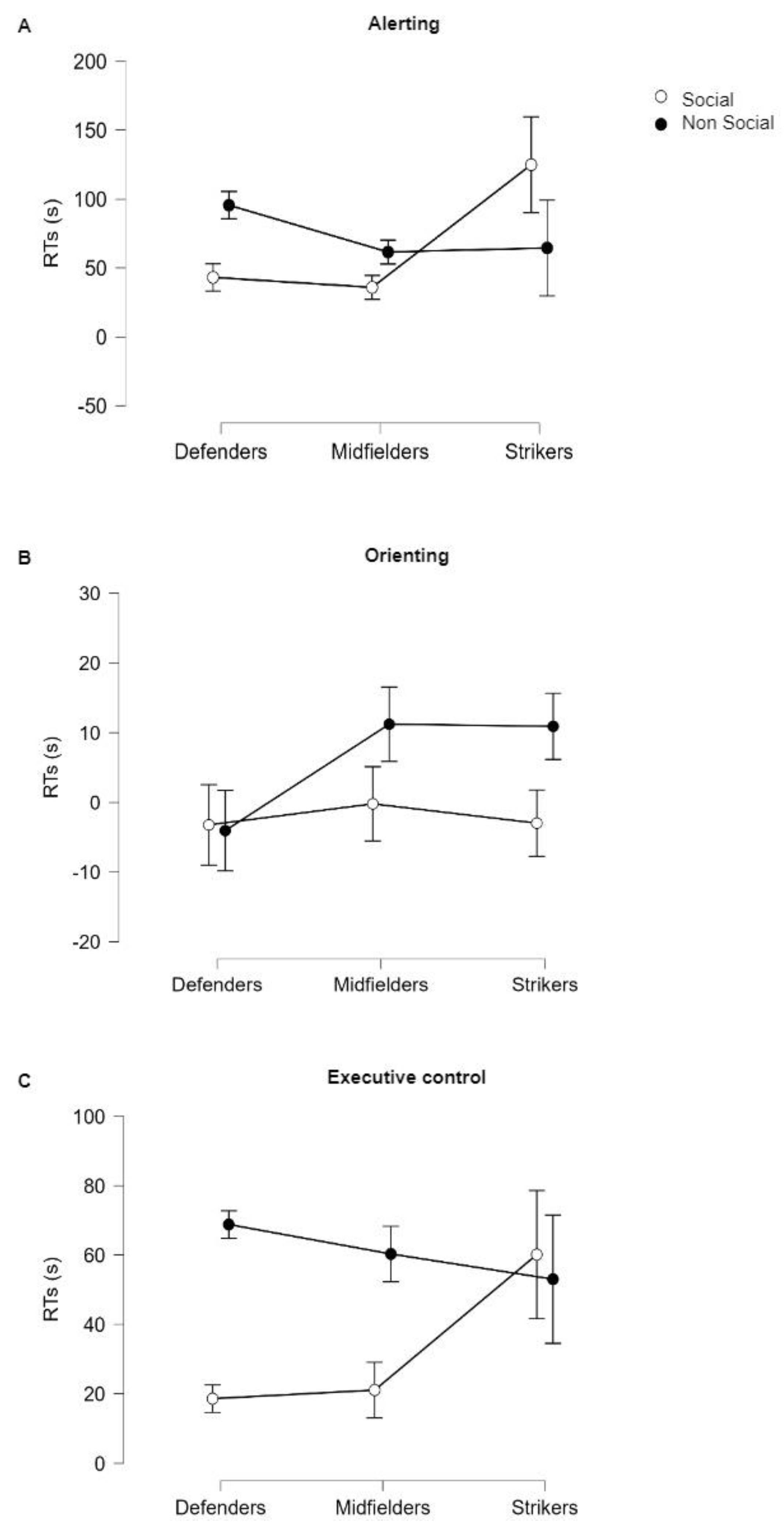
3.1. Stimulus × Role Effects on Reaction Times (RTs)
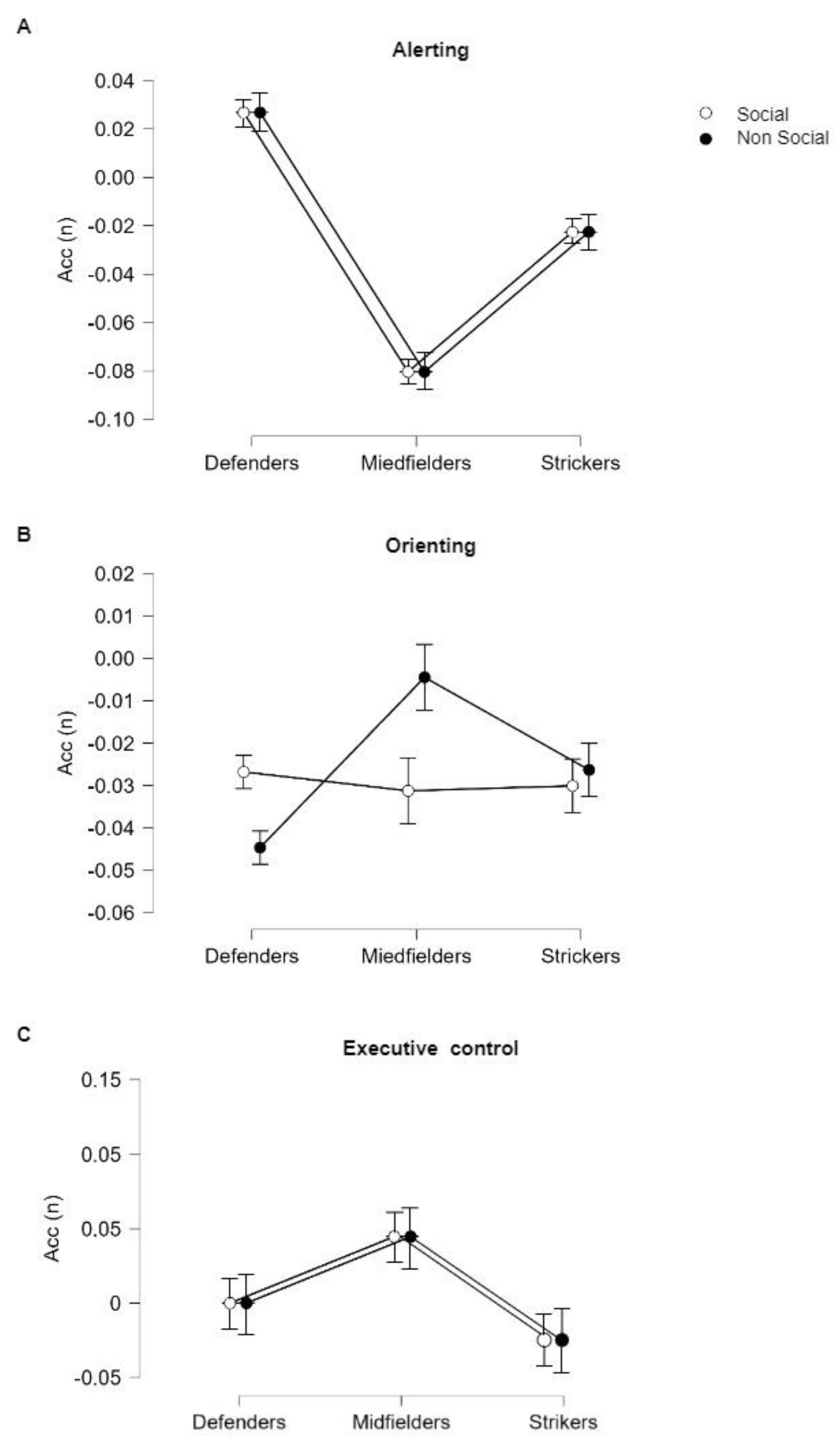
3.2. Stimulus × Role Effects on Accuracy (Acc)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davies, D.R.; Parasuraman, R. Arieties of Attention Eds.; Academic Press Series in Cognition and Perception; Academic Press: Orlando, FL, USA, 1984; ISBN 978-0-12-544970-0. [Google Scholar]
- Posner, M.I.; Petersen, S.E. The Attention System of the Human Brain. Annu. Rev. Neurosci. 1990, 13, 25–42. [Google Scholar] [CrossRef]
- Fougnie, D. The Relationship between Attention and Working Memory. New Res. Short-Term Mem. 2008, 1, 45. [Google Scholar]
- Fan, J.; McCandliss, B.D.; Sommer, T.; Raz, A.; Posner, M.I. Testing the Efficiency and Independence of Attentional Networks. J. Cogn. Neurosci. 2002, 14, 340–347. [Google Scholar] [CrossRef]
- Fan, J.; Gu, X.; Guise, K.G.; Liu, X.; Fossella, J.; Wang, H.; Posner, M.I. Testing the Behavioral Interaction and Integration of Attentional Networks. Brain Cogn. 2009, 70, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, D.T.Y.; Williams, A.M.; Ward, P.; Janelle, C.M. Perceptual-Cognitive Expertise in Sport: A Meta-Analysis. J. Sport Exerc. Psychol. 2007, 29, 457–478. [Google Scholar] [CrossRef]
- Fegatelli, D.; Giancamilli, F.; Mallia, L.; Chirico, A.; Lucidi, F. The Use of Eye Tracking (ET) in Targeting Sports: A Review of the Studies on Quiet Eye (QE). In Intelligent Interactive Multimedia Systems and Services 2016; Pietro, G.D., Gallo, L., Howlett, R.J., Jain, L.C., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 715–730. [Google Scholar]
- Chirico, A.; Fegatelli, D.; Galli, F.; Mallia, L.; Alivernini, F.; Cordone, S.; Giancamilli, F.; Pecci, S.; Tosi, G.M.; Giordano, A.; et al. A Study of Quiet Eye’s Phenomenon in the Shooting Section of “Laser Run” of Modern Pentathlon. J. Cell. Physiol. 2019, 234, 9247–9254. [Google Scholar] [CrossRef] [PubMed]
- Janelle, C.M.; Fawver, B.J.; Beatty, G.F. Emotion and Sport Performance. In Handbook of Sport Psychology; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2020; pp. 254–298. ISBN 978-1-119-56812-4. [Google Scholar]
- Montuori, S.; D’Aurizio, G.; Foti, F.; Liparoti, M.; Lardone, A.; Pesoli, M.; Sorrentino, G.; Mandolesi, L.; Curcio, G.; Sorrentino, P. Executive Functioning Profiles in Elite Volleyball Athletes: Preliminary Results by a Sport-Specific Task Switching Protocol. Hum. Mov. Sci. 2019, 63, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Fontani, G.; Lodi, L.; Felici, A.; Migliorini, S.; Corradeschi, F. Attention in Athletes of High and Low Experience Engaged in Different Open Skill Sports. Percept. Mot. Skills 2006, 102, 791–805. [Google Scholar] [CrossRef]
- Voss, M.W.; Kramer, A.F.; Basak, C.; Prakash, R.S.; Roberts, B. Are Expert Athletes ‘Expert’ in the Cognitive Laboratory? A Meta-Analytic Review of Cognition and Sport Expertise. Appl. Cogn. Psychol. 2010, 24, 812–826. [Google Scholar] [CrossRef]
- Giancamilli, F.; Galli, F.; Chirico, A.; Fegatelli, D.; Mallia, L.; Palombi, T.; Cordone, S.; Alivernini, F.; Mandolesi, L.; Lucidi, F. When the Going Gets Tough, What Happens to Quiet Eye? The Role of Time Pressure and Performance Pressure during Basketball Free Throws. Psychol. Sport Exerc. 2022, 58, 102057. [Google Scholar] [CrossRef]
- Williams, A.M.; Jackson, R.C. Anticipation in Sport: Fifty Years on, What Have We Learned and What Research Still Needs to Be Undertaken? Psychol. Sport Exerc. 2019, 42, 16–24. [Google Scholar] [CrossRef]
- Vickers, J.N.; Reeves, M.-A.; Chambers, K.L.; Martell, S. Decision Training: Cognitive Strategies for Enhancing Motor Performance. In Skill Acquisition in Sport; Routledge: Abingdon, UK, 2004; ISBN 978-0-203-64656-4. [Google Scholar]
- Corbetta, M.; Shulman, G.L. Control of Goal-Directed and Stimulus-Driven Attention in the Brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef]
- Birmingham, E.; Kingstone, A. Human Social Attention. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2009; Volume 176, pp. 309–320. ISBN 978-0-444-53426-2. [Google Scholar]
- Federico, F. Natural Environment and Social Relationship in the Development of Attentional Network. Front. Psychol. 2020, 11, 1345. [Google Scholar] [CrossRef] [PubMed]
- Nummenmaa, L.; Calder, A.J. Neural Mechanisms of Social Attention. Trends Cogn. Sci. 2009, 13, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, F.; Ristic, J. How Attention Gates Social Interactions. Ann. New York Acad. Sci. 2018, 1426, 179–198. [Google Scholar] [CrossRef] [PubMed]
- Dalmaso, M.; Castelli, L.; Galfano, G. Social Modulators of Gaze-Mediated Orienting of Attention: A Review. Psychon. Bull. Rev. 2020, 27, 833–855. [Google Scholar] [CrossRef] [PubMed]
- Federico, F.; Marotta, A.; Martella, D.; Casagrande, M. Development in Attention Functions and Social Processing: Evidence from the Attention Network Test. Br. J. Dev. Psychol. 2017, 35, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Chaddock, L.; Pontifex, M.B.; Hillman, C.H.; Kramer, A.F. A Review of the Relation of Aerobic Fitness and Physical Activity to Brain Structure and Function in Children. J. Int. Neuropsychol. Soc. 2011, 17, 975–985. [Google Scholar] [CrossRef] [Green Version]
- Kramer, A.F.; Erickson, K.I. Effects of Physical Activity on Cognition, Well-Being, and Brain: Human Interventions. Alzheimer’s Dement. 2007, 3, S45–S51. [Google Scholar] [CrossRef]
- Federico, F.; Marotta, A.; Adriani, T.; Maccari, L.; Casagrande, M. Attention Network Test—The Impact of Social Information on Executive Control, Alerting and Orienting. Acta Psychol. 2013, 143, 65–70. [Google Scholar] [CrossRef]
- Rueda, M.R.; Posner, M.I.; Rothbart, M.K. Attentional Control and Self-Regulation. In Handbook of Self-Regulation: Research, Theory, and Applications; The Guilford Press: New York, NY, USA, 2004; pp. 283–300. ISBN 978-1-57230-991-3. [Google Scholar]
- Martella, D.; Casagrande, M.; Lupiáñez, J. Alerting, Orienting and Executive Control: The Effects of Sleep Deprivation on Attentional Networks. Exp. Brain Res. 2011, 210, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Geva, R.; Zivan, M.; Warsha, A.; Olchik, D. Alerting, Orienting or Executive Attention Networks: Differential Patters of Pupil Dilations. Front. Behav. Neurosci. 2013, 7, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dichter, G.S.; Belger, A. Social Stimuli Interfere with Cognitive Control in Autism. NeuroImage 2007, 35, 1219–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, R.T. Developmental Deficits in Social Perception in Autism: The Role of the Amygdala and Fusiform Face Area. Int. J. Dev. Neurosci. 2005, 23, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Bindemann, M.; Burton, A.M.; Langton, S.R.H.; Schweinberger, S.R.; Doherty, M.J. The Control of Attention to Faces. J. Vis. 2007, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Langton, S.R.H.; Law, A.S.; Burton, A.M.; Schweinberger, S.R. Attention Capture by Faces. Cognition 2008, 107, 330–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driver, J.; Davis, G.; Ricciardelli, P.; Kidd, P.; Maxwell, E.; Baron-Cohen, S. Gaze Perception Triggers Reflexive Visuospatial Orienting. Vis. Cogn. 1999, 6, 509–540. [Google Scholar] [CrossRef]
- Marotta, A.; Lupiáñez, J.; Casagrande, M. Investigating Hemispheric Lateralization of Reflexive Attention to Gaze and Arrow Cues. Brain Cogn. 2012, 80, 361–366. [Google Scholar] [CrossRef]
- Friesen, C.K.; Kingstone, A. The Eyes Have It! Reflexive Orienting Is Triggered by Nonpredictive Gaze. Psychon. Bull. Rev. 1998, 5, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Riechelmann, E.; Raettig, T.; Böckler, A.; Huestegge, L. Gaze Interaction: Anticipation-Based Control of the Gaze of Others. Psychol. Res. 2021, 85, 302–321. [Google Scholar] [CrossRef]
- Martell, S.G.; Vickers, J.N. Gaze Characteristics of Elite and Near-Elite Athletes in Ice Hockey Defensive Tactics. Hum. Mov. Sci. 2004, 22, 689–712. [Google Scholar] [CrossRef] [PubMed]
- Yarrow, K.; Brown, P.; Krakauer, J.W. Inside the Brain of an Elite Athlete: The Neural Processes That Support High Achievement in Sports. Nat. Rev. Neurosci. 2009, 10, 585–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzolatti, G.; Sinigaglia, C. The Mirror Mechanism: A Basic Principle of Brain Function. Nat. Rev. Neurosci. 2016, 17, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Aglioti, S.M.; Cesari, P.; Romani, M.; Urgesi, C. Action Anticipation and Motor Resonance in Elite Basketball Players. Nat. Neurosci. 2008, 11, 1109–1116. [Google Scholar] [CrossRef]
- Wu, Y.; Zeng, Y.; Zhang, L.; Wang, S.; Wang, D.; Tan, X.; Zhu, X.; Zhang, J.; Zhang, J. The Role of Visual Perception in Action Anticipation in Basketball Athletes. Neuroscience 2013, 237, 29–41. [Google Scholar] [CrossRef]
- Schumacher, N.; Schmidt, M.; Wellmann, K.; Braumann, K.-M. General Perceptual-Cognitive Abilities: Age and Position in Soccer. PLoS ONE 2018, 13, e0202627. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| No. (n) | Age (Mean ± SD) | BMI (Mean ± SD) | |
|---|---|---|---|
| Strikers | 38 | 21.01 ± 3.61 | 23.73 ± 1.43 |
| Midfielders | 32 | 20.03 ± 3.23 | 23.55 ± 2.34 |
| Defenders | 32 | 18.50 ± 1.34 | 22.75 ± 1.45 |
| Factors | F(df) | p | |
|---|---|---|---|
| Alerting | Stimulus | 0.092(1,99) | 0.762 |
| Role | 1.791(2,99) | 0.172 | |
| Stimulus × Role | 2.109(2,99) | 00.080 | |
| Orienting | Stimulus | 3.604(1,99) | 0.061 |
| Role | 0.888(2,99) | 0.416 | |
| Stimulus × Role | 1.109(2,99) | 0.334 | |
| Executive Control | Stimulus | 6.761(1,99) | 0.011 * |
| Role | 1.081(2,99) | 0.343 | |
| Stimulus × Role | 2.921(2,99) | 0.059 |
| Factors | F(df) | p | |
|---|---|---|---|
| Alerting | Stimulus | 0.087(1,99) | 0.923 |
| Role | 17.306(2,99) | <0.001 *** | |
| Stimulus * Role | 0.098(2,99) | 0.751 | |
| Orienting | Stimulus | 0.684(1,99) | 0.410 |
| Role | 0.750(2,99) | 0.475 | |
| Stimulus * Role | 6.022(2,99) | 0.003 ** | |
| Executive Control | Stimulus | 0.074(1,99) | 0.687 |
| Role | 0.429(2,99) | 0.635 | |
| Stimulus * Role | 7.089(2,99) | 0.024 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Passarello, N.; Mellone, M.; Sorrentino, P.; Chirico, A.; Lucidi, F.; Mandolesi, L.; Federico, F. The Effects of Social Processing and Role Type on Attention Networks: Insights from Team Ball Athletes. Brain Sci. 2023, 13, 476. https://doi.org/10.3390/brainsci13030476
Passarello N, Mellone M, Sorrentino P, Chirico A, Lucidi F, Mandolesi L, Federico F. The Effects of Social Processing and Role Type on Attention Networks: Insights from Team Ball Athletes. Brain Sciences. 2023; 13(3):476. https://doi.org/10.3390/brainsci13030476
Chicago/Turabian StylePassarello, Noemi, Michela Mellone, Pierpaolo Sorrentino, Andrea Chirico, Fabio Lucidi, Laura Mandolesi, and Francesca Federico. 2023. "The Effects of Social Processing and Role Type on Attention Networks: Insights from Team Ball Athletes" Brain Sciences 13, no. 3: 476. https://doi.org/10.3390/brainsci13030476