
An EEG Study of the Influence of Target Appearing in the Upper and Lower Visual Fields on Brain Attention Resource Allocation
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
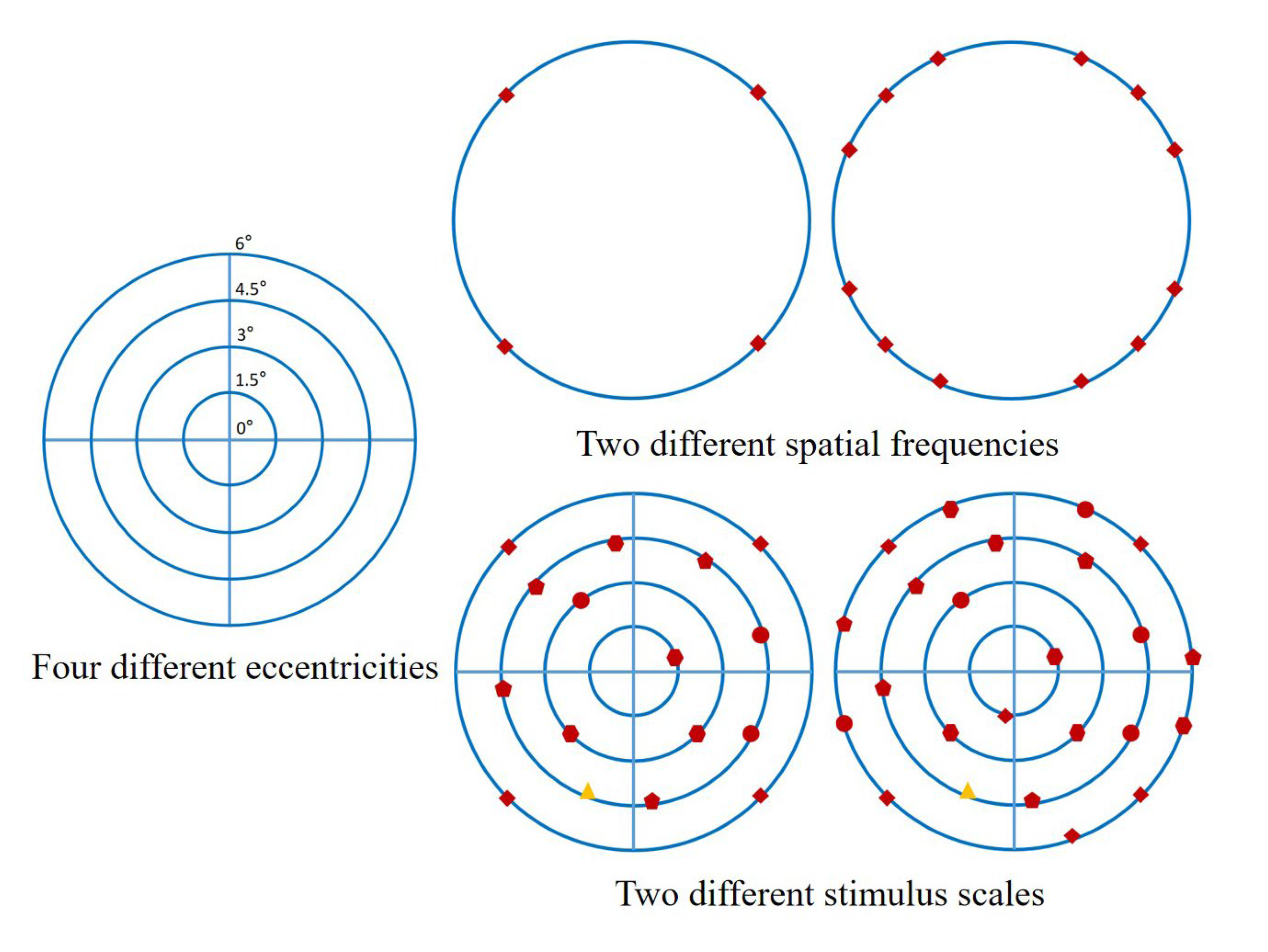
2.2. Experimental Paradigms
2.3. Experiment Procedures
2.4. Data Acquisition
2.5. Date Analysis
2.6. Statistical Analysis
3. Results
3.1. Exp.1 Behavioral Results
3.2. Exp.1 Time-Domain Results
3.3. Exp.1 Time-Frequency Results
3.4. Exp.2 Behavioral Results
3.5. Exp.2 Time-Domain Results
3.6. Exp.2 Time-Frequency Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Q.; Liang, T.; Zhang, J.; Fu, X.; Wu, J. Electrophysiological evidence for temporal dynamics associated with attentional processing in the zoom lens paradigm. PeerJ 2018, 6, e4538. [Google Scholar] [CrossRef] [Green Version]
- Posner, M.I. Orienting of Attention. Q. J. Exp. Psychol. 1980, 32, 3–25. [Google Scholar] [CrossRef]
- Stapleton, T.; Ashburn, A.; Stack, E. A pilot study of attention deficits, balance control and falls in the subacute stage following stroke. Clin. Rehabil. 2001, 15, 437–444. [Google Scholar] [CrossRef]
- Egelko, S.; Riley, E.; Simon, D.; Diller, L.; Ezrachi, O. Unilateral spatial neglect: Biases in contralateral search and fine spatial attention. Arch. Clin. Neuropsychol. 1988, 3, 213–225. [Google Scholar] [CrossRef]
- Ptak, R.; Schnider, A. The attention network of the human brain: Relating structural damage associated with spatial neglect to functional imaging correlates of spatial attention. Neuropsychologia 2011, 49, 3063–3070. [Google Scholar] [CrossRef]
- Wang, J.; Wang, W.; Hou, Z.-G. Toward Improving Engagement in Neural Rehabilitation: Attention Enhancement Based on Brain–Computer Interface and Audiovisual Feedback. IEEE Trans. Cogn. Dev. Syst. 2020, 12, 787–796. [Google Scholar] [CrossRef]
- Vinci-Booher, S.; Caron, B.; Bullock, D.; James, K.; Pestilli, F. Development of white matter tracts between and within the dorsal and ventral streams. Anat. Embryol. 2022, 227, 1457–1477. [Google Scholar] [CrossRef]
- Previc, F.H. Functional specialization in the lower and upper visual fields in humans: Its ecological origins and neurophysiological implications. Behav. Brain Sci. 1990, 13, 519–542. [Google Scholar] [CrossRef]
- Goodale, M.A.; Danckert, J. Superior performance for visually guided pointing in the lower visual field. Exp. Brain Res. 2001, 137, 303–308. [Google Scholar] [CrossRef]
- Genzano, V.R.; Di Nocera, F.; Ferlazzo, F. Upper/lower visual field asymmetry on a spatial relocation memory task. Neuroreport 2001, 12, 1227–1230. [Google Scholar] [CrossRef] [Green Version]
- Previc, F.H.; Blume, J.L. Visual search asymmetries in three-dimensional space. Vis. Res. 1993, 33, 2697–2704. [Google Scholar] [CrossRef]
- Zhou, W.; King, W. Attentional sensitivity and asymmetries of vertical saccade generation in monkey. Vis. Res. 2002, 42, 771–779. [Google Scholar] [CrossRef] [Green Version]
- Hafed, Z.M.; Chen, C.-Y. Sharper, Stronger, Faster Upper Visual Field Representation in Primate Superior Colliculus. Curr. Biol. 2016, 26, 1647–1658. [Google Scholar] [CrossRef] [Green Version]
- Edgar, G.K.; Smith, A.T. Hemifield Differences in Perceived Spatial Frequency. Perception 1990, 19, 759–766. [Google Scholar] [CrossRef]
- Carrasco, M.; Evert, D.L.; Chang, I.; Katz, S.M. The eccentricity effect: Target eccentricity affects performance on conjunction searches. Percept. Psychophys. 1995, 57, 1241–1261. [Google Scholar] [CrossRef] [Green Version]
- Carrasco, M.; McLean, T.L.; Katz, S.M.; Frieder, K.S. Feature asymmetries in visual search: Effects of display duration, target eccentricity, orientation and spatial frequency. Vis. Res. 1998, 38, 347–374. [Google Scholar] [CrossRef] [Green Version]
- Lum, J.A.G.; Lammertink, I.; Clark, G.; Fuelscher, I.; Hyde, C.; Enticott, P.; Ullman, M.T. Visuospatial sequence learning on the serial reaction time task modulates the P1 event-related potential. Psychophysiology 2019, 56, e13292. [Google Scholar] [CrossRef]
- Song, W.; Gao, Y.; Luo, Y. Early scale effect and hemisphere superiority on the visual spatial attention: From the electrophysiological evidence of ERP. Prog. Nat. Sci. 2004, 14, 875–879. [Google Scholar] [CrossRef]
- Spyropoulos, G.; Bosman, C.A.; Fries, P. A theta rhythm in macaque visual cortex and its attentional modulation. CSHL 2017, 115, E5614–E5623. [Google Scholar] [CrossRef] [Green Version]
- Neuper, C.; Pfurtscheller, G. Event-related dynamics of cortical rhythms: Frequency-specific features and functional correlates. Int. J. Psychophysiol. 2001, 43, 41–58. [Google Scholar] [CrossRef]
- Sauseng, P.; Klimesch, W.; Stadler, W.; Schabus, M.; Doppelmayr, M.; Hanslmayr, S.; Gruber, W.R.; Birbaumer, N. A shift of visual spatial attention is selectively associated with human EEG alpha activity. Eur. J. Neurosci. 2005, 22, 2917–2926. [Google Scholar] [CrossRef]
- Thut, G.; Nietzel, A.; Brandt, S.A.; Pascual-Leone, A. α-Band Electroencephalographic Activity over Occipital Cortex Indexes Visuospatial Attention Bias and Predicts Visual Target Detection. J. Neurosci. 2006, 26, 9494–9502. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, W.; Hou, Z.-G.; Shi, W.; Liang, X.; Ren, S.; Peng, L.; Zhou, Y.-J. BCI and Multimodal Feedback Based Attention Regulation for Lower Limb Rehabilitation. In Proceedings of the 2019 International Joint Conference on Neural Networks (IJCNN), Budapest, Hungary, 14–19 July 2019. [Google Scholar] [CrossRef]
- Wang, J.; Wang, W.; Ren, S.; Shi, W.; Hou, Z.-G. Engagement Enhancement Based on Human-in-the-Loop Optimization for Neural Rehabilitation. Front. Neurorobotics 2020, 14, 596019. [Google Scholar] [CrossRef]
- Wang, J.; Wang, W.; Hou, Z.-G. EEG-Based Focus of Attention Tracking and Regulation During Dual-Task Training for Neural Rehabilitation of Stroke Patients. IEEE Trans. Biomed. Eng. 2022, 70, 920–930. [Google Scholar] [CrossRef]
- Dombrowe, I.; Hilgetag, C.C. Occipitoparietal alpha-band responses to the graded allocation of top-down spatial attention. J. Neurophysiol. 2014, 112, 1307–1316. [Google Scholar] [CrossRef]
- Bonacci, L.M.; Bressler, S.; Kwasa, J.A.C.; Noyce, A.L.; Shinn-Cunningham, B.G. Effects of Visual Scene Complexity on Neural Signatures of Spatial Attention. Front. Hum. Neurosci. 2020, 14, 91. [Google Scholar] [CrossRef] [Green Version]
- Bacigalupo, F.; Luck, S.J. Lateralized Suppression of Alpha-Band EEG Activity As a Mechanism of Target Processing. J. Neurosci. 2019, 39, 900–917. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhao, C.; Guo, J.; Kong, Y.; Li, H.; Du, B.; Ding, Y.; Song, Y. Visual Working Memory Guides Spatial Attention: Evidence from alpha oscillations and sustained potentials. Neuropsychologia 2021, 151, 107719. [Google Scholar] [CrossRef]
- Qu, Z.; Song, Y.; Ding, Y. Asymmetry between the upper and lower visual fields: An event-related potential study. Chin. Sci. Bull. 2006, 51, 536–541. [Google Scholar] [CrossRef]
- Clark, V.P.; Fan, S.; Hillyard, S.A. Identification of early visual evoked potential generators by retinotopic and topographic analyses. Hum. Brain Mapp. 1994, 2, 170–187. [Google Scholar] [CrossRef]
- Gunter, T.C.; Wijers, A.A.; Jackson, J.L.; Mulder, G. Visual spatial attention to stimuli presented on the vertical and horizontal meridian: An ERP study. Psychophysiology 1994, 31, 140–153. [Google Scholar] [CrossRef] [Green Version]
- Ladouce, S.; Donaldson, D.I.; Dudchenko, P.A.; Ietswaart, M. Mobile EEG identifies the re-allocation of attention during real-world activity. Sci. Rep. 2019, 9, 15851. [Google Scholar] [CrossRef] [Green Version]
- Bosworth, R.G.; Petrich, J.A.; Dobkins, K.R. Effects of spatial attention on motion discrimination are greater in the left than right visual field. Vis. Res. 2012, 52, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Proverbio, A.M.; Zani, A.; Avella, C. Hemispheric Asymmetries for Spatial Frequency Discrimination in a Selective Attention Task. Brain Cogn. 1997, 34, 311–320. [Google Scholar] [CrossRef]
- Asanowicz, D.; Kruse, L.; Śmigasiewicz, K.; Verleger, R. Lateralization of spatial rather than temporal attention underlies the left hemifield advantage in rapid serial visual presentation. Brain Cogn. 2017, 118, 54–62. [Google Scholar] [CrossRef]
- Bergerbest, D.; Shilkrot, O.; Joseph, M.; Salti, M. Right visual-field advantage in the attentional blink: Asymmetry in attentional gating across time and space. Atten. Percept. Psychophys. 2017, 79, 1979–1992. [Google Scholar] [CrossRef]
- Carlei, C.; Kerzel, D. Stronger interference from distractors in the right hemifield during visual search. Laterality 2017, 23, 152–165. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Li, R.; Ming, D. Review on the relationship between selective attention and neural oscillations. J. Biomed. Eng. 2019, 36, 320–324. [Google Scholar]
- Lu, R.; Xi, J.; Zhang, X.; Shi, J. High fluid intelligence is characterized by flexible allocation of attentional resources: Evidence from EEG. Neuropsychologia 2021, 164, 108094. [Google Scholar] [CrossRef]
- Carrasco, M.; Talgar, C.P.; Cameron, E.L. Characterizing visual performance fields: Effects of transient covert attention, spatial frequency, eccentricity, task and set size. Spat. Vis. 2001, 15, 61–75. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Xu, J.; Sheng, J.; Zhou, H.; Liu, Z.; Li, Y.; Hu, J.; Zuo, G. An EEG Study of the Influence of Target Appearing in the Upper and Lower Visual Fields on Brain Attention Resource Allocation. Brain Sci. 2023, 13, 433. https://doi.org/10.3390/brainsci13030433
Li H, Xu J, Sheng J, Zhou H, Liu Z, Li Y, Hu J, Zuo G. An EEG Study of the Influence of Target Appearing in the Upper and Lower Visual Fields on Brain Attention Resource Allocation. Brain Sciences. 2023; 13(3):433. https://doi.org/10.3390/brainsci13030433
Chicago/Turabian StyleLi, Han, Jialin Xu, Junpeng Sheng, Huilin Zhou, Zhen Liu, Yahui Li, Jingyan Hu, and Guokun Zuo. 2023. "An EEG Study of the Influence of Target Appearing in the Upper and Lower Visual Fields on Brain Attention Resource Allocation" Brain Sciences 13, no. 3: 433. https://doi.org/10.3390/brainsci13030433