
Intermittent Exposure to Aflatoxin B1 Did Not Affect Neurobehavioral Parameters and Biochemical Markers of Oxidative Stress
 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Animals
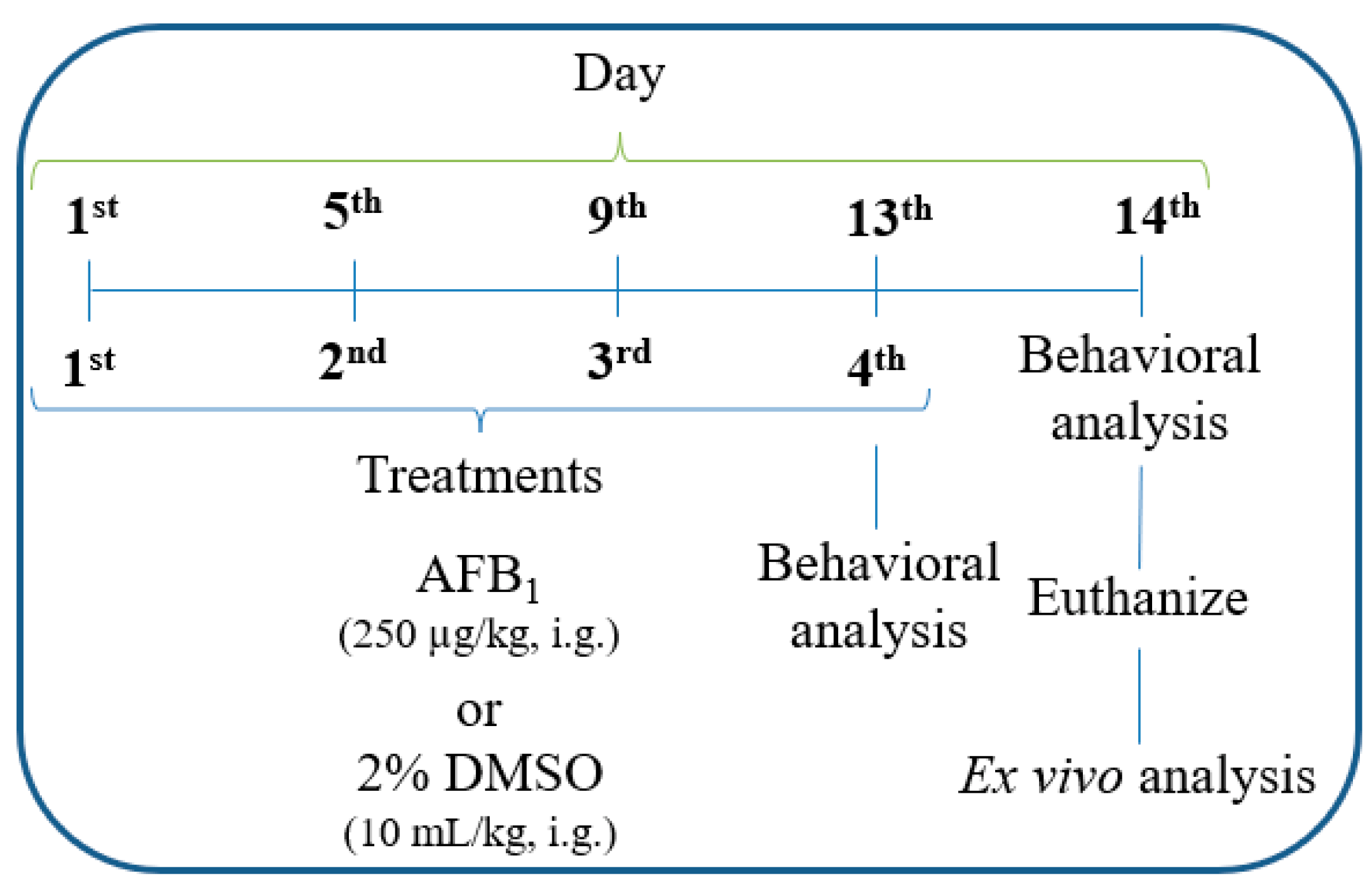
2.2. Experimental Design
2.3. Behavioral Analysis
2.3.1. Object Recognition Test
2.3.2. Open Field Test
2.3.3. Marble Burying Test
2.3.4. Nest Construction Test
2.3.5. Splash Test
2.4. Ex Vivo Analysis
2.4.1. Plasma Biochemical Analysis
2.4.2. Protein Determination
2.4.3. NPSH Levels
2.4.4. Ascorbic Acid (AA) Levels
2.4.5. TBARS Levels
2.4.6. FRAP Potential
2.4.7. Na+, K+-ATPase Activity
2.4.8. CAT Activity
2.4.9. GST Activity
2.4.10. Western Blot
2.5. Statistical Analysis
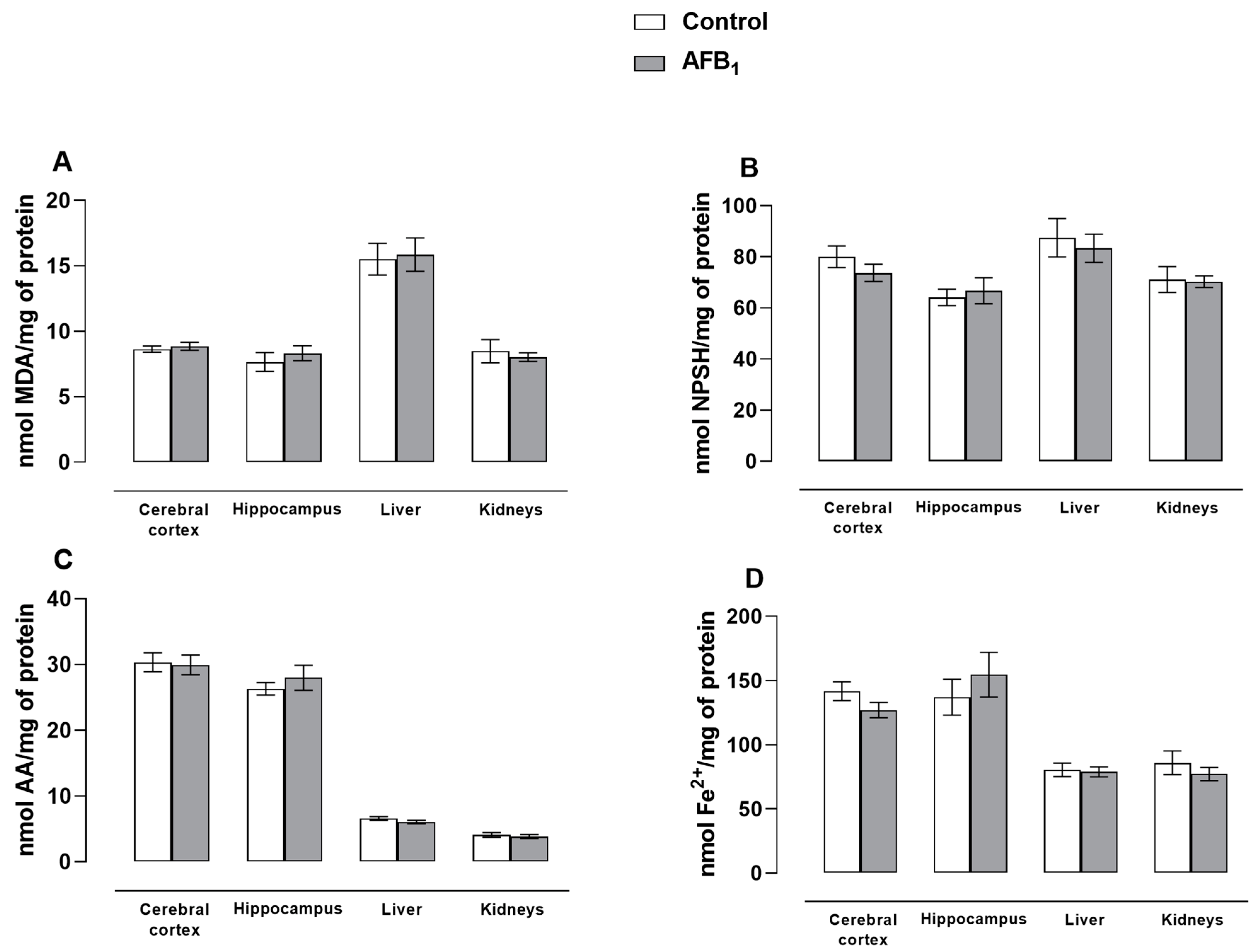
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patriarca, A.; Fernández Pinto, V. Prevalence of mycotoxins in foods and decontamination. Curr. Opin. Food Sci. 2017, 14, 50–60. [Google Scholar] [CrossRef]
- Ruadrew, S.; Craft, J.; Aidoo, K. Occurrence of toxigenic Aspergillus spp. and aflatoxins in selected food commodities of Asian origin sourced in the West of Scotland. Food Chem. Toxicol. 2013, 55, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Benford, D.; Leblanc, J.-C.; Setzer, R.W. Application of the margin of exposure (MoE) approach to substances in food that are genotoxic and carcinogenic: Example: Aflatoxin B1 (AFB1). Food Chem. Toxicol. 2010, 48, S34–S41. [Google Scholar] [CrossRef]
- Udomkun, P.; Wiredu, A.N.; Nagle, M.; Muller, J.; Vanlauwe, B.; Bandyopadhyay, R. Innovative technologies to manage aflatoxins in foods and feeds and the profitability of application—A review. Food Control 2017, 76, 127–138. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; International Agency for Research on Cancer: Lyon, France, 2012; Volume 100F, pp. 225–244. [Google Scholar]
- Stein, R.A.; Bulboacă, A.E. Chapter 21—Mycotoxins. In Foodborne Diseases, 3rd ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 407–446. [Google Scholar]
- Andrade, P.D.; de Mello, M.H.; Franca, J.A.; Caldas, E.D. Aflatoxins in food products consumed in Brazil: A preliminary dietary risk assessment. Food Addit. Contam. Part A 2013, 30, 127–136. [Google Scholar] [CrossRef]
- Popkin, B.M. Global nutrition dynamics: The world is shifting rapidly toward a diet linked with noncommunicable diseases. Am. J. Clin. Nutr. 2006, 84, 289–298. [Google Scholar] [CrossRef]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef]
- Pankaj, S.K.; Shi, H.; Keener, K.M. A review of novel physical and chemical decontamination technologies for aflatoxin in food. Trends Food Sci. Technol. 2018, 71, 73–83. [Google Scholar] [CrossRef]
- Raksha Rao, K.; Vipin, A.V.; Hariprasad, P.; Anu Appaiah, K.A.; Venkateswaran, G. Biological detoxification of Aflatoxin B1 by Bacillus licheniformis CFR1. Food Control 2017, 71, 234–241. [Google Scholar] [CrossRef]
- Sun, X.; Sun, C.; Zhang, X.; Zhang, H.; Ji, J.; Liu, Y.; Tang, L. Aflatoxin B1 decontamination by UV-mutated live and immobilized Aspergillus niger. Food Control 2016, 61, 235–242. [Google Scholar] [CrossRef]
- Adedara, I.A.; Owumi, S.E.; Oyelere, A.K.; Farombi, E.O. Neuroprotective role of gallic acid in aflatoxin B1 -induced behavioral abnormalities in rats. J. Biochem. Mol. Toxicol. 2021, 35, e22684. [Google Scholar] [CrossRef]
- Qian, G.; Wang, F.; Tang, L.; Massey, M.E.; Mitchell, N.J.; Su, J.; Williams, J.H.; Phillips, T.D.; Wang, J.S. Integrative toxicopathological evaluation of aflatoxin B(1) exposure in F344 rats. Toxicol. Pathol. 2013, 41, 1093–1105. [Google Scholar] [CrossRef] [Green Version]
- Souto, N.S.; Claudia Monteiro Braga, A.; Lutchemeyer de Freitas, M.; Rechia Fighera, M.; Royes LF, F.; Schneider Oliveira, M.; Furian, A.F. Aflatoxin B1 reduces non-enzymatic antioxidant defenses and increases protein kinase C activation in the cerebral cortex of young rats. Nutr. Neurosci. 2018, 21, 268–275. [Google Scholar] [CrossRef]
- Wei, W.; Li, R.; He, L.; Wang, X.; Ji, X.; Zhou, Y. Evaluation of toxicological impacts on Sprague-Dawley rat by successively low dose of aflatoxin B1. J. Sci. Food Agric. 2014, 94, 3017–3022. [Google Scholar] [CrossRef]
- Trebak, F.; Alaoui, A.; Alexandre, D.; El Ouezzani, S.; Anouar, Y.; Chartrel, N.; Magoul, R. Impact of aflatoxin B1 on hypothalamic neuropeptides regulating feeding behavior. Neurotoxicology 2015, 49, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, A.R.; Rosa, É.V.F.; Sari, M.H.M.; Sampaio, T.B.; Dos Santos, J.T.; Jardim, N.S.; Müller, S.G.; Oliveira, M.S.; Nogueira, C.W.; Furian, A.F. Therapeutic potential of beta-caryophyllene against aflatoxin B1-Induced liver toxicity: Biochemical and molecular insights in rats. Chem. Biol. Interact. 2021, 348, 109635. [Google Scholar] [CrossRef] [PubMed]
- Souto, N.S.; Dassi, M.; Braga, A.C.M.; Rosa, E.V.F.; Fighera, M.R.; Royes, L.F.F.; Oliveira, M.S.; Furian, A.F. Behavioural and biochemical effects of one-week exposure to aflatoxin B1 and aspartame in male Wistar rats. World Mycotoxin J. 2019, 12, 293–305. [Google Scholar] [CrossRef]
- De Oliveira, C.C.; de Oliveira, C.V.; Grigoletto, J.; Ribeiro, L.R.; Funck, V.R.; Grauncke, A.C.; de Souza, T.L.; Souto, N.S.; Furian, A.F.; Menezes, I.R.; et al. Anticonvulsant activity of beta-caryophyllene against pentylenetetrazol-induced seizures. Epilepsy Behav. 2016, 56, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Sanathara, N.M.; Garau, C.; Alachkar, A.; Wang, L.; Wang, Z.; Nishimori, K.; Xu, X.; Civelli, O. Melanin concentrating hormone modulates oxytocin-mediated marble burying. Neuropharmacology 2017, 128, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Deacon, R. Assessing burrowing, nest construction, and hoarding in mice. J. Vis. Exp. 2012, 59, e2607. [Google Scholar] [CrossRef] [Green Version]
- Pesarico, A.P.; Sartori, G.; Bruning, C.A.; Mantovani, A.C.; Duarte, T.; Zeni, G.; Nogueira, C.W. A novel isoquinoline compound abolishes chronic unpredictable mild stress-induced depressive-like behavior in mice. Behav. Brain Res. 2016, 307, 73–83. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Jacques-Silva, M.C.; Nogueira, C.W.; Broch, L.C.; Flores, E.M.; Rocha, J.B. Diphenyl diselenide and ascorbic acid changes deposition of selenium and ascorbic acid in liver and brain of mice. Pharm. Toxicol. 2001, 88, 119–125. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of "antioxidant power": The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Funck, V.R.; Ribeiro, L.R.; Pereira, L.M.; de Oliveira, C.V.; Grigoletto, J.; Fighera, M.R.; Royes, L.F.; Furian, A.F.; Oliveira, M.S. Long-term decrease in Na+,K+-ATPase activity after pilocarpine-induced status epilepticus is associated with nitration of its alpha subunit. Epilepsy Res. 2014, 108, 1705–1710. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzym. 1984, 105, 121–126. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Bagchi, M.; Zafra-Stone, S.; Bagchi, D.; Patel, S. Chapter 27—Oxidative stress and chemical toxicity A2—Gupta, Ramesh C. In Veterinary Toxicology, 2nd ed.; Academic Press: Boston, MA, USA, 2012; pp. 426–439. [Google Scholar]
- Godfrey, S.; Kitya, D.; Lubega, A.; Ogwal-Okeng, J.; William, W.; David, B. Review of the Biological and Health Effects of Aflatoxins on Body Organs and Body Systems. Aflatoxins—Recent Adv. Future Prospect. 2013, 12, 239–265. [Google Scholar]
- Song, C.; Yang, J.; Wang, Y.; Ding, G.; Guo, L.; Qin, J. Mechanisms and transformed products of aflatoxin B1 degradation under multiple treatments: A review. Crit. Rev. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef] [PubMed]
- Urrego Novoa, J.R.; Díaz, G.J. Aflatoxinas: Mecanismos de toxicidad en la etiología de cáncer hepático celular. Rev. Fac. Med. 2006, 54, 108–116. [Google Scholar]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2007, 22, 659–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Human Organization (WHO). Evaluation of Certain Contaminants in Food; WHO Technical Reports Series; WHO: Geneva, Switzerland, 2017; pp. 40–55. [Google Scholar]
- Kujbida, P.; Maia, P.P.; de Araújo, A.N.; Mendes, L.D.; de Oliveira, M.L.; Silva-Rocha, W.P.; de Brito, G.Q.; Chaves, G.M.; Martins, I. Risk assessment of the occurrence of aflatoxin and fungi in peanuts and cashew nuts. Braz. J. Pharm. Sci. 2019, 55, e18135. [Google Scholar] [CrossRef]
- Souto, N.S.; Dassi, M.; Braga, A.C.M.; Rosa, E.V.F.; Fighera, M.R.; Royes, L.F.F.; Oliveira, M.S.; Sari, M.H.M.; Furian, A.F. Hepatic susceptibility to oxidative damage after repeated concomitant exposure to aspartame and aflatoxin B1 in rats. Drug Chem. Toxicol. 2022, 45, 2780–2785. [Google Scholar] [CrossRef]
- El-Barbary, M.I. Evaluation of the efficacy of glutathione against aflatoxicosis in Oreochromis niloticus on hepatic antioxidants and gene expression of CYP1A and GPx. Egypt. J. Aquat. Res. 2015, 41, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wang, L.; Zheng, C.; Liu, L.; Wang, J.; Li, D.; Tan, Y.; Zhao, X.; He, L.; Shu, W. Microcystin-LR increases genotoxicity induced by aflatoxin B1 through oxidative stress and DNA base excision repair genes in human hepatic cell lines. Environ. Pollut. 2018, 233, 455–463. [Google Scholar] [CrossRef]
- Mahfouz, M.E.; Sherif, A.H. A multiparameter investigation into adverse effects of aflatoxin on Oreochromis niloticus health status. J. Basic Appl. Zool. 2015, 71, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Hamid, A.A.M.; Firgany, A.E.-D.L. Vitamin E supplementation ameliorates aflatoxin B1-induced nephrotoxicity in rats. Acta Histochem. 2015, 117, 767–779. [Google Scholar] [CrossRef]
- Ikegwuonu, F.I. The neurotoxicity of aflatoxin B1 in the rat. Toxicology 1983, 28, 247–259. [Google Scholar] [CrossRef]
- Qureshi, H.; Hamid, S.S.; Ali, S.S.; Anwar, J.; Siddiqui, A.A.; Khan, N.A. Cytotoxic effects of aflatoxin B1 on human brain microvascular endothelial cells of the blood-brain barrier. Med. Mycol. 2015, 53, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Supriya, C.; Reddy, P.S. Prenatal exposure to aflatoxin B1: Developmental, behavioral, and reproductive alterations in male rats. Naturwissenschaften 2015, 102, 26. [Google Scholar] [CrossRef]
- Bahey, N.G.; Elaziz, H.O.A.; Gadalla, K.K.E.S. Toxic effect of aflatoxin B1 and the role of recovery on the rat cerebral cortex and hippocampus. Tissue Cell 2015, 47, 559–566. [Google Scholar] [CrossRef]
- Bruneau, J.C.; Stack, E.; O’Kennedy, R.; Loscher, C.E. Aflatoxins B1, B2 and G1 modulate cytokine secretion and cell surface marker expression in J774A.1 murine macrophages. Toxicol. Vitr. 2012, 26, 686–693. [Google Scholar] [CrossRef]
- Kielian, T. Toll-like receptors in central nervous system glial inflammation and homeostasis. J. Neurosci. Res. 2006, 83, 711–730. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, A.; Mehrzad, J.; Mahmoudi, M.; Schneider, M. Environmentally Relevant Level of Aflatoxin B1 Dysregulates Human Dendritic Cells through Signaling on Key Toll-Like Receptors. Int. J. Toxicol. 2014, 33, 175–186. [Google Scholar] [CrossRef]
- Coulombe, R.A.; Sharma, R.P. Effect of repeated dietary exposure of aflatoxin B1~on brain biogenic amines and metabolites in the rat. Toxicol. Appl. Pharmacol. 1985, 80, 496–501. [Google Scholar] [CrossRef]
- Benarroch, E.E. Na+, K+-ATPase: Functions in the nervous system and involvement in neurologic disease. Neurology 2011, 76, 287–293. [Google Scholar] [CrossRef]
- Isakov, N. Protein kinase C (PKC) isoforms in cancer, tumor promotion and tumor suppression. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Furian, A.F.; Fighera, M.R.; Royes, L.F.; Oliveira, M.S. Recent advances in assessing the effects of mycotoxins using animal models. Curr. Opin. Food Sci. 2022, 47, 100874. [Google Scholar] [CrossRef]
- Bozzo, G.; Pugliese, N.; Samarelli, R.; Schiavone, A.; Dimuccio, M.M.; Circella, E.; Bonerba, E.; Ceci, E.; Camarda, A. Ochratoxin A and Aflatoxin B1 Detection in Laying Hens for Omega 3-Enriched Eggs Production. Agriculture 2023, 13, 138. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DMSO | AFB1 | p Value | |
|---|---|---|---|
| GST (nmol CDNB/mg protein/min) | |||
| Liver | 61.13 ± 9.43 | 50.10 ± 8.30 | 0.39 |
| Kidneys | 15.07 ± 2.17 | 10.13 ± 1.37 | 0.07 |
| CAT (K/mg protein) | |||
| Liver | 1.47 ± 0.15 | 1.95 ± 0.24 | 0.12 |
| Kidneys | 0.71 ± 0.08 | 0.59 ± 0.08 | 0.34 |
| DMSO | AFB1 | p Value | |
|---|---|---|---|
| ALT (U/L) | 48.88 ± 3.44 | 56.25 ± 4.92 | 0.23 |
| AST (U/L) | 249.30 ± 14.69 | 284.80 ± 18.78 | 0.15 |
| CREA (mg/dL) | 0.52 ± 0.01 | 0.52 ± 0.02 | 0.80 |
| ALB (g/L) | 2.95 ± 0.11 | 2.98 ± 0.04 | 0.76 |
| TBARS (nmol MDA/mg protein) | 3.91 ± 0.21 | 4.06 ± 0.70 | 0.83 |
| FRAP (nmol Fe2+/mg protein) | 11.95 ± 0.36 | 14.77 ± 1.61 | 0.11 |
| DMSO | AFB1 | p Value | |
|---|---|---|---|
| Marble Burying | |||
| Buried marbles (number) | 3.75 ± 0.70 | 4.00 ± 0.82 | 0.82 |
| Nest Test | |||
| Nest score | 4.25 ± 0.36 | 3.25 ± 0.45 | 0.10 |
| Splash Test | |||
| Latency to grooming (s) | 53.38 ± 6.56 | 53.00 ± 9.30 | 0.97 |
| Grooming (s) | 115.10 ± 24.11 | 137.90 ± 18.87 | 0.46 |
| Object Recognition | |||
| Short-term memory | 0.56 ± 0.08 | 0.59 ± 0.06 | 0.80 |
| Long-term memory | 0.53 ± 0.04 | 0.48 ± 0.05 | 0.48 |
| Open Field | |||
| Latency to explore (s) | 2.50 ± 0.56 | 2.25 ± 0.59 | 0.76 |
| Time spent in center (%) | 1.84 ± 0.49 | 3.03 ± 0.96 | 0.28 |
| Time spent in periphery (%) | 98.16 ± 0.49 | 96.97 ± 0.96 | 0.28 |
| Crossings (number) | 51.50 ± 4.96 | 64.50 ± 8.03 | 0.19 |
| Rearings (number) | 24.38 ± 2.79 | 29.00 ± 5.47 | 0.46 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braga, A.C.M.; Souto, N.S.; Cabral, F.L.; Dassi, M.; Rosa, É.V.F.; Guarda, N.d.S.; Royes, L.F.F.; Fighera, M.R.; Moresco, R.N.; Oliveira, M.S.; et al. Intermittent Exposure to Aflatoxin B1 Did Not Affect Neurobehavioral Parameters and Biochemical Markers of Oxidative Stress. Brain Sci. 2023, 13, 386. https://doi.org/10.3390/brainsci13030386
Braga ACM, Souto NS, Cabral FL, Dassi M, Rosa ÉVF, Guarda NdS, Royes LFF, Fighera MR, Moresco RN, Oliveira MS, et al. Intermittent Exposure to Aflatoxin B1 Did Not Affect Neurobehavioral Parameters and Biochemical Markers of Oxidative Stress. Brain Sciences. 2023; 13(3):386. https://doi.org/10.3390/brainsci13030386
Chicago/Turabian StyleBraga, Ana Claudia Monteiro, Naieli Schiefelbein Souto, Fernanda Licker Cabral, Micheli Dassi, Érica Vanessa Furlan Rosa, Naiara dos Santos Guarda, Luiz Fernando Freire Royes, Michele Rechia Fighera, Rafael Noal Moresco, Mauro Schneider Oliveira, and et al. 2023. "Intermittent Exposure to Aflatoxin B1 Did Not Affect Neurobehavioral Parameters and Biochemical Markers of Oxidative Stress" Brain Sciences 13, no. 3: 386. https://doi.org/10.3390/brainsci13030386