
Integrative Analysis Reveals the Expression Pattern of SOX9 in Satellite Glial Cells after Sciatic Nerve Injury
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Extraction
2.2. DEGs Analysis
2.3. Functional Enrichment Analysis of DEGs
2.4. Protein–Protein Interactions (PPIs) Analysis of DEGs
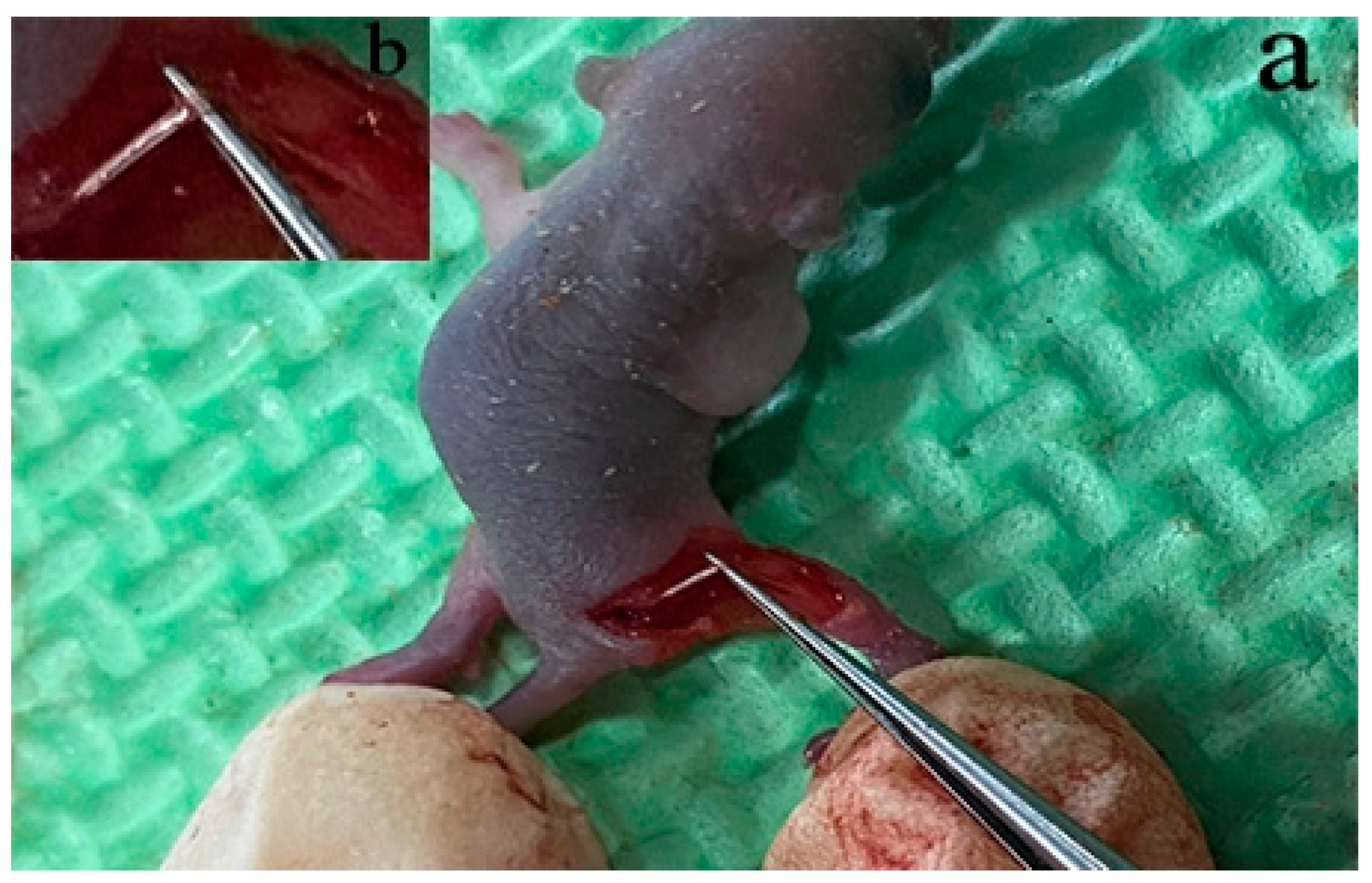
2.5. SNI Model in Mice
2.6. Cell Culture
2.7. Immunofluorescence Staining
2.8. Real-Time (RT) Polymerase Chain Reaction Assay
2.9. Western Blotting Assay
2.10. Statistical Analysis
3. Results
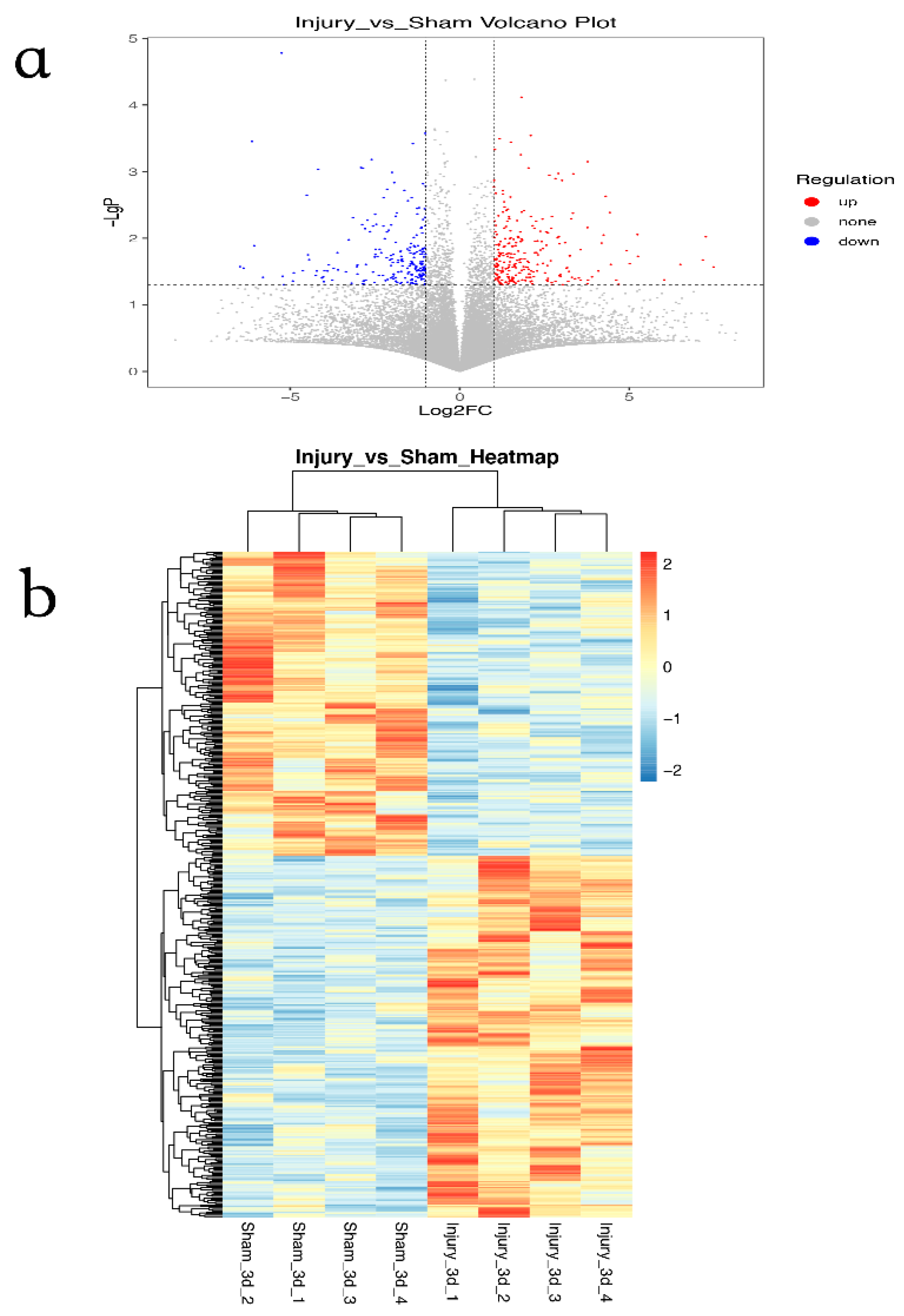
3.1. Identification of DEGs in GSE120284
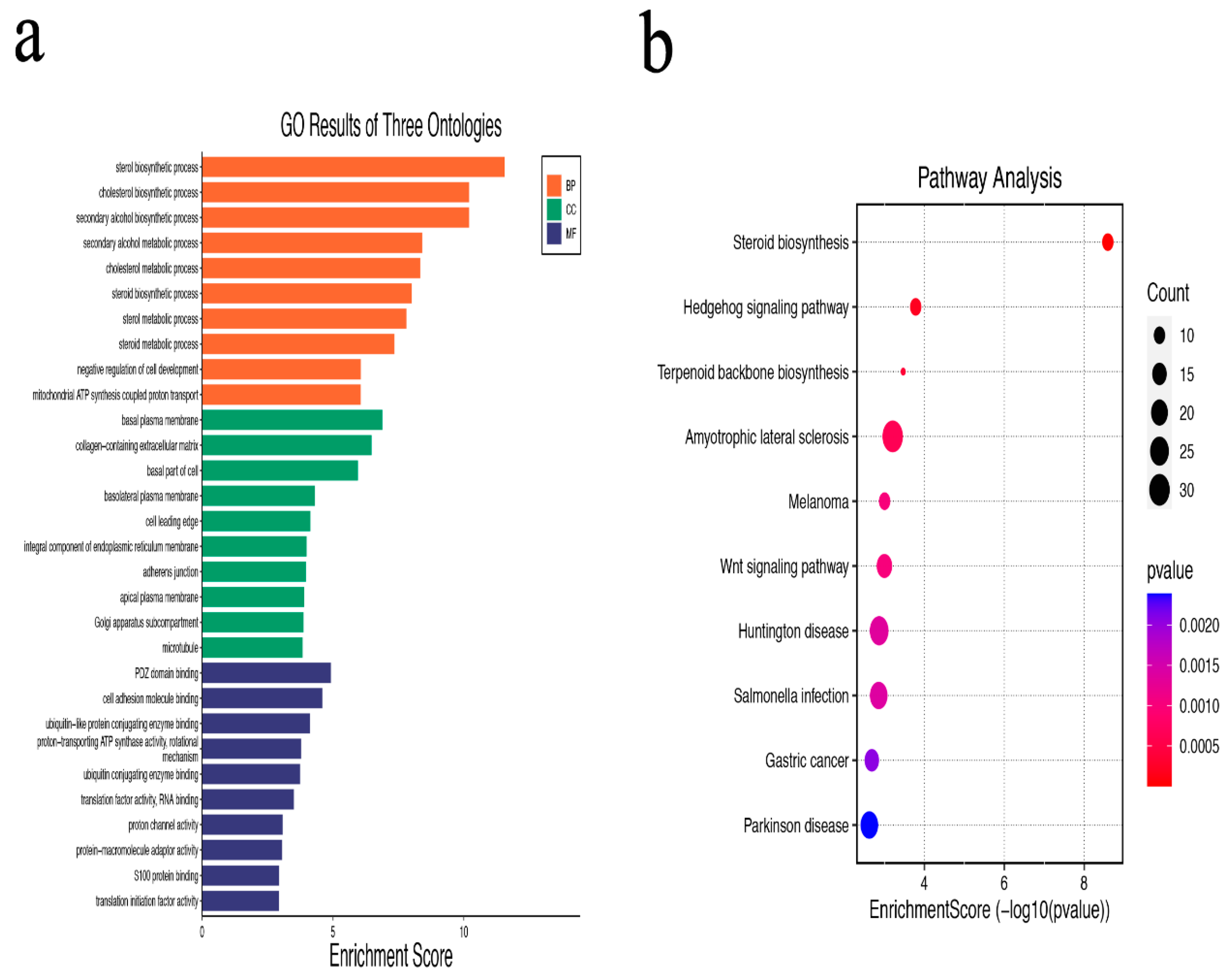
3.2. GO and KEGG Pathway Enrichment Analysis of DEGs
3.3. PPI Network Construction and Core Genes Analysis
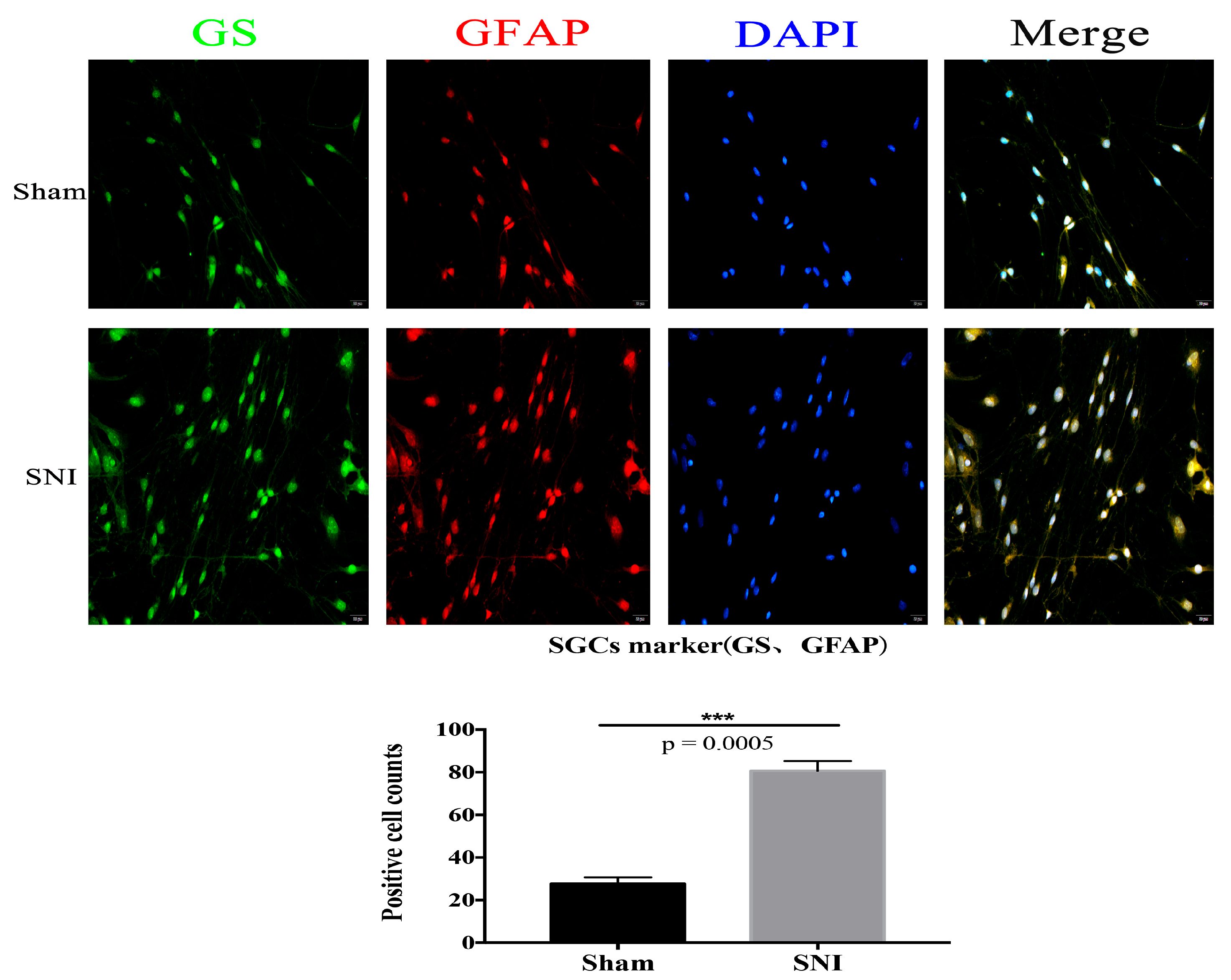
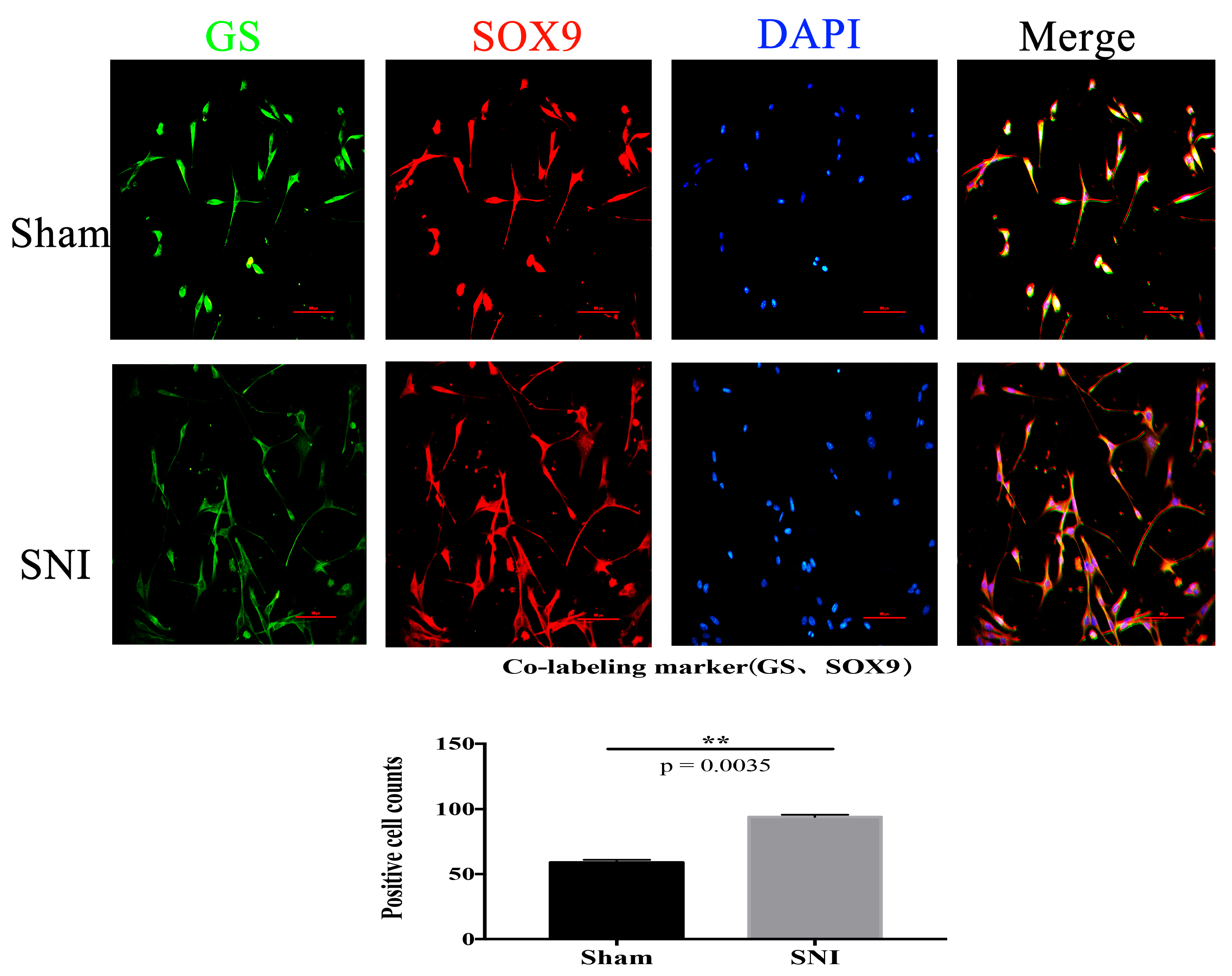
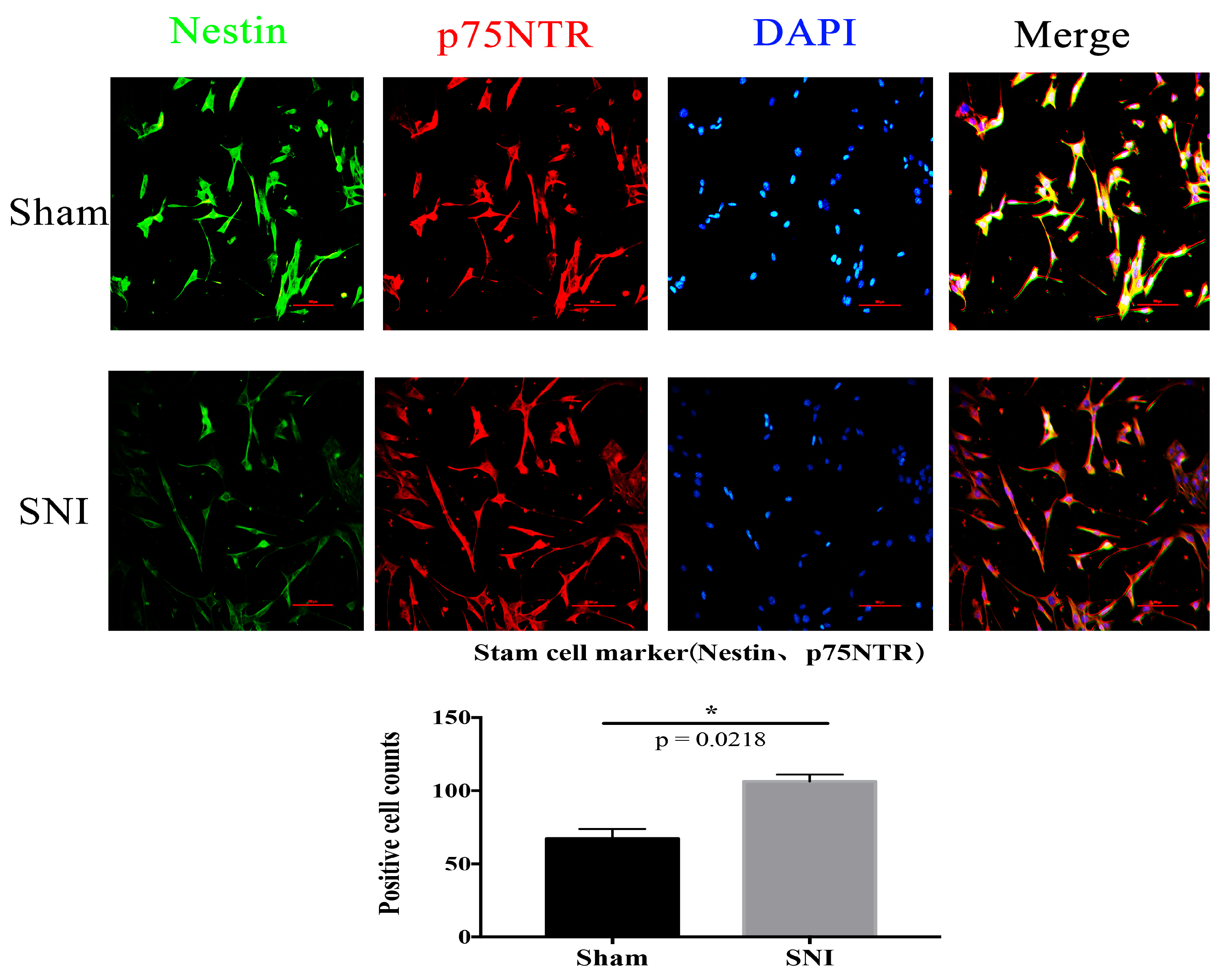
3.4. SNI Model and SGCs Culture
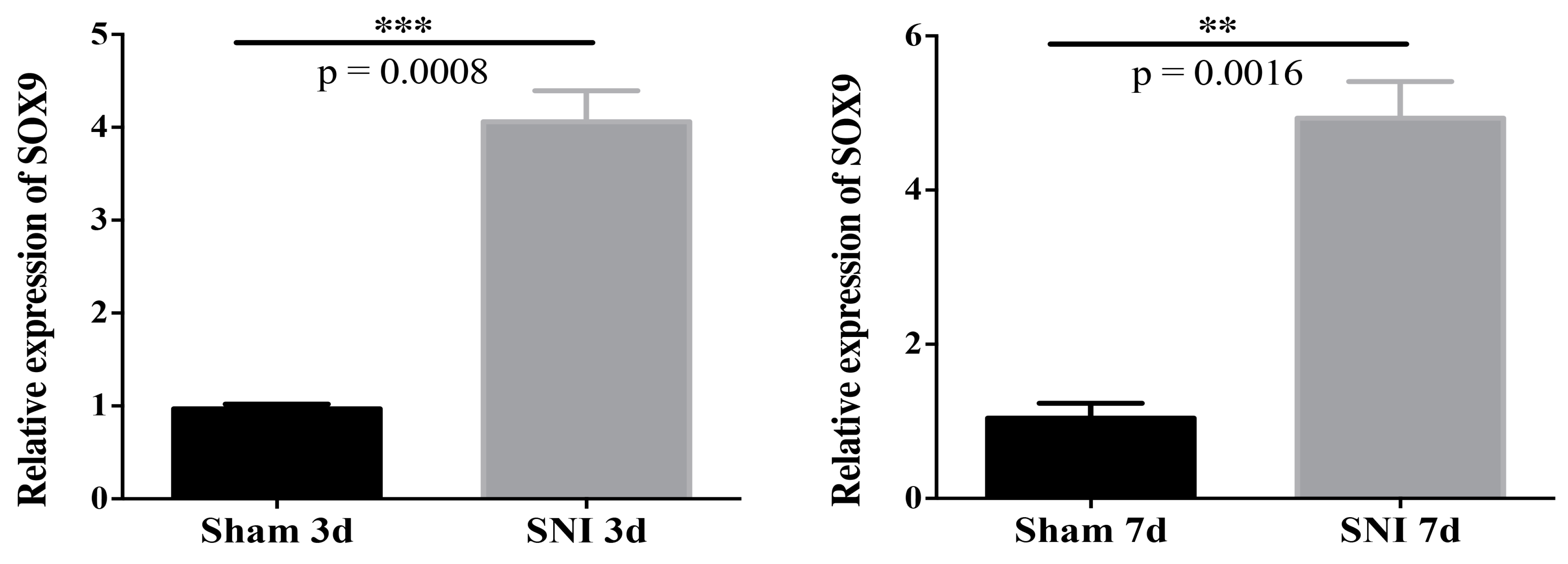
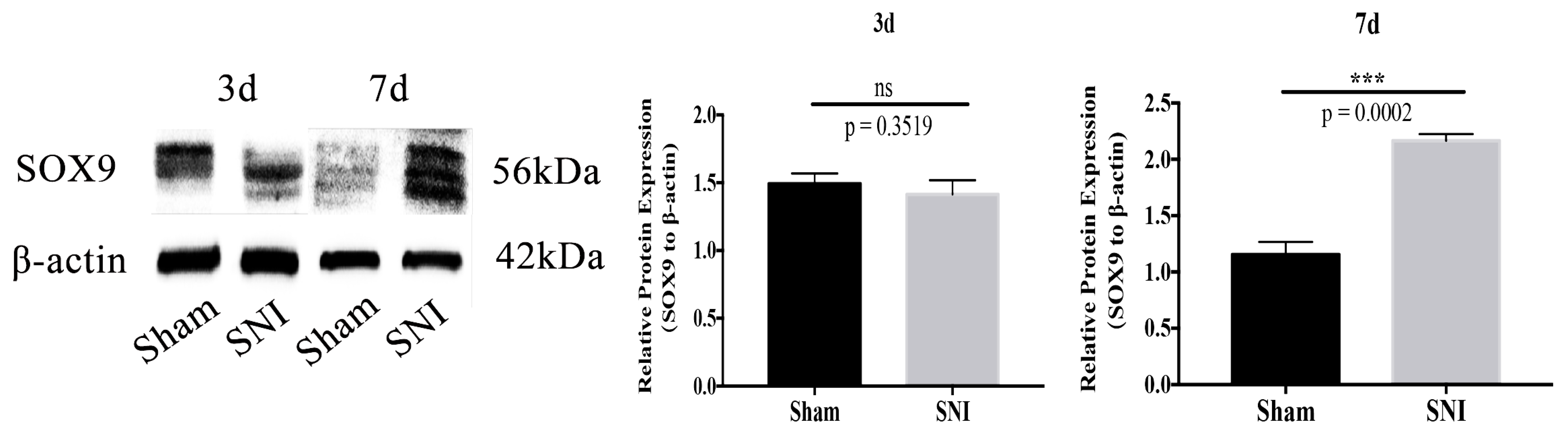
3.5. Sox9 mRNA and Protein Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CNS | Central Nervous System |
| DEGs | Differentially Expressed Genes |
| DRG | Dorsal Root Ganglion |
| ECM | Extracellular Matrix |
| GFAP | Glial Fibrillary Acidic Protein |
| GEO | Gene Expression Omnibus |
| GO | Gene Ontology |
| GS | Glutamine Synthetase |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LR | Lumbar Radiculopathy |
| Mmps | Matrix Metalloproteinases |
| NFH | Neurofilament Heavychain |
| PBS | Phosphate Buffered Saline |
| PNS | Peripheral Nervous System |
| PPI | Protein–Protein Interaction |
| p75NTR | p75 Neurotrophic Factor Receptor |
| SCI | Spinal Cord Injury |
| SCs | Schwann Cells |
| SNI | Sciatic Nerve Injury |
| SGCs | Satellite Glial Cells |
| Sox9 | SRY-box9 |
| STRING | Search Tool for the Retrieval of Interacting Genes/Proteins |
| TFs | Transcription Factors |
References
- Divac, N.; Aksić, M.; Rasulić, L.; Jakovčevski, M.; Basailović, M.; Jakovčevski, I. Pharmacology of repair after peripheral nerve injury. Int. J. Clin. Pharmacol. Ther. 2021, 59, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, D.; Hurtado, M.L.; De Gregorio, C.; Oñate, M.; Martínez, G.; Catenaccio, A.; Wishart, T.M.; Court, F.A. Collateral Sprouting of Peripheral Sensory Neurons Exhibits a Unique Transcriptomic Profile. Mol. Neurobiol. 2020, 57, 4232–4249. [Google Scholar] [CrossRef] [PubMed]
- Zambusi, A.; Ninkovic, J. Regeneration of the central nervous system-principles from brain regeneration in adult zebrafish. World J. Stem Cells 2020, 12, 8–24. [Google Scholar] [CrossRef]
- Liu, K.; Ma, W.; Li, C.; Li, J.; Zhang, X.; Liu, J.; Liu, W.; Wu, Z.; Zang, C.; Liang, Y.; et al. Advances in transcription factors related to neuroglial cell reprogramming. Transl. Neurosci. 2020, 11, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.J.; John, J.S.; Clarke, M.; Wright, A.; Ekberg, J. The Glia Response after Peripheral Nerve Injury: A Comparison between Schwann Cells and Olfactory Ensheathing Cells and Their Uses for Neural Regenerative Therapies. Int. J. Mol. Sci. 2017, 18, 287. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wang, W. Astrocyte transplantation for repairing the injured spinal cord. J. Biomed. Res. 2022, 36, 312–320. [Google Scholar] [CrossRef]
- Chen, G.; Liu, M.-H.; Li, W.; Zheng, J.-J.; Xu, Y.-G. Differential neuronal reprogramming induced by NeuroD1 from astrocytes in grey matter versus white matter. Neural Regen. Res. 2020, 15, 342–351. [Google Scholar] [CrossRef]
- Wang, F.; Cheng, L.; Zhang, X. Reprogramming Glial Cells into Functional Neurons for Neuro-regeneration: Challenges and Promise. Neurosci. Bull. 2021, 37, 1625–1636. [Google Scholar] [CrossRef]
- Chen, W.-H.; Lin, Y.-X.; Lin, L.; Zhang, B.-Q.; Xu, S.-X.; Wang, W. Identification of potential candidate proteins for reprogramming spinal cord-derived astrocytes into neurons: A proteomic analysis. Neural Regen. Res. 2021, 16, 2257–2263. [Google Scholar] [CrossRef]
- Nocera, G.; Jacob, C. Mechanisms of Schwann cell plasticity involved in peripheral nerve repair after injury. Cell. Mol. Life Sci. 2020, 77, 3977–3989. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Yang, C.; Liu, H.; Tang, C.; Huang, S. Effect of Schwann cell transplantation combined with electroacupuncture on axonal regeneration and remyelination in rats with spinal cord injury. Anat. Rec. 2021, 304, 2506–2520. [Google Scholar] [CrossRef]
- Jessen, K.R.; Mirsky, R. The repair Schwann cell and its function in regenerating nerves. J. Physiol. 2016, 594, 3521–3531. [Google Scholar] [CrossRef]
- Nadeau, J.R.; Wilson-Gerwing, T.D.; Verge, V.M. Induction of a reactive state in perineuronal satellite glial cells akin to that produced by nerve injury is linked to the level of p75NTR expression in adult sensory neurons. Glia 2014, 62, 763–777. [Google Scholar] [CrossRef]
- Hanani, M.; Verkhratsky, A. Satellite Glial Cells and Astrocytes, a Comparative Review. Neurochem. Res. 2021, 46, 2525–2537. [Google Scholar] [CrossRef] [PubMed]
- Masserdotti, G.; Gillotin, S.; Sutor, B.; Drechsel, D.; Irmler, M.; Jørgensen, H.F.; Sass, S.; Theis, F.J.; Beckers, J.; Berninger, B.; et al. Transcriptional Mechanisms of Proneural Factors and REST in Regulating Neuronal Reprogramming of Astrocytes. Cell Stem Cell 2015, 17, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Jager, S.E.; Pallesen, L.T.; Richner, M.; Harley, P.; Hore, Z.; McMahon, S.; Denk, F.; Vægter, C.B. Changes in the transcriptional fingerprint of satellite glial cells following peripheral nerve injury. Glia 2020, 68, 1375–1395. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zeng, L.; Liu, Y.; Long, Z.; Li, Y.; Wu, Y. Ultramicro changes of pathology and motor func-tion of sciatic nerve after transverse injury in rats. Chin. J. Clin. Rehabil. 2003, 7, 2396–2397. (In Chinese) [Google Scholar] [CrossRef]
- Guo, J.-H.; Li, L.-Y.; Wang, X.-B.; Ma, W.; Luo, T.; Yang, J.-W.; Wang, X.-P.; Dai, Y.-F. A novel primary culture method for high-purity satellite glial cells derived from rat dorsal root ganglion. Neural Regen. Res. 2019, 14, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Guy, R.; Grynspan, F.; Ben-Zur, T.; Panski, A.; Lamdan, R.; Danon, U.; Yaffe, D.; Offen, D. Human Muscle Progenitor Cells Overexpressing Neurotrophic Factors Improve Neuronal Regeneration in a Sciatic Nerve Injury Mouse Model. Front. Neurosci. 2019, 13, 151. [Google Scholar] [CrossRef]
- Abdollahifar, M.-A.; Farahni, R.M.; Rashidiani-Rashidabadi, A.; Heidari, M.H.; Sajadi, E.; Hejazi, F.; Fathabady, F.F.; Sadeghi, Y.; Aliaghaei, A.; Raoofi, A. Sciatic nerve injury alters the spatial arrangement of neurons and glial cells in the anterior horn of the spinal cord. Neural Regen. Res. 2019, 14, 1833–1840. [Google Scholar] [CrossRef]
- Yao, C.; Qin, J.; Wu, J.-C.; Wang, Q.-H.; Zhou, S.-L.; Mao, S.-S. Transcription factor networks involved in cell death in the dorsal root ganglia following peripheral nerve injury. Neural Regen. Res. 2018, 13, 1622–1627. [Google Scholar] [CrossRef] [PubMed]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The Human Transcription Factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.E.; Harrison, B.J.; Rau, K.K.; Hougland, M.T.; Bunge, M.B.; Mendell, L.M.; Petruska, J.C. Skin Incision Induces Expression of Axonal Regeneration-Related Genes in Adult Rat Spinal Sensory Neurons. J. Pain 2010, 11, 1066–1073. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-H.; Yu, Y.-W.; Chuang, Y.-C.; Lee, C.-H.; Chen, C.-C. ATF3-Expressing Large-Diameter Sensory Afferents at Acute Stage as Bio-Signatures of Persistent Pain Associated with Lumbar Radiculopathy. Cells 2021, 10, 992. [Google Scholar] [CrossRef] [PubMed]
- Theodore, L.N.; Hagedorn, E.J.; Cortes, M.; Natsuhara, K.; Liu, S.Y.; Perlin, J.R.; Yang, S.; Daily, M.L.; Zon, L.I.; North, T.E. Distinct Roles for Matrix Metalloproteinases 2 and 9 in Embryonic Hematopoietic Stem Cell Emergence, Migration, and Niche Colonization. Stem Cell Rep. 2017, 8, 1226–1241. [Google Scholar] [CrossRef] [PubMed]
- Inui, T.; Hoffer, M.; Balaban, C.D. Mild blast wave exposure produces intensity-dependent changes in MMP2 expression patches in rat brains—Findings from different blast severities. Brain Res. 2021, 1767, 147541. [Google Scholar] [CrossRef] [PubMed]
- Goyal, S.; Gupta, S. Rachana RNA Interference in Treatment of Diabetic Neuropathy. Indo Glob. J. Pharm. Sci. 2014, 4, 154. [Google Scholar] [CrossRef]
- Sarkar, A.; Hochedlinger, K. The Sox Family of Transcription Factors: Versatile Regulators of Stem and Progenitor Cell Fate. Cell Stem Cell 2013, 12, 15–30. [Google Scholar] [CrossRef]
- Mwangi, S.M.; Usta, Y.; Raja, S.M.; Anitha, M.; Chandrasekharan, B.; Parsadanian, A.; Sitaraman, S.V.; Srinivasan, S. Glial cell line-derived neurotrophic factor enhances neurogenin3 gene expression and β-cell proliferation in the developing mouse pancreas. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G283–G292. [Google Scholar] [CrossRef]
- Vong, K.I.; Leung, C.K.Y.; Behringer, R.R.; Kwan, K.M. Sox9 is critical for suppression of neurogenesis but not initiation of gliogenesis in the cerebellum. Mol. Brain 2015, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Qiu, B.; de Vries, R.J.; Caiazzo, M. Direct Cell Reprogramming of Mouse Fibroblasts into Functional Astrocytes Using Lentiviral Overexpression of the Transcription Factors NFIA, NFIB, and SOX9. Methods Mol. Biol. 2021, 2352, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Y.; Say, E.H.M.; Zhou, X.-F. Isolation and Characterization of Neural Crest Progenitors from Adult Dorsal Root Ganglia. Stem Cells 2007, 25, 2053–2065. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Yasaka, T.; Takano, M.; Ishihara, K. Neuronal and glial expression of inward rectifier potassium channel subunits Kir2.x in rat dorsal root ganglion and spinal cord. Neurosci. Lett. 2016, 617, 59–65. [Google Scholar] [CrossRef]
- Vogel, J.K.; Weider, M.; Engler, L.A.; Hillgärtner, S.; Schmitt, C.; Hermans-Borgmeyer, I.; Wegner, M. Sox9 overexpression exerts multiple stage-dependent effects on mouse spinal cord development. Glia 2020, 68, 932–946. [Google Scholar] [CrossRef]
- Zhang, H.; Xue, W.; Xue, X.; Fan, Y.; Yang, Y.; Zhao, Y.; Chen, B.; Yin, Y.; Yang, B.; Xiao, Z.; et al. Spatiotemporal dynamic changes, proliferation, and differentiation characteristics of Sox9-positive cells after severe complete transection spinal cord injury. Exp. Neurol. 2021, 337, 113556. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Ge, X.; Chen, X.; Xu, Y.; Yuan, W.-E.; Ouyang, Y. Enhancement of sciatic nerve regeneration with dual delivery of vascular endothelial growth factor and nerve growth factor genes. J. Nanobiotechnol. 2020, 18, 46. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Li, M.; Shen, D.; Ding, F.; Lu, S.; Zhao, Q.; Gu, X. Gene expression profiling of the rat sciatic nerve in early Wallerian degeneration after injury☆. Neural Regen. Res. 2012, 7, 1285–1292. [Google Scholar] [CrossRef] [PubMed]
- Ferdowsi, S.; Abdolmaleki, A.; Asadi, A.; Zahri, S. Effect of Azithromycin on Sciatic Nerve Injury in the Wistar Rats. Neurochem. Res. 2022, 48, 161–171. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Description | p-Value | Gene ID |
|---|---|---|---|
| mmu00100 | Steroid biosynthesis | 0.00000000256 | Sc5d/Dhcr24/Hsd17b7/Tm7sf2/Lss/Dhcr7/Msmo1/Sqle/Nsdhl/Cyp51 |
| mmu04340 | Hedgehog signaling pathway | 0.000165308 | Kif3a/Btrc/Bcl2/Smurf2/Prkacb/Cdon/Ccnd1/Kif7/Spop/Csnk1d |
| mmu00900 | Terpenoid backbone biosynthesis | 0.000339229 | Hmgcr/Hmgcs1/Fdps/Acat2/Pcyox1/Ggps1 |
| mmu05014 | Amyotrophic lateral sclerosis | 0.000622229 | Atp5h/Atg14/Chchd10/Atp5b/Atp5j/Atp5c1/Uqcr11/Bcl2/Atp5o/Pik3r4/Wipi2/Optn/Cox6c/Cox7a1/Tpr/Nup107/Nxt2/Cyc1/Atxn2l/Tuba1c/Tnfrsf1a/Xbp1/Seh1l/Tubb2b/Tubb6/Hspa5/Map1lc3a/Dctn2/Tuba1a/Bax |
| mmu05218 | Melanoma | 0.000991453 | Cdh1/Igf1/Hgf/E2f1/Fgf1/Ccnd1/Gadd45a/Cdkn1a/Bax/Fgf3 |
| mmu04310 | Wnt signaling pathway | 0.001001666 | Ctnnd2/Frzb/Daam2/Map3k7/Btrc/Dvl2/Fzd3/Fzd1/AF366264/Prkacb/Ccnd1/Vangl1/Cxxc4/Siah1b/Tle6/Rnf43/Fzd6 |
| mmu05016 | Huntington disease | 0.001358182 | Atp5h/Ppargc1a/Atg14/Atp5b/Atp5j/Atp5c1/Uqcr11/Slc1a3/Atp5o/Pik3r4/Kcnj10/Wipi2/Sp1/Cox6c/Cox7a1/Cyc1/Tuba1c/Vdac3/Tubb2b/Tubb6/Itpr1/Dctn2/Tuba1a/Cltb/Bax |
| mmu05132 | Salmonella infection | 0.001392876 | Dynlt3/Map3k7/Vps33a/Bcl2/Wasf3/Rab7b/Dynll2/Tuba1c/Tnfrsf1a/Rhog/Myl12b/Tubb2b/Tubb6/Pik3c2b/Podxl/Dctn2/Casp4/Tuba1a/Nfkbia/Rps3/Casp7/Bax |
| mmu05226 | Gastric cancer | 0.00207011 | Cdh1/Hgf/E2f1/Dvl2/Fgf1/Fzd3/Bcl2/Fzd1/Ccnd1/Gadd45a/Abcb1a/Cdkn1a/Fzd6/Bax/Fgf3 |
| mmu05012 | Parkinson disease | 0.002389542 | Atp5h/Atp5b/Keap1/Atp5j/Atp5c1/Uqcr11/Atp5o/Prkacb/Cox6c/Cox7a1/Mfn1/Cyc1/Tuba1c/Vdac3/Maoa/Xbp1/Tubb2b/Tubb6/Hspa5/Itpr1/Tuba1a/Bax |
| Different Algorithms | ||||
|---|---|---|---|---|
| Bottleneck | EPC | Closeness | Readiality | |
| Top 10 core genes | Ank3 ↑ | Smurf2 ↓ | Ccl2 ↑ | Ccl2 ↑ |
| Cenpf ↑ | Ace ↑ | Ctgf ↑ | Cenpf ↑ | |
| Atf3 ↑ | Ccl2 ↑ | Hgf ↓ | Ctgf ↑ | |
| Gngt2 ↑ | Ctgf ↑ | Atf3 ↑ | Atf3 ↑ | |
| Rab11fip5 ↑ | Hgf ↓ | Plau ↑ | Plau ↑ | |
| Kif22 ↑ | Atf3 ↑ | Cdkn1a ↑ | Cdkn1a ↑ | |
| Kif3a ↓ | Cdkn1a ↑ | Mmp2 ↑ | Mmp2 ↑ | |
| Mmp2 ↑ | Kif22 ↑ | Igf1 ↓ | Igf1 ↓ | |
| Igf1 ↓ | Mmp2 ↑ | Gm3839 ↑ | Sox9 ↑ | |
| Sox9 ↑ | Sox9 ↑ | Sox9 ↑ | Cks2 ↑ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; Ma, W.; Yang, J.; Liu, W.; Zhang, S.; Zhu, K.; Liu, J.; Xiang, X.; Wang, G.; Wu, H.; et al. Integrative Analysis Reveals the Expression Pattern of SOX9 in Satellite Glial Cells after Sciatic Nerve Injury. Brain Sci. 2023, 13, 281. https://doi.org/10.3390/brainsci13020281
Liu K, Ma W, Yang J, Liu W, Zhang S, Zhu K, Liu J, Xiang X, Wang G, Wu H, et al. Integrative Analysis Reveals the Expression Pattern of SOX9 in Satellite Glial Cells after Sciatic Nerve Injury. Brain Sciences. 2023; 13(2):281. https://doi.org/10.3390/brainsci13020281
Chicago/Turabian StyleLiu, Kuangpin, Wei Ma, Jinwei Yang, Wei Liu, Sijia Zhang, Kewei Zhu, Jie Liu, Xianglin Xiang, Guodong Wang, Hongjie Wu, and et al. 2023. "Integrative Analysis Reveals the Expression Pattern of SOX9 in Satellite Glial Cells after Sciatic Nerve Injury" Brain Sciences 13, no. 2: 281. https://doi.org/10.3390/brainsci13020281