
Could Vitamins Have a Positive Impact on the Treatment of Parkinson’s Disease?

Abstract
:1. Introduction
2. Oxidative Stress and Neuroinflammation at the Core of PD Pathogenesis
3. Role of Vitamins in the Pathogenesis of PD
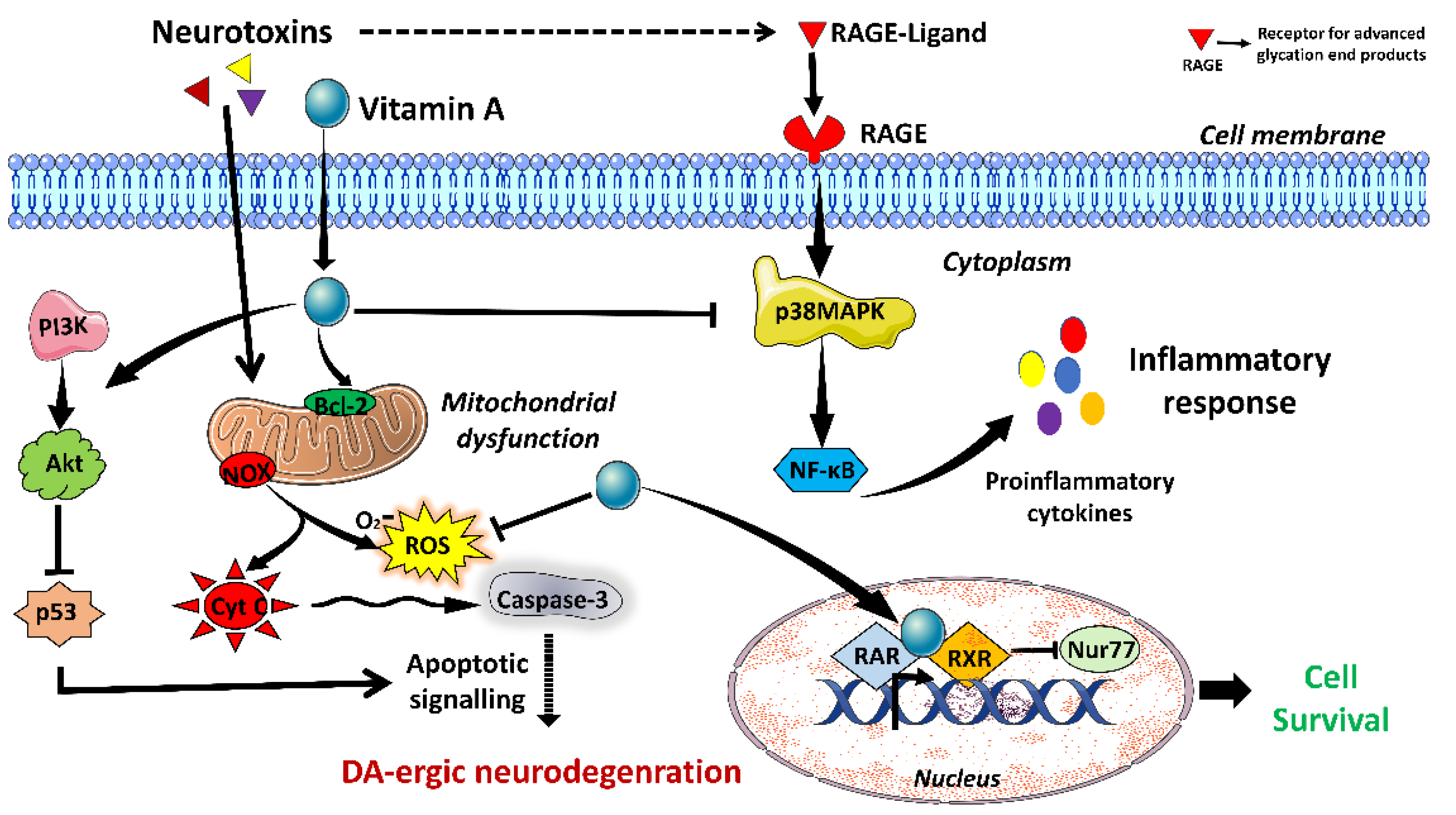
3.1. Vitamin A
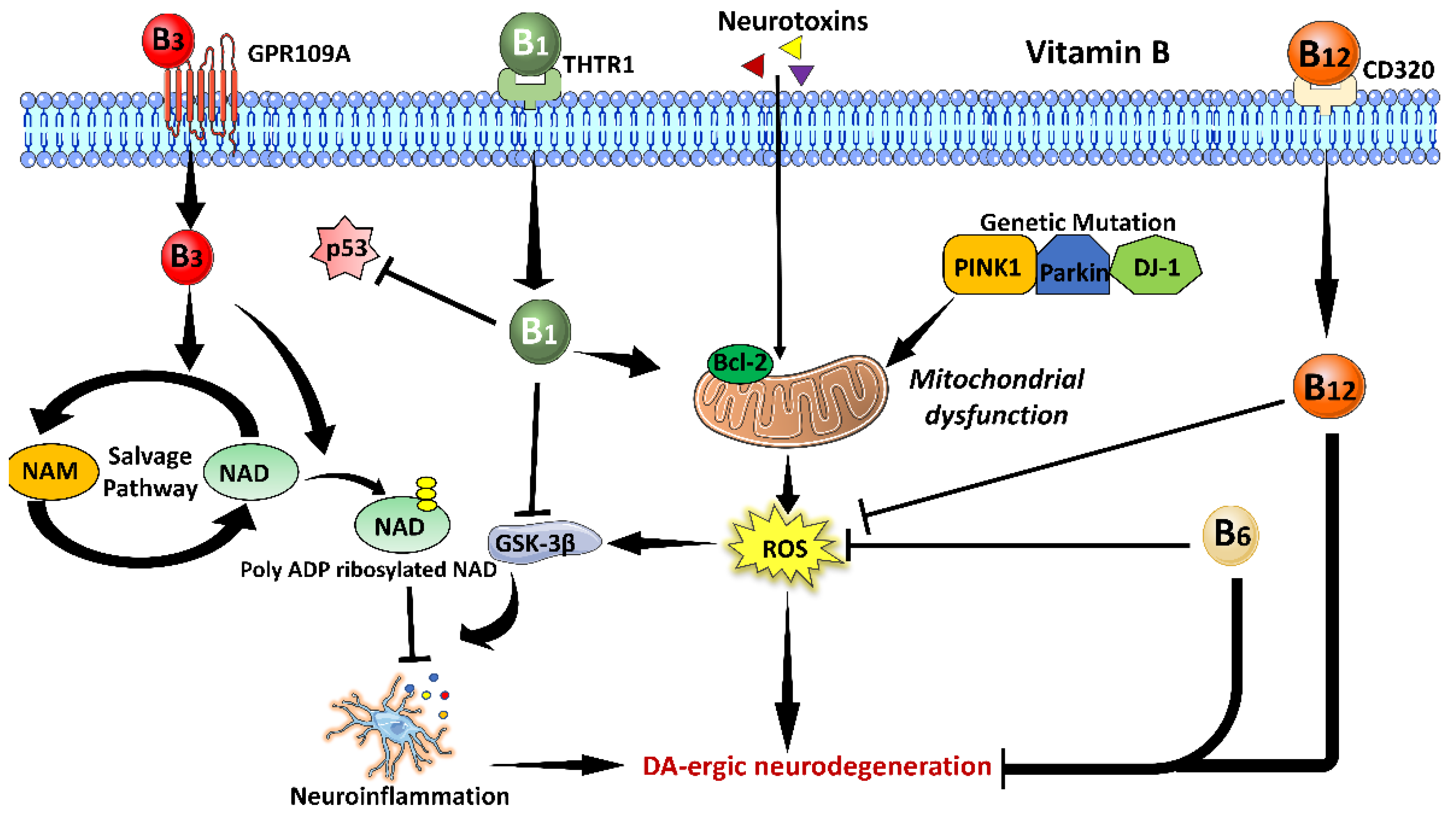
3.2. Vitamin B Family
3.2.1. Vitamin B1
3.2.2. Vitamin B3
3.2.3. Vitamin B6
3.2.4. Vitamin B12
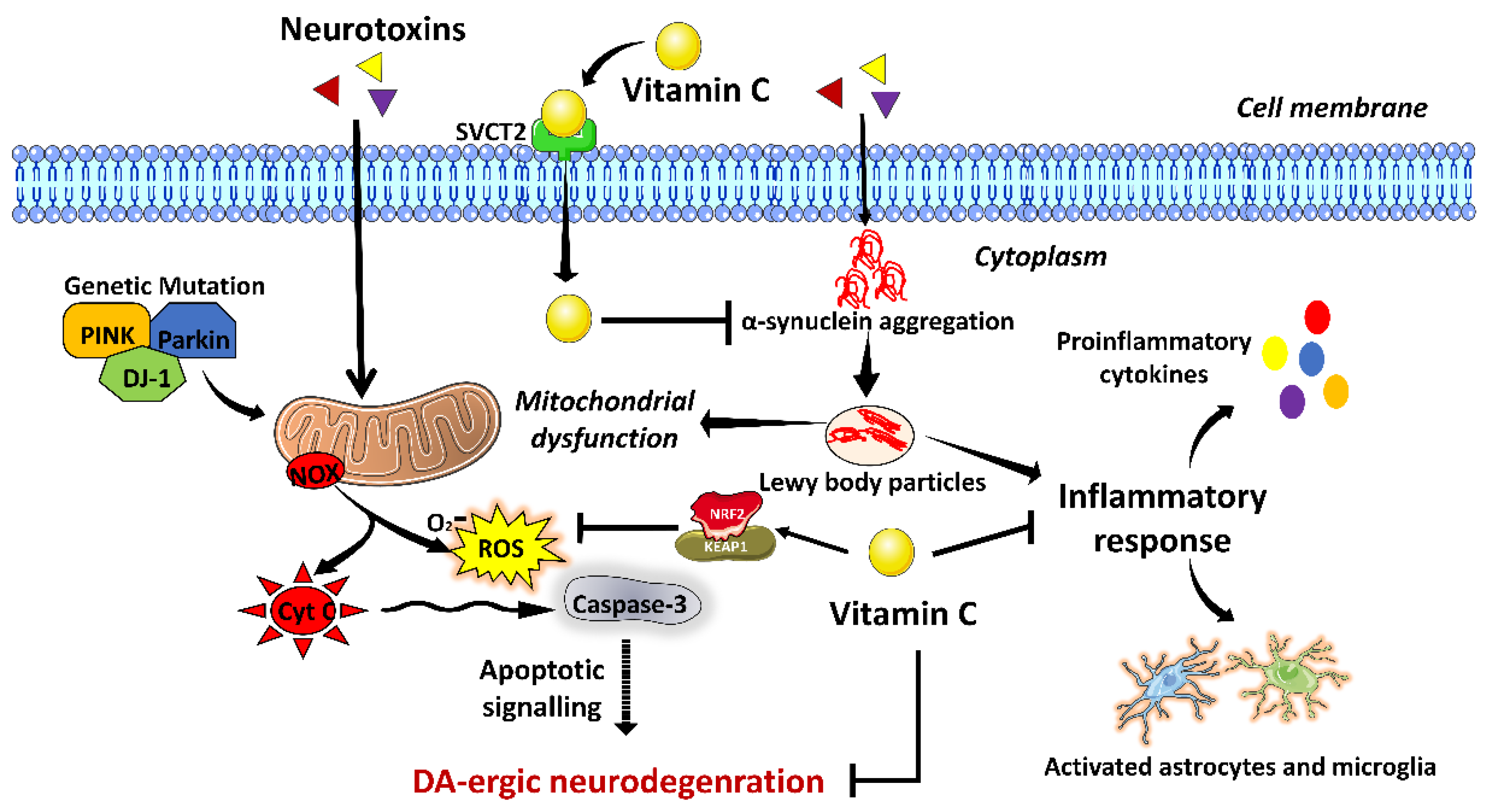
3.3. Vitamin C
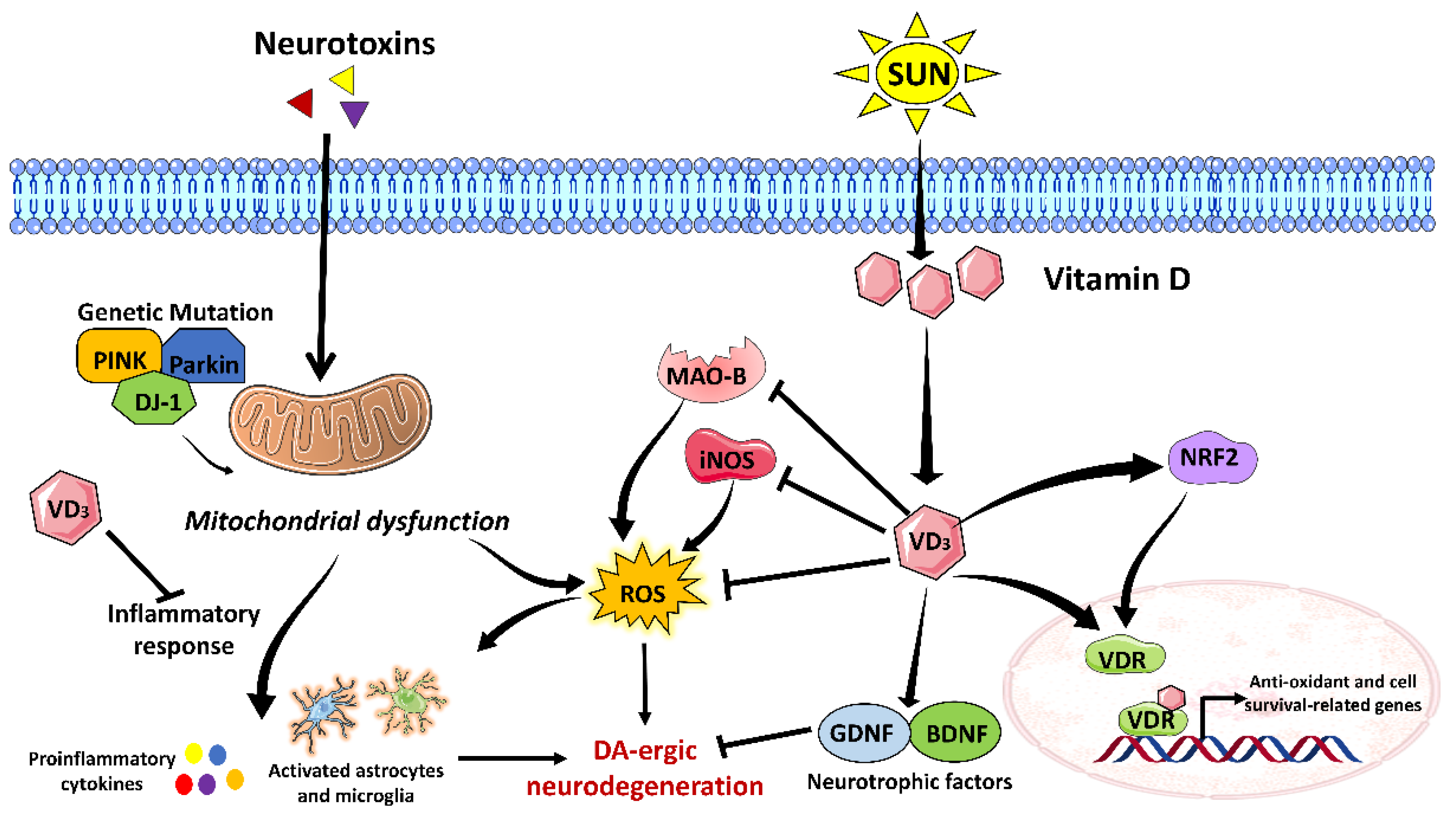
3.4. Vitamin D
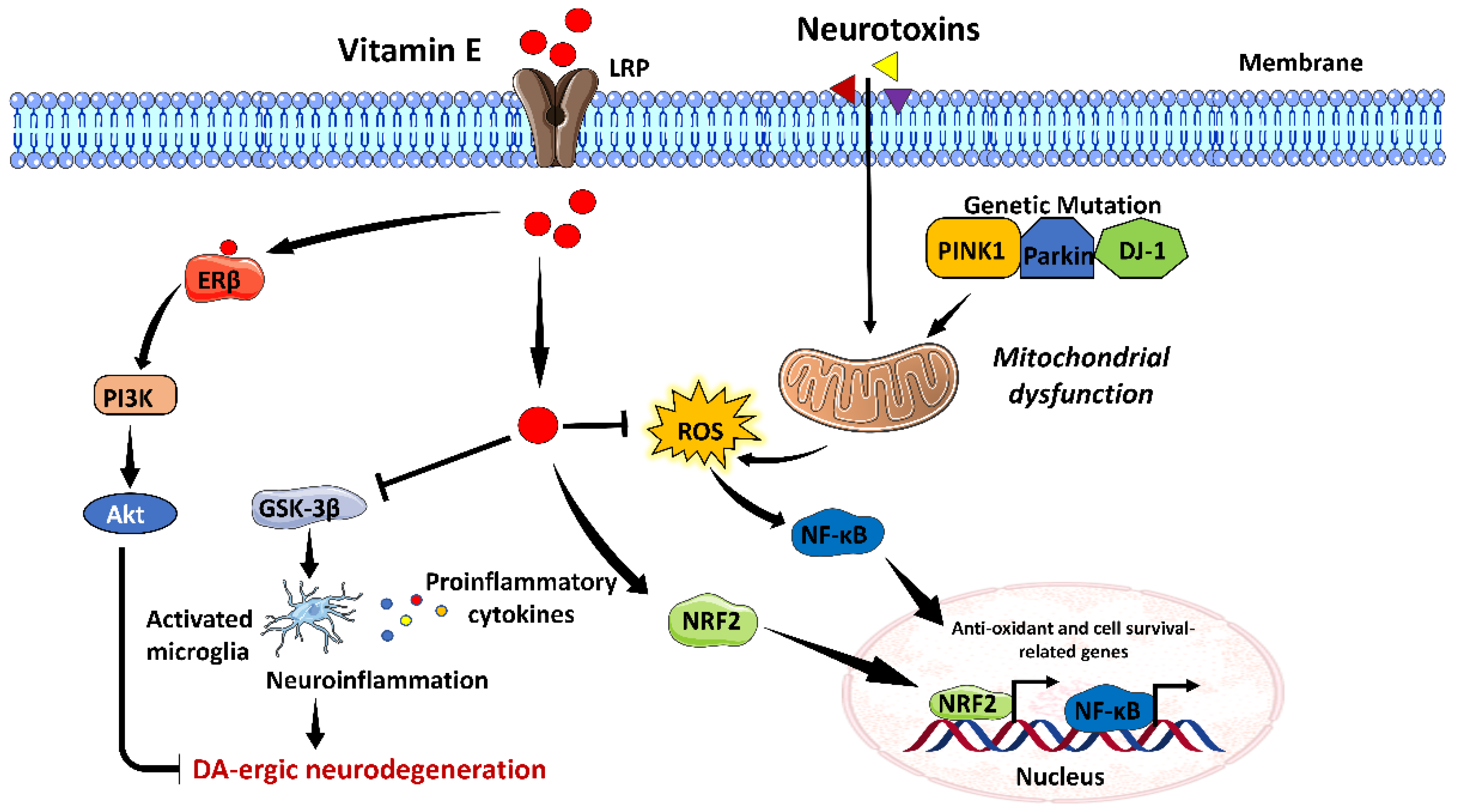
3.5. Vitamin E
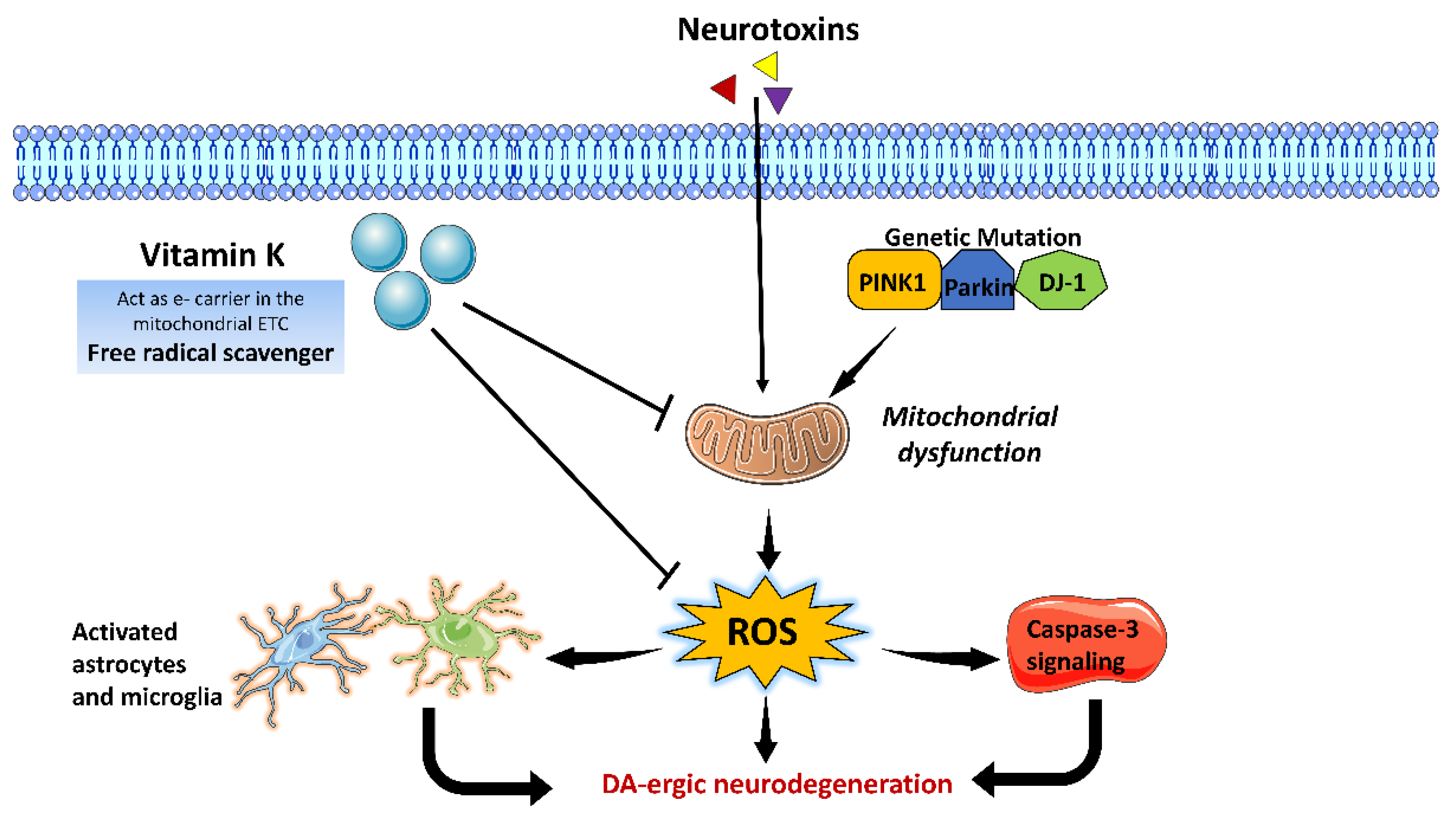
3.6. Vitamin K
4. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kunzler, A.; Kolling, E.A.; da Silva, J.D., Jr.; Gasparotto, J.; de Bittencourt Pasquali, M.A.; Moreira, J.C.F.; Gelain, D.P. Retinol (vitamin A) increases α-synuclein, β-amyloid peptide, tau phosphorylation and RAGE content in human SH-SY5Y neuronal cell line. Neurochem. Res. 2017, 42, 2788–2797. [Google Scholar] [CrossRef]
- Man Anh, H.; Linh, D.M.; My Dung, V.; Thi Phuong Thao, D. Evaluating dose-and time-dependent effects of vitamin c treatment on a parkinson’s disease fly model. Park. Dis. 2019, 2019, 9720546. [Google Scholar] [CrossRef]
- Kunzler, A.; Ribeiro, C.T.; Gasparotto, J.; Petiz, L.L.; da Rosa Silva, H.T.; da Silva, J.D., Jr.; Bortolin, R.; de Souza, P.O.; Barreto, F.; Espitia-Perez, P. The effects of retinol oral supplementation in 6-hydroxydopamine dopaminergic denervation model in Wistar rats. Neurochem. Int. 2019, 125, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Giri, B.; Belanger, K.; Seamon, M.; Bradley, E.; Purohit, S.; Chong, R.; Morgan, J.C.; Baban, B.; Wakade, C. Niacin ameliorates neuro-inflammation in Parkinson’s disease via GPR109A. Int. J. Mol. Sci. 2019, 20, 4559. [Google Scholar] [CrossRef]
- Alster, P.; Madetko, N.; Friedman, A. Neutrophil-to-lymphocyte ratio (NLR) at boundaries of Progressive Supranuclear Palsy Syndrome (PSPS) and Corticobasal Syndrome (CBS). Neurol. I Neurochir. Pol. 2021, 55, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Madetko, N.; Migda, B.; Alster, P.; Turski, P.; Koziorowski, D.; Friedman, A. Platelet-to-lymphocyte ratio and neutrophil-tolymphocyte ratio may reflect differences in PD and MSA-P neuroinflammation patterns. Neurol. I Neurochir. Pol. 2022, 56, 148–155. [Google Scholar] [CrossRef]
- Ganguly, J.; Bernaola, M.T.; Jog, M. Role of Vitamins in Advanced therapy for Parkinson’s disease: Decoding the paradox. Can. J. Neurol. Sci. 2022, 49, 3–4. [Google Scholar] [CrossRef]
- Barmaki, H.; Morovati, A.; Eydivandi, Z.; Naleshkenani, F.J.; Saedi, S.; Musavi, H.; Abbasi, M.; Hemmati-Dinarvand, M. The association between serum oxidative stress indexes and pathogenesis of Parkinson’s disease in the northwest of Iran. Iran. J. Public Health 2021, 50, 606. [Google Scholar] [CrossRef] [PubMed]
- Karahalil, B.; Miser Salihoğlu, E.; Elkama, A.; Orhan, G.; Saygın, E.; Yardim Akaydin, S. Individual susceptibility has a major impact on strong association between oxidative stress, defence systems and Parkinson’s disease. Basic Clin. Pharmacol. Toxicol. 2022, 130, 158–170. [Google Scholar] [CrossRef]
- Chia, S.J.; Tan, E.-K.; Chao, Y.-X. Historical perspective: Models of Parkinson’s disease. Int. J. Mol. Sci. 2020, 21, 2464. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.-H.; Chen, C.-M. The role of oxidative stress in Parkinson’s disease. Antioxidants 2020, 9, 597. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Vondrakova, D.; Lawson, M.; Valko, M. Metals, oxidative stress and neurodegenerative disorders. Mol. Cell. Biochem. 2010, 345, 91–104. [Google Scholar] [CrossRef]
- Jenner, P.; Dexter, D.; Sian, J.; Schapira, A.; Marsden, C. Oxidative stress as a cause of nigral cell death in Parkinson’s disease and incidental Lewy body disease. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 1992, 32, S82–S87. [Google Scholar] [CrossRef]
- Ciulla, M.; Marinelli, L.; Cacciatore, I.; Stefano, A.D. Role of dietary supplements in the management of Parkinson’s disease. Biomolecules 2019, 9, 271. [Google Scholar] [CrossRef] [PubMed]
- Kalampokini, S.; Becker, A.; Fassbender, K.; Lyros, E.; Unger, M.M. Nonpharmacological modulation of chronic inflammation in Parkinson’s disease: Role of diet interventions. Park. Dis. 2019, 2019, 7535472. [Google Scholar] [CrossRef] [PubMed]
- Yemula, N.; Dietrich, C.; Dostal, V.; Hornberger, M. Parkinson’s disease and the gut: Symptoms, nutrition, and microbiota. J. Park. Dis. 2021, 11, 1491–1505. [Google Scholar] [CrossRef]
- Al-Amin, M.; Bradford, D.; Sullivan, R.K.; Kurniawan, N.D.; Moon, Y.; Han, S.H.; Zalesky, A.; Burne, T.H. Vitamin D deficiency is associated with reduced hippocampal volume and disrupted structural connectivity in patients with mild cognitive impairment. Hum. Brain Mapp. 2019, 40, 394–406. [Google Scholar] [CrossRef]
- Allan Butterfield, D.; Castegna, A.; Drake, J.; Scapagnini, G.; Calabrese, V. Vitamin E and neurodegenerative disorders associated with oxidative stress. Nutr. Neurosci. 2002, 5, 229–239. [Google Scholar] [CrossRef]
- De Lau, L.; Koudstaal, P.; Witteman, J.; Hofman, A.; Breteler, M. Dietary folate, vitamin B12, and vitamin B6 and the risk of Parkinson disease. Neurology 2006, 67, 315–318. [Google Scholar] [CrossRef]
- Ferland, G. Vitamin K, an emerging nutrient in brain function. Biofactors 2012, 38, 151–157. [Google Scholar] [CrossRef]
- Takeda, A.; Nyssen, O.P.; Syed, A.; Jansen, E.; Bueno-de-Mesquita, B.; Gallo, V. Vitamin A and carotenoids and the risk of Parkinson’s disease: A systematic review and meta-analysis. Neuroepidemiology 2014, 42, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.-H.; Shen, H.; Diaz-Ruiz, O.; Bäckman, C.M.; Bae, E.; Yu, S.-J.; Wang, Y. Early post-treatment with 9-cis retinoic acid reduces neurodegeneration of dopaminergic neurons in a rat model of Parkinson’s disease. BMC Neurosci. 2012, 13, 120. [Google Scholar] [CrossRef] [PubMed]
- Gelain, D.P.; de Bittencourt Pasquali, M.A.; Caregnato, F.F.; Moreira, J.C.F. Vitamin A (retinol) up-regulates the receptor for advanced glycation endproducts (RAGE) through p38 and Akt oxidant-dependent activation. Toxicology 2011, 289, 38–44. [Google Scholar] [CrossRef]
- Jiménez-Jiménez, F.J.; Molina, J.; Fernández-Calle, P.; Vázquez, A.; Pondal, M.; del Ser, T.; Gómez-Pastor, A.; Codoceo, R. Serum levels of vitamin A in Parkinson’s disease. J. Neurol. Sci. 1992, 111, 73–76. [Google Scholar] [CrossRef]
- Ying, A.F.; Khan, S.; Wu, Y.; Jin, A.; Wong, A.S.; Tan, E.K.; Yuan, J.M.; Koh, W.P.; Tan, L.C. Dietary antioxidants and risk of Parkinson’s disease in the Singapore Chinese Health Study. Mov. Disord. 2020, 35, 1765–1773. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Ke, Z.; Luo, J. Thiamine deficiency and neurodegeneration: The interplay among oxidative stress, endoplasmic reticulum stress, and autophagy. Mol. Neurobiol. 2017, 54, 5440–5448. [Google Scholar] [CrossRef]
- Lương, K.v.; Nguyễn, L.T. The beneficial role of thiamine in Parkinson disease. CNS Neurosci. Ther. 2013, 19, 461–468. [Google Scholar] [CrossRef]
- Costantini, A.; Pala, M.I.; Compagnoni, L.; Colangeli, M. High-dose thiamine as initial treatment for Parkinson’s disease. Case Rep. 2013, 2013, bcr2013009289. [Google Scholar] [CrossRef]
- Håglin, L.; Johansson, I.; Forsgren, L.; Bäckman, L. Intake of vitamin B before onset of Parkinson’s disease and atypical parkinsonism and olfactory function at the time of diagnosis. Eur. J. Clin. Nutr. 2017, 71, 97–102. [Google Scholar] [CrossRef]
- Håglin, L.; Domellöf, M.; Bäckman, L.; Forsgren, L. Low plasma thiamine and phosphate in male patients with Parkinson’s disease is associated with mild cognitive impairment. Clin. Nutr. ESPEN 2020, 37, 93–99. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, M.; Li, C.; Jiang, X.; Su, Y.; Zhang, Y. Benefits of Vitamins in the Treatment of Parkinson’s Disease. Oxidative Med. Cell. Longev. 2019, 2019, 9426867. [Google Scholar] [CrossRef]
- Griffin, S.M.; Pickard, M.R.; Hawkins, C.P.; Williams, A.C.; Fricker, R.A. Nicotinamide restricts neural precursor proliferation to enhance catecholaminergic neuronal subtype differentiation from mouse embryonic stem cells. PLoS ONE 2020, 15, e0233477. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Li, X.; Gao, H.; Feng, Z.; Li, X.; Zhao, L.; Jia, X.; Zhang, H.; Liu, J. High doses of nicotinamide prevent oxidative mitochondrial dysfunction in a cellular model and improve motor deficit in a Drosophila model of Parkinson’s disease. J. Neurosci. Res. 2008, 86, 2083–2090. [Google Scholar] [CrossRef]
- Lehmann, S.; Loh, S.H.; Martins, L.M. Enhancing NAD+ salvage metabolism is neuroprotective in a PINK1 model of Parkinson’s disease. Biol. Open 2017, 6, 141–147. [Google Scholar] [CrossRef]
- Wakade, C.; Chong, R. A novel treatment target for Parkinson’s disease. J. Neurol. Sci. 2014, 347, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Motawi, T.K.; Sadik, N.A.; Hamed, M.A.; Ali, S.A.; Khalil, W.K.; Ahmed, Y.R. Potential therapeutic effects of antagonizing adenosine A2A receptor, curcumin and niacin in rotenone-induced Parkinson’s disease mice model. Mol. Cell. Biochem. 2020, 465, 89–102. [Google Scholar] [CrossRef]
- Wakade, C.; Giri, B.; Malik, A.; Khodadadi, H.; Morgan, J.C.; Chong, R.K.; Baban, B. Niacin modulates macrophage polarization in Parkinson’s disease. J. Neuroimmunol. 2018, 320, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Rojo-Sebastián, A.; González-Robles, C.; de Yébenes, J.G. Vitamin B6 deficiency in patients with Parkinson disease treated with levodopa/carbidopa. Clin. Neuropharmacol. 2020, 43, 151–157. [Google Scholar] [CrossRef]
- di Salvo, M.L.; Safo, M.K.; Contestabile, R. Biomedical aspects of pyridoxal 5’-phosphate availability. Front. Biosci. -Elite 2012, 4, 897–913. [Google Scholar]
- Elstner, M.; Morris, C.M.; Heim, K.; Lichtner, P.; Bender, A.; Mehta, D.; Schulte, C.; Sharma, M.; Hudson, G.; Goldwurm, S. Single-cell expression profiling of dopaminergic neurons combined with association analysis identifies pyridoxal kinase as Parkinson’s disease gene. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 2009, 66, 792–798. [Google Scholar] [CrossRef]
- Modica, J.S.; Bonno, D.; Lizarraga, K.J. Pearls and Oy-sters: Vitamin B6 deficiency presenting with new-onset epilepsy and status epilepticus in a patient with Parkinson disease. Neurology 2020, 94, e2605–e2607. [Google Scholar] [CrossRef]
- Murakami, K.; Miyake, Y.; Sasaki, S.; Tanaka, K.; Fukushima, W.; Kiyohara, C.; Tsuboi, Y.; Yamada, T.; Oeda, T.; Miki, T. Dietary intake of folate, vitamin B6, vitamin B12 and riboflavin and risk of Parkinson’s disease: A case–control study in Japan. Br. J. Nutr. 2010, 104, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Wang, Y.; Wei, W.; Zhao, W.; Lu, F.; Liu, F. Vitamin B12 inhibits α-synuclein fibrillogenesis and protects against amyloid-induced cytotoxicity. Food Funct. 2019, 10, 2861–2870. [Google Scholar] [CrossRef] [PubMed]
- Ozer, F.; Meral, H.; Hanoglu, L.; Aydemir, T.; Yilsen, M.; Cetin, S.; Ozturk, O.; Seval, H.; Koldas, M. Plasma homocysteine levels in patients treated with levodopa: Motor and cognitive associations. Neurol. Res. 2006, 28, 853–858. [Google Scholar] [CrossRef]
- McCarter, S.J.; Teigen, L.M.; McCarter, A.R.; Benarroch, E.E.; Louis, E.K.S.; Savica, R. Low vitamin B12 and Parkinson disease: Potential link to reduced cholinergic transmission and severity of disease. Mayo Clin. Proc. 2019, 94, 757–762. [Google Scholar] [CrossRef] [PubMed]
- McCarter, S.J.; Stang, C.; Turcano, P.; Mielke, M.M.; Ali, F.; Bower, J.H.; Savica, R. Higher vitamin B12 level at Parkinson’s disease diagnosis is associated with lower risk of future dementia. Park. Relat. Disord. 2020, 73, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Dietiker, C.; Kim, S.; Zhang, Y.; Christine, C.W.; Investigators, N.N.-P. Characterization of vitamin b12 supplementation and correlation with clinical outcomes in a large longitudinal study of early Parkinson’s disease. J. Mov. Disord. 2019, 12, 91. [Google Scholar] [CrossRef]
- Orozco-Barrios, C.E.; Battaglia-Hsu, S.-F.; Arango-Rodriguez, M.L.; Ayala-Davila, J.; Chery, C.; Alberto, J.-M.; Schroeder, H.; Daval, J.-L.; Martinez-Fong, D.; Gueant, J.-L. Vitamin B12-impaired metabolism produces apoptosis and Parkinson phenotype in rats expressing the transcobalamin-oleosin chimera in substantia nigra. PLoS ONE 2009, 4, e8268. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhao, Z.; Yang, N.; Xin, C.; Li, Z.; Xu, J.; Ma, B.; Lim, K.-L.; Li, L.; Wu, Q. Vitamin B12 Ameliorates the Pathological Phenotypes of Multiple Parkinson’s Disease Models by Alleviating Oxidative Stress. Antioxidants 2023, 12, 153. [Google Scholar] [CrossRef]
- Uddin, M.S.; Millat, M.; Baral, P.K.; Ferdous, M.; Uddin, M.; Sarwar, M.; Islam, M.S. The protective role of vitamin C in the management of COVID-19: A Review. J. Egypt. Public Health Assoc. 2021, 96, 33. [Google Scholar] [CrossRef]
- He, X.-B.; Kim, M.; Kim, S.-Y.; Yi, S.-H.; Rhee, Y.-H.; Kim, T.; Lee, E.-H.; Park, C.-H.; Dixit, S.; Harrison, F.E. Vitamin C facilitates dopamine neuron differentiation in fetal midbrain through TET1-and JMJD3-dependent epigenetic control manner. Stem Cells 2015, 33, 1320–1332. [Google Scholar] [CrossRef] [Green Version]
- Paraskevas, G.P.; Kapaki, E.; Petropoulou, O.; Anagnostouli, M.; Vagenas, V.; Papageorgiou, C. Plasma levels of antioxidant vitamins C and E are decreased in vascular parkinsonism. J. Neurol. Sci. 2003, 215, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Casani, S.; Gómez-Pastor, R.; Matallana, E.; Paricio, N. Antioxidant compound supplementation prevents oxidative damage in a Drosophila model of Parkinson’s disease. Free Radic. Biol. Med. 2013, 61, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhao, W.; Hu, Z.-J.; Ge, S.-M.; Huo, Y.; Liu, L.-X.; Gao, B.-L. Protective effects and mechanisms of high-dose vitamin C on sepsis-associated cognitive impairment in rats. Sci. Rep. 2021, 11, 14511. [Google Scholar] [CrossRef]
- Lee, J.-M.; Lee, J.-H.; Song, M.-K.; Kim, Y.-J. Nxp031 improves cognitive impairment in a chronic cerebral hypoperfusion-induced vascular dementia rat model through nrf2 signaling. Int. J. Mol. Sci. 2021, 22, 6285. [Google Scholar] [CrossRef]
- De Nuccio, F.; Cianciulli, A.; Porro, C.; Kashyrina, M.; Ruggiero, M.; Calvello, R.; Miraglia, A.; Nicolardi, G.; Lofrumento, D.D.; Panaro, M.A. Inflammatory Response Modulation by Vitamin C in an MPTP Mouse Model of Parkinson’s Disease. Biology 2021, 10, 1155. [Google Scholar] [CrossRef] [PubMed]
- Song, M.K.; Adams, L.; Lee, J.H.; Kim, Y.-S. NXP031 prevents dopaminergic neuronal loss and oxidative damage in the AAV-WT-α-synuclein mouse model of Parkinson’s disease. PLoS ONE 2022, 17, e0272085. [Google Scholar] [CrossRef]
- Wąsik, A.; Antkiewicz-Michaluk, L. The mechanism of neuroprotective action of natural compounds. Pharmacol. Rep. 2017, 69, 851–860. [Google Scholar] [CrossRef]
- Moretti, M.; Fraga, D.B.; Rodrigues, A.L.S. Preventive and therapeutic potential of ascorbic acid in neurodegenerative diseases. CNS Neurosci. Ther. 2017, 23, 921–929. [Google Scholar] [CrossRef]
- Bayo-Olugbami, A.; Nafiu, A.B.; Amin, A.; Ogundele, O.M.; Lee, C.C.; Owoyele, B.V. Vitamin D attenuated 6-OHDA-induced behavioural deficits, dopamine dysmetabolism, oxidative stress, and neuro-inflammation in mice. Nutr. Neurosci. 2022, 25, 823–834. [Google Scholar] [CrossRef]
- Kimlin, M.G. Geographic location and vitamin D synthesis. Mol. Asp. Med. 2008, 29, 453–461. [Google Scholar] [CrossRef]
- Yeum, K.-J.; Song, B.C.; Joo, N.-S. Impact of geographic location on vitamin D status and bone mineral density. Int. J. Environ. Res. Public Health 2016, 13, 184. [Google Scholar] [CrossRef]
- Lv, L.; Tan, X.; Peng, X.; Bai, R.; Xiao, Q.; Zou, T.; Tan, J.; Zhang, H.; Wang, C. The relationships of vitamin D, vitamin D receptor gene polymorphisms, and vitamin D supplementation with Parkinson’s disease. Transl. Neurodegener. 2020, 9, 34. [Google Scholar] [CrossRef]
- Zhang, H.J.; Zhang, J.R.; Mao, C.J.; Li, K.; Wang, F.; Chen, J.; Liu, C.F. Relationship between 25-Hydroxyvitamin D, bone density, and Parkinson’s disease symptoms. Acta Neurol. Scand. 2019, 140, 274–280. [Google Scholar] [CrossRef]
- Sleeman, I.; Aspray, T.; Lawson, R.; Coleman, S.; Duncan, G.; Khoo, T.K.; Schoenmakers, I.; Rochester, L.; Burn, D.; Yarnall, A. The role of vitamin D in disease progression in early Parkinson’s disease. J. Park. Dis. 2017, 7, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; He, Y.; Beck, J.; da Silva Teixeira, S.; Harrison, K.; Xu, Y.; Sisley, S. Defining vitamin D receptor expression in the brain using a novel VDRCre mouse. J. Comp. Neurol. 2021, 529, 2362–2375. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, N.; Lu, Y.; Tan, K. Vitamin D receptor polymorphisms and the susceptibility of Parkinson’s disease. Neurosci. Lett. 2019, 699, 206–211. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhou, R.; Zhang, Z.; Li, K. The association between vitamin D status, vitamin D supplementation, sunlight exposure, and Parkinson’s disease: A systematic review and meta-analysis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 666. [Google Scholar] [CrossRef] [PubMed]
- Gatto, N.M.; Paul, K.C.; Sinsheimer, J.S.; Bronstein, J.M.; Bordelon, Y.; Rausch, R.; Ritz, B. Vitamin D receptor gene polymorphisms and cognitive decline in Parkinson’s disease. J. Neurol. Sci. 2016, 370, 100–106. [Google Scholar] [CrossRef]
- Kim, J.E.; Oh, E.; Park, J.; Youn, J.; Kim, J.S.; Jang, W. Serum 25-hydroxyvitamin D3 level may be associated with olfactory dysfunction in de novo Parkinson’s disease. J. Clin. Neurosci. 2018, 57, 131–135. [Google Scholar] [CrossRef]
- Calvello, R.; Cianciulli, A.; Nicolardi, G.; De Nuccio, F.; Giannotti, L.; Salvatore, R.; Porro, C.; Trotta, T.; Panaro, M.A.; Lofrumento, D.D. Vitamin D treatment attenuates neuroinflammation and dopaminergic neurodegeneration in an animal model of Parkinson’s disease, shifting M1 to M2 microglia responses. J. Neuroimmune Pharmacol. 2017, 12, 327–339. [Google Scholar] [CrossRef]
- Lima, L.A.; Lopes, M.J.P.; Costa, R.O.; Lima, F.A.V.; Neves, K.R.T.; Calou, I.B.; Andrade, G.M.; Viana, G.S. Vitamin D protects dopaminergic neurons against neuroinflammation and oxidative stress in hemiparkinsonian rats. J. Neuroinflammation 2018, 15, 249. [Google Scholar] [CrossRef] [PubMed]
- Abu-Elfotuh, K.; Hamdan, A.M.E.; Abbas, A.N.; Alahmre, A.T.S.; Elewa, M.A.; Masoud, R.A.E.; Ali, A.A.; Othman, M.; Kamal, M.M.; Hassan, F.A.M. Evaluating the neuroprotective activities of vinpocetine, punicalagin, niacin and vitamin E against behavioural and motor disabilities of manganese-induced Parkinson’s disease in Sprague Dawley rats. Biomed. Pharmacother. 2022, 153, 113330. [Google Scholar] [CrossRef]
- Iqbal, A.; Anwar, F.; Saleem, U.; Khan, S.S.; Karim, A.; Ahmad, B.; Gul, M.; Iqbal, Z.; Ismail, T. Inhibition of Oxidative Stress and the NF-κB Pathway by a Vitamin E Derivative: Pharmacological Approach against Parkinson’s Disease. ACS Omega 2022, 7, 45088–45095. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Nehru, B. Beneficial effect of vitamin E in rotenone induced model of PD: Behavioural, neurochemical and biochemical study. Exp. Neurobiol. 2013, 22, 214. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-M.; Elliot, J.; Hobson, P.; O’Hare, E. Effects of intrahippocampal NAC61–95 injections on memory in the rat and attenuation with vitamin E. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Kwak, S.G.; Kwak, S. Effect of dietary vitamins C and E on the risk of Parkinson’s disease: A meta-analysis. Clin. Nutr. 2021, 40, 3922–3930. [Google Scholar] [CrossRef]
- Ricciarelli, R.; Argellati, F.; Pronzato, M.A.; Domenicotti, C. Vitamin E and neurodegenerative diseases. Mol. Asp. Med. 2007, 28, 591–606. [Google Scholar] [CrossRef] [PubMed]
- Taghizadeh, M.; Tamtaji, O.R.; Dadgostar, E.; Kakhaki, R.D.; Bahmani, F.; Abolhassani, J.; Aarabi, M.H.; Kouchaki, E.; Memarzadeh, M.R.; Asemi, Z. The effects of omega-3 fatty acids and vitamin E co-supplementation on clinical and metabolic status in patients with Parkinson’s disease: A randomized, double-blind, placebo-controlled trial. Neurochem. Int. 2017, 108, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-X.; Yu, X.-D.; Cheng, Q.-Z.; Tang, L.; Shen, M.-Q. The association of serum vitamin K2 levels with Parkinson’s disease: From basic case-control study to big data mining analysis. Aging 2020, 12, 16410. [Google Scholar] [CrossRef]
- Vos, M.; Esposito, G.; Edirisinghe, J.N.; Vilain, S.; Haddad, D.M.; Slabbaert, J.R.; Van Meensel, S.; Schaap, O.; De Strooper, B.; Meganathan, R. Vitamin K2 is a mitochondrial electron carrier That rescues pink1 deficiency. Science 2012, 336, 1306–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, F.L.; Cerqueira, E.C.; de Freitas, M.S.; Gonçalves, D.L.; Costa, L.T.; Follmer, C. Vitamins K interact with N-terminus α-synuclein and modulate the protein fibrillization in vitro. Exploring the interaction between quinones and α-synuclein. Neurochem. Int. 2013, 62, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-X.; Li, Y.-P.; Gao, F.; Hu, Q.-S.; Zhang, Y.; Chen, D.; Wang, G.-H. Vitamin K2 suppresses rotenone-induced microglial activation in vitro. Acta Pharmacol. Sin. 2016, 37, 1178–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vitamin | Study Design | Sample Size | Results | Vitamin and PD Association | Ref. |
|---|---|---|---|---|---|
| Vitamin A | Cohort-study | 42 PD patients and 42 healthy controls | No significant difference in the serum levels of vitamin A and retinol-binding protein between the control and PD group | No association between vitamin A and the risk of PD | [24] |
| Population-based cohort study | 63,257 men and women, including 544 patients with incident PD | No precise dose-dependent association between dietary intake of vitamin A, E, and C and the risk of PD | No association between vitamin A and the risk of PD | [25] | |
| Vitamin B1 | Case-study | 5 PD patients | Intake of daily 100–200 mg doses of parenteral thiamine improved movement, arm swings, and tremors in thiamine deficient PD patients | TD is associated with an increased risk of PD, and its supplementation may be beneficial | [27] |
| Case-study | 3 PD patients | A high dose of thiamine intake significantly improved motor coordination-related UPDRS, ranging from 31.3% to 77.3% | A high dose of thiamine intake is associated with improvement in PD symptoms | [28] | |
| Case-controlled study | 96 PD patients and 375 control subjects | Deficiency of thiamine and folate caused olfactory dysfunction in PD patients | TD is associated with an increased risk of PD | [29] | |
| Case-controlled study | 75 PD patients and 24 control subjects | In male PD patients, higher levels of phosphate and thiamine concentration, as well as higher MNA-total score, were correlated with a lower risk of MCI | Thiamine insufficiency and low phosphate levels increase the risk for PD-associated cognitive deficits | [30] | |
| Vitamin B3 | Case-controlled study | 46 PD patients | PD patients with vitamin B3 deficiency were associated with GPR109A-mediated inflammation. Supplementation with 100 mg and 200 mg doses showed ameliorative effect | Vitamin B3 deficiency is associated with an increased risk of PD. | [38] |
| Vitamin B6 | Case-controlled study | 249 PD patients and 368 control subjects | Low consumption of vitamin B6 was associated with an elevated risk of PD | A deficiency of vitamin B6 increases the risk of PD | [51] |
| Case-study | 83-year-old woman with hypertension, coronary artery disease, and PD | Early detection and treatment of pyridoxine deficiency may reduce new-onset epileptic seizures and status epilepticus in PD patients | Vitamin B6 deficiency is associated with an increased risk of PD | [42] | |
| Population based cohort-study | 5289 total participants, including 72 patients with incident PD | No association between dietary folate and vitamin B12 and the risk of PD. Vitamin B6 decreased the risk of PD. | Dietary vitamin B6 correlated with reduced risk of PD | [19] | |
| Vitamin B12 | Population-based cohort study | NA | Higher serum vitamin B12 at baseline level of PD diagnosis was correlated with a reduced risk of dementia | Vitamin B12 is associated with decreased risk of PD | [46] |
| Longitudinal cohort- study | 1741 participants | A low hazard ratio in subjects taking vitamin B12 + MVI and MVI groups for developing sensory symptoms of PD | Vitamin B12 is associated with a reduced risk of PD | [47] | |
| Vitamin C | Cohort-study | 75 PD patients and 75 healthy subjects | Patients with PD had considerably increased nitrite oxide and peroxynitrite but low vitamin C levels in the serum | Vitamin C deficiency is associated with an increased risk of PD | [8] |
| Vitamin D | Cohort-study | 182 PD patients and 185 control subjects | PD patients had lower serum levels of 25 (OH)D than healthy controls | Vitamin D deficiency is associated with an increased risk of PD | [64] |
| Patients with MCI were categorized as serum 25 (OH)D deficient (n = 27) or not deficient (n = 29) based on serum 25 (OH)D levels. | In older persons with MCI, low vitamin D levels were related with lower volumes of hippocampus subfields and connection impairments, which aggravated neurocognitive results. | Low vitamin D is associated with progression from MCI to major cognitive disorders. | [17] | ||
| Observational study | 145 PD patients and 94 control subjects | PD patients had lower serum levels of 25 (OH)D than healthy controls at baseline and at 18th-month follow-up session | A deficiency of 25 (OH)D is associated with increased motor severity and risk of bone fracture in PD patients | [65] | |
| Meta-analysis | NA | Significant associations between rs2228570 and PD risk were found in allelic, dominant, and additive models but not in the recessive model. | VDR polymorphism is associated with an increased risk of PD | [67] | |
| Meta-analysis | NA | Both 25 (OH)D insufficiency and deficiency were correlated with an increased risk of PD. However, vitamin D supplementation did not improve motor symptoms in PD patients | Deficiency of 25 (OH)D and reduced exposure to sunlight is associated with an increased risk of PD | [68] | |
| Observational study | 39 drug-naive, de novo PD patients | Vitamin D was involved in the etiology of olfactory impairment in PD | Vitamin D deficiency increases the risk of olfactory dysfunction | [70] | |
| Vitamin E | Meta-analysis | NA | High vitamin E consumption considerably decreased the chance of developing PD | High vitamin E intake is associated with a reduced risk of PD | [77] |
| Randomized double-blind placebo-controlled study | 60 PD patients | Co-supplementation with omega-3 fatty acids and vitamin E improved UPDRS in persons with PD | Vitamin E supplementation is associated with decreased risk of PD in older adults | [79] | |
| Vitamin K | Case-controlled study | 93 PD patients and 95 healthy controls | PD patients were deficient in serum vitamin K2 level | Vitamin K2 deficiency is associated with an increased risk of PD | [80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandeep; Sahu, M.R.; Rani, L.; Kharat, A.S.; Mondal, A.C. Could Vitamins Have a Positive Impact on the Treatment of Parkinson’s Disease? Brain Sci. 2023, 13, 272. https://doi.org/10.3390/brainsci13020272
Sandeep, Sahu MR, Rani L, Kharat AS, Mondal AC. Could Vitamins Have a Positive Impact on the Treatment of Parkinson’s Disease? Brain Sciences. 2023; 13(2):272. https://doi.org/10.3390/brainsci13020272
Chicago/Turabian StyleSandeep, Manas Ranjan Sahu, Linchi Rani, Arun S. Kharat, and Amal Chandra Mondal. 2023. "Could Vitamins Have a Positive Impact on the Treatment of Parkinson’s Disease?" Brain Sciences 13, no. 2: 272. https://doi.org/10.3390/brainsci13020272