
The Role of Beta-Endorphin in Food Deprivation-Mediated Increases in Food Intake and Binge-Eating




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subject
2.2. Diet
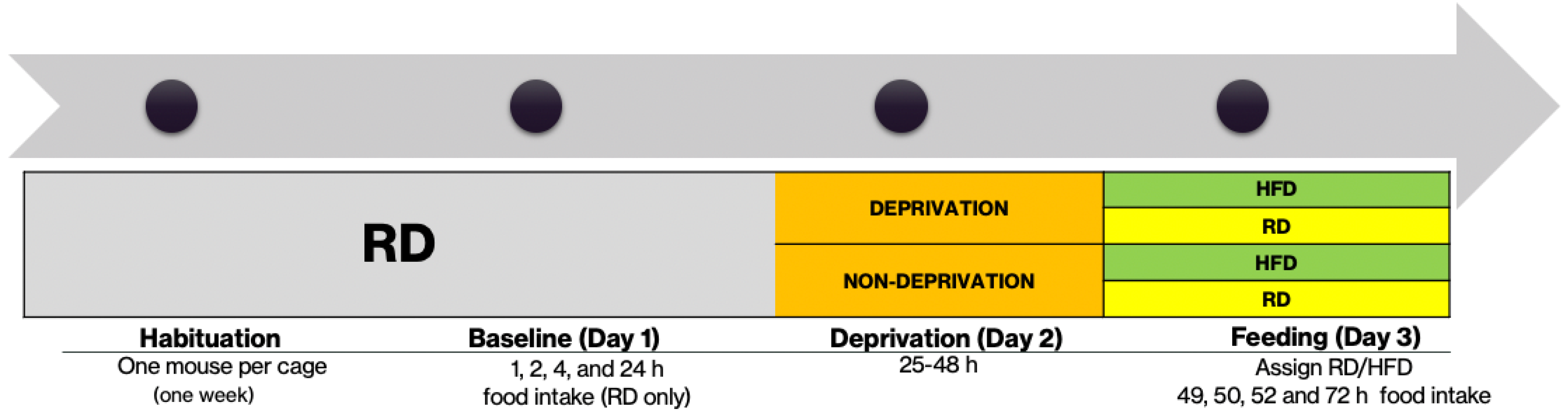
2.3. Food Deprivation Protocol
2.4. Binge-Eating Protocol
2.5. Statistical Analysis
3. Results
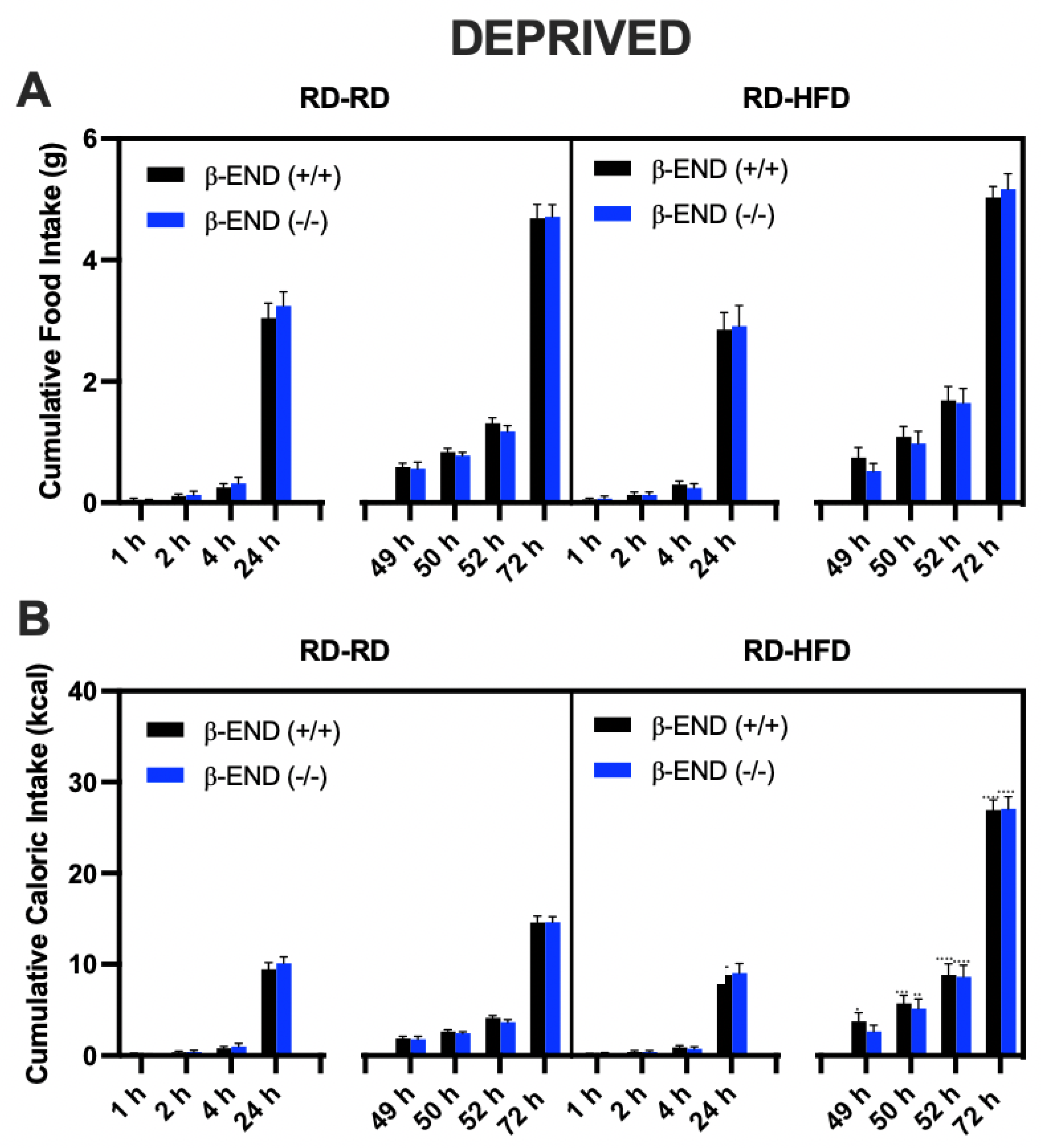
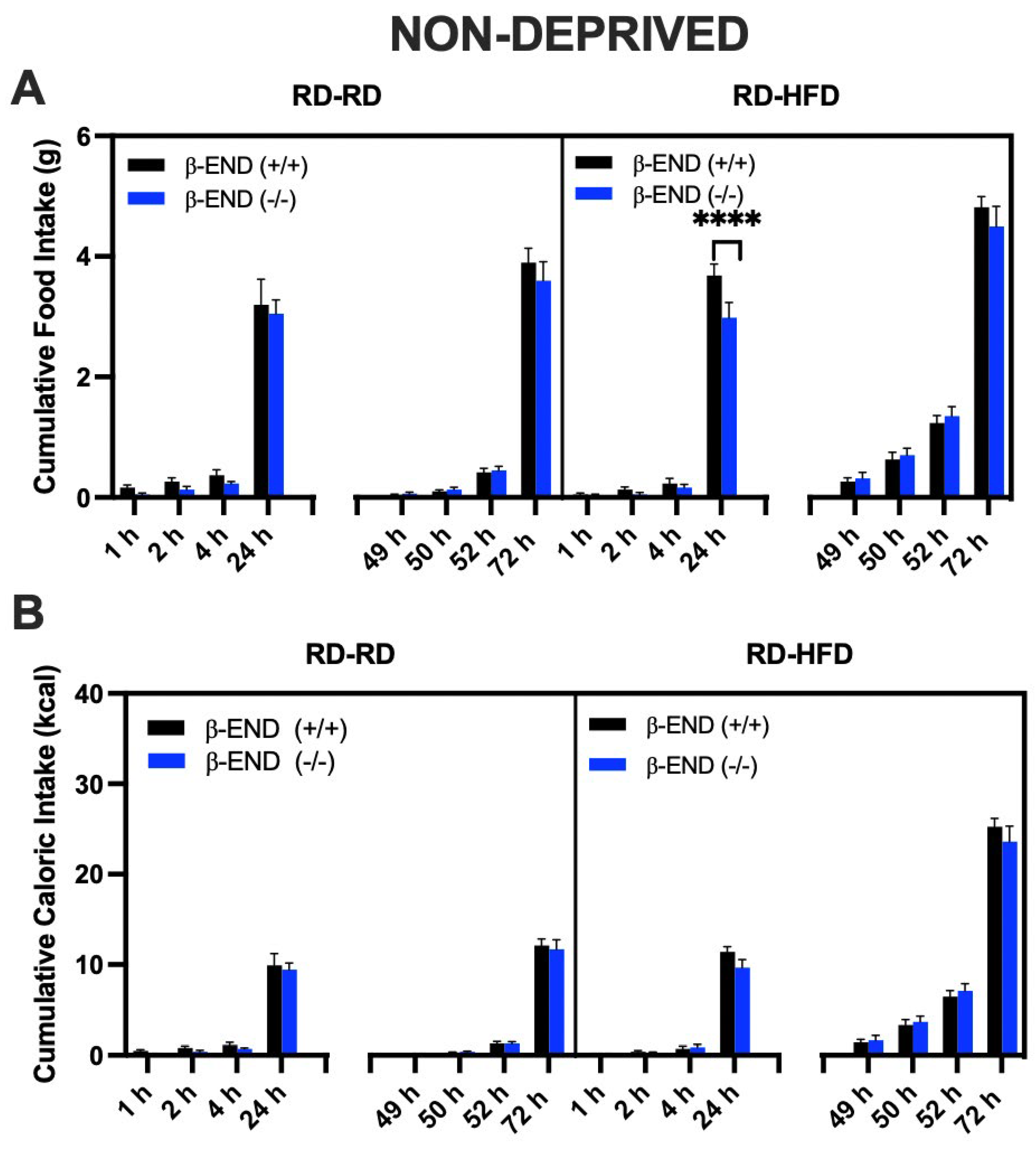
3.1. Food Intake Was Increased in Food-Deprived compared to Non-Deprived Mice Lacking Beta-Endorphin and Their Wildtype Controls
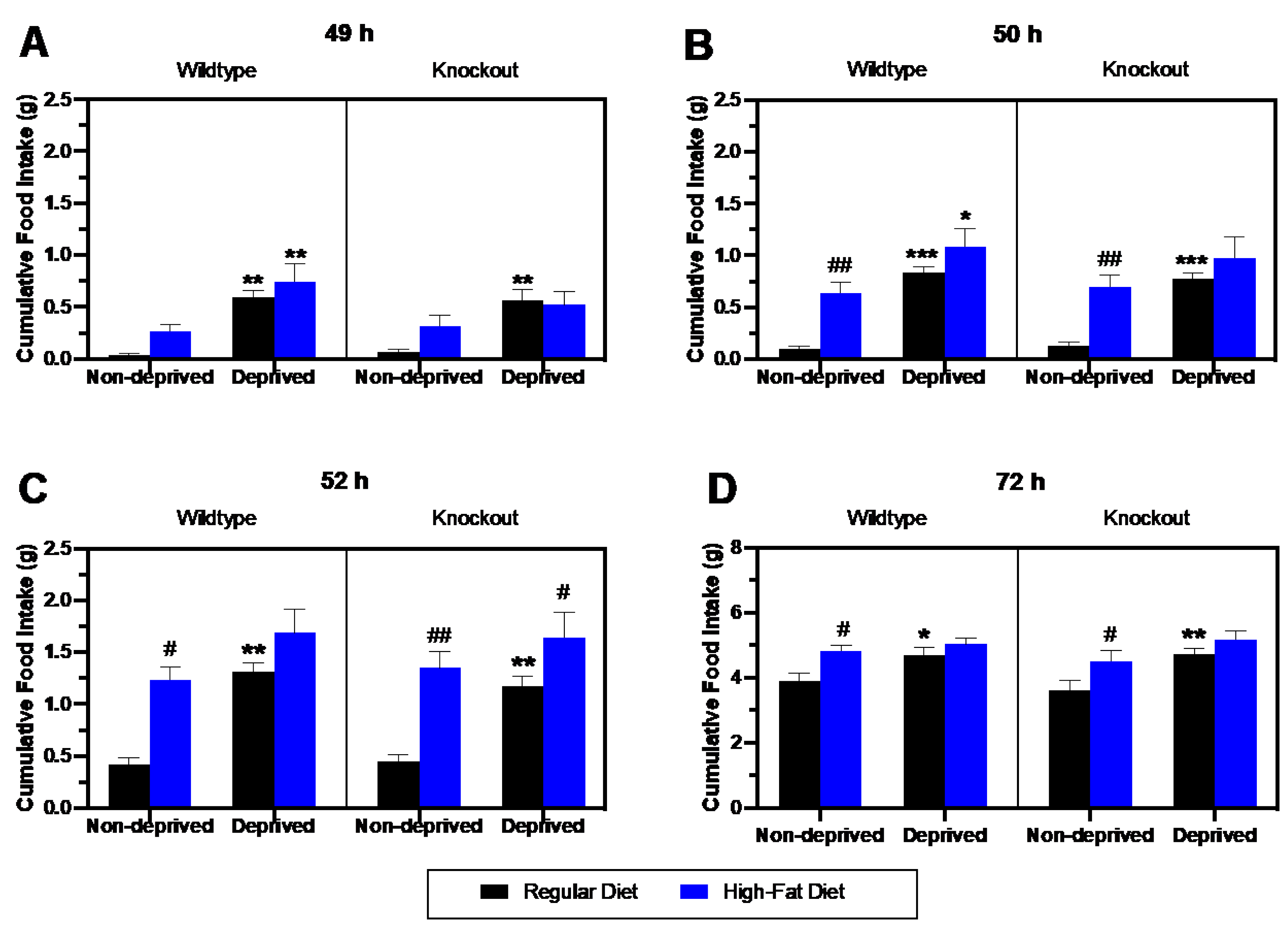
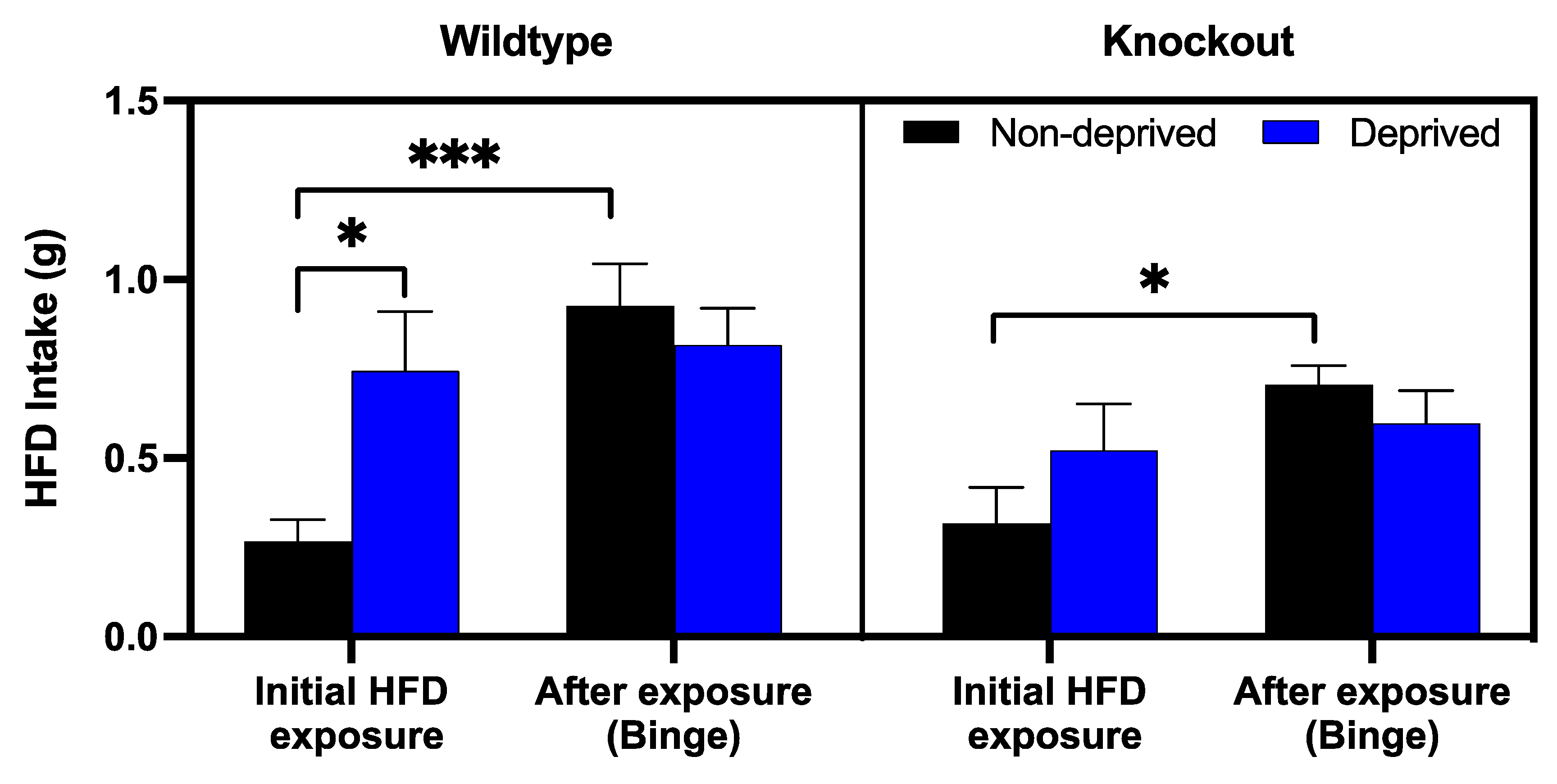
3.2. Comparison of Regular Diet vs. High-Fat Diet Intake in Deprived vs. Non-Deprived Mice Lacking Beta-Endorphin and Their Wildtype Controls
3.3. The Role of Beta-Endorphin in Binge Eating in Deprived and Non-Deprived Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Volkow, N.D.; Wang, G.; Baler, R. Reward, dopamine and the control of food intake: Implications for obesity. Trends Cogn. Sci. 2011, 15, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthoud, H.R.; Lenard, N.; Shin, A. Food reward, hyperphagia, and obesity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1266–R1277. [Google Scholar] [CrossRef] [Green Version]
- Luo, N.; Marcelin, G.; Liu, S.M.; Schwartz, G.; Chua, S., Jr. Neuropeptide Y and agouti-related peptide mediate complementary functions of hyperphagia and reduced energy expenditure in leptin receptor deficiency. Endocrinology 2011, 152, 883–889. [Google Scholar] [CrossRef] [Green Version]
- Barnes, M.J.; Primeaux, S.; Bray, G. Food deprivation increases the mRNA expression of micro-opioid receptors in the ventral medial hypothalamus and arcuate nucleus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R1385–R1390. [Google Scholar] [CrossRef] [PubMed]
- Robinson, B.; Moran, T. Acute food deprivation and chronic food restriction differentially affect hypothalamic NPY mRNA expression. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, R1030–R1036. [Google Scholar]
- Nathan, P.J.; Bullmore, E. From taste hedonics to motivational drive: Central mu-opioid receptors and binge-eating behaviour. Int. J. Neuropsychopharmacol. 2009, 12, 995–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjimarkou, M.M.; Singh, A.; Kandov, Y.; Israel, Y.; Pan, Y.X.; Rossi, G.C.; Pasternak, G.W.; Bodnar, R.J. Opioid receptor involvement in food deprivation-induced feeding: Evaluation of selective antagonist and antisense oligodeoxynucleotide probe effects in mice and rats. J. Pharmacol. Exp. Ther. 2004, 311, 1188–1202. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Holtzman, S. Suppression of deprivation-induced food and water intake in rats and mice by naloxone. Pharmacol. Biochem. Behav. 1979, 11, 567–573. [Google Scholar] [CrossRef]
- Levine, A.S.; Weldon, D.T.; Grace, M.; Cleary, J.P.; Billington, C.J. Naloxone blocks that portion of feeding driven by sweet taste in food-restricted rats. Am. J. Physiol. 1995, 268, R248–R252. [Google Scholar] [CrossRef]
- Hobbs, D.J.; Koch, J.; Bodnar, R. Naltrexone, dopamine receptor agonists and antagonists, and food intake in rats: 1. Food deprivation. Pharmacol. Biochem. Behav. 1994, 49, 197–204. [Google Scholar] [CrossRef]
- Nogueiras, R.; Romero-Picó, A.; Vazquez, M.J.; Novelle, M.G.; López, M.; Diéguez, C. The opioid system and food intake: Homeostatic and hedonic mechanisms. Obes. Facts 2012, 5, 196–207. [Google Scholar] [CrossRef]
- Knuth, U.A.; Friesen, H. Changes of beta-endorphin and somatostatin concentrations in different hypothalamic areas of female rats after chronic starvation. Life Sci. 1983, 33, 827–833. [Google Scholar] [CrossRef]
- Majeed, N.H.; Lasoń, W.; Przewłocka, B.; Przewłocki, R. Brain and peripheral opioid peptides after changes in ingestive behavior. Neuroendocrinology 1986, 42, 267–272. [Google Scholar] [CrossRef]
- Mitev, Y.; Almeida, O.; Patchev, V. Pituitary-adrenal function and hypothalamic beta-endorphin release in vitro following food deprivation. Brain Res. Bull. 1993, 30, 7–10. [Google Scholar] [CrossRef]
- Kim, E.M.; Welch, C.C.; Grace, M.K.; Billington, C.J.; Levine, A.S. Chronic food restriction and acute food deprivation decrease mRNA levels of opioid peptides in arcuate nucleus. Am. J. Physiol. 1996, 270, R1019–R1024. [Google Scholar] [CrossRef]
- Daimon, C.M.; Hentges, S.T. β-endorphin differentially contributes to food anticipatory activity in male and female mice undergoing activity-based anorexia. Physiol. Rep. 2021, 9, e14788. [Google Scholar] [CrossRef]
- Rubinstein, M.; Mogil, J.S.; Japón, M.; Chan, E.C.; Allen, R.G.; Low, M.J. Absence of opioid stress-induced analgesia in mice lacking beta-endorphin by site-directed mutagenesis. Proc. Natl. Acad. Sci. USA 1996, 93, 3995–4000. [Google Scholar] [CrossRef] [Green Version]
- Cabeza de Vaca, S.; Carr, K.D. Food restriction enhances the central rewarding effect of abused drugs. J. Neurosci. 1998, 18, 7502–7510. [Google Scholar] [CrossRef] [Green Version]
- Jewett, D.C.; Klockars, A.; Smith, T.R.; Brunton, C.; Head, M.A.; Tham, R.L.; Kwilasz, A.J.; Hahn, T.W.; Wiebelhaus, J.M.; Ewan, E.E.; et al. Effects of opioid receptor ligands in rats trained to discriminate 22 from 2 hours of food deprivation suggest a lack of opioid involvement in eating for hunger. Behav. Brain Res. 2020, 380, 112369. [Google Scholar] [CrossRef]
- Giuliano, C.; Robbins, T.W.; Nathan, P.J.; Bullmore, E.T.; Everitt, B.J. Inhibition of opioid transmission at the mu-opioid receptor prevents both food seeking and binge-like eating. Neuropsychopharmacology 2012, 37, 2643–2652. [Google Scholar] [CrossRef] [Green Version]
- Hayward, M.D.; Pintar, J.; Low, M. Selective reward deficit in mice lacking beta-endorphin and enkephalin. J. Neurosci. 2002, 22, 8251–8258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gheibi, N.; Saroukhani, M.; Azhdari-Zarmehri, H. The effect of food deprivation on nociception in formalin test and plasma levels of noradrenaline and corticosterone in rats. Basic Clin. Neurosci. 2013, 4, 341–347. [Google Scholar] [PubMed]
- Knuth, U.A.; Sikand, G.S.; Casanueva, F.F.; Havlicek, V.; Friesen, H.G. Changes in beta-endorphin content in discrete areas of the hypothalamus throughout proestrus and diestrus of the rat. Life Sci. 1983, 33, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Frank, A.; Brown, L.; Clegg, D. The role of hypothalamic estrogen receptors in metabolic regulation. Front. Neuroendocrinol. 2014, 35, 550–557. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tolentino, L.; Iqbal, A.; Rahman, S.; Lutfy, K. The Role of Beta-Endorphin in Food Deprivation-Mediated Increases in Food Intake and Binge-Eating. Brain Sci. 2023, 13, 212. https://doi.org/10.3390/brainsci13020212
Tolentino L, Iqbal A, Rahman S, Lutfy K. The Role of Beta-Endorphin in Food Deprivation-Mediated Increases in Food Intake and Binge-Eating. Brain Sciences. 2023; 13(2):212. https://doi.org/10.3390/brainsci13020212
Chicago/Turabian StyleTolentino, Laica, Asif Iqbal, Shafiqur Rahman, and Kabirullah Lutfy. 2023. "The Role of Beta-Endorphin in Food Deprivation-Mediated Increases in Food Intake and Binge-Eating" Brain Sciences 13, no. 2: 212. https://doi.org/10.3390/brainsci13020212