

Actin Alpha 2, Smooth Muscle (ACTA2) Is Involved in the Migratory Potential of Malignant Gliomas, and Its Increased Expression at Recurrence Is a Significant Adverse Prognostic Factor
 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Droplet Digital Polymerase Chain Reaction (dd-PCR)
2.3. Small Interfering RNA Transfection
2.4. Proliferation Assay
2.5. Migration Assay
2.6. Immunohistochemistry
2.7. Clinical Tumor Samples and Medical Information
2.8. Validation Using Data from the TCGA Database’s Cohort of Glioma Patients
2.9. Statistical Analysis
3. Results
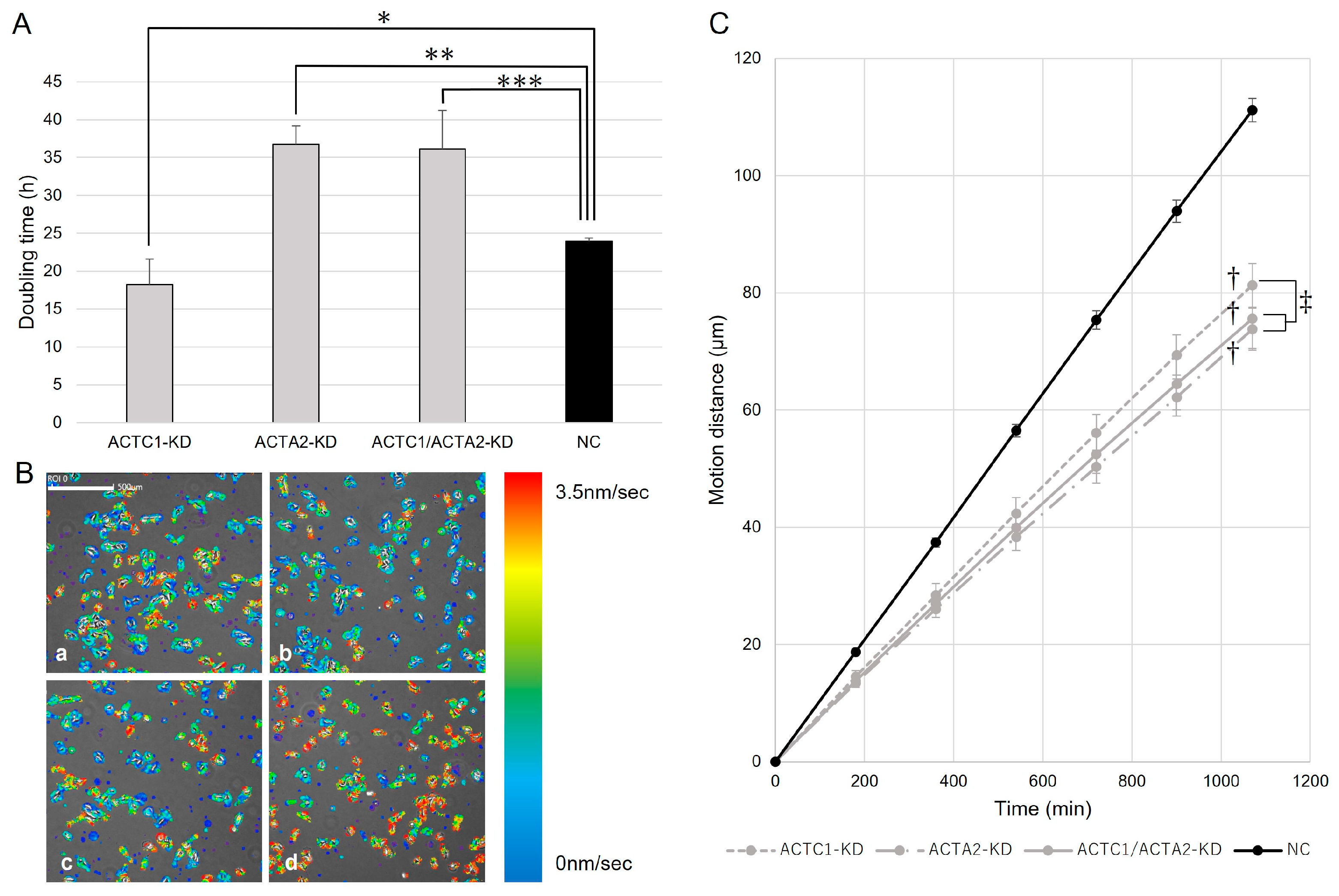
3.1. ACTC1 and ACTA2 KD through siRNA Transfection
3.2. Cell Proliferation of U251MG
3.3. Cell Migration of U251MG
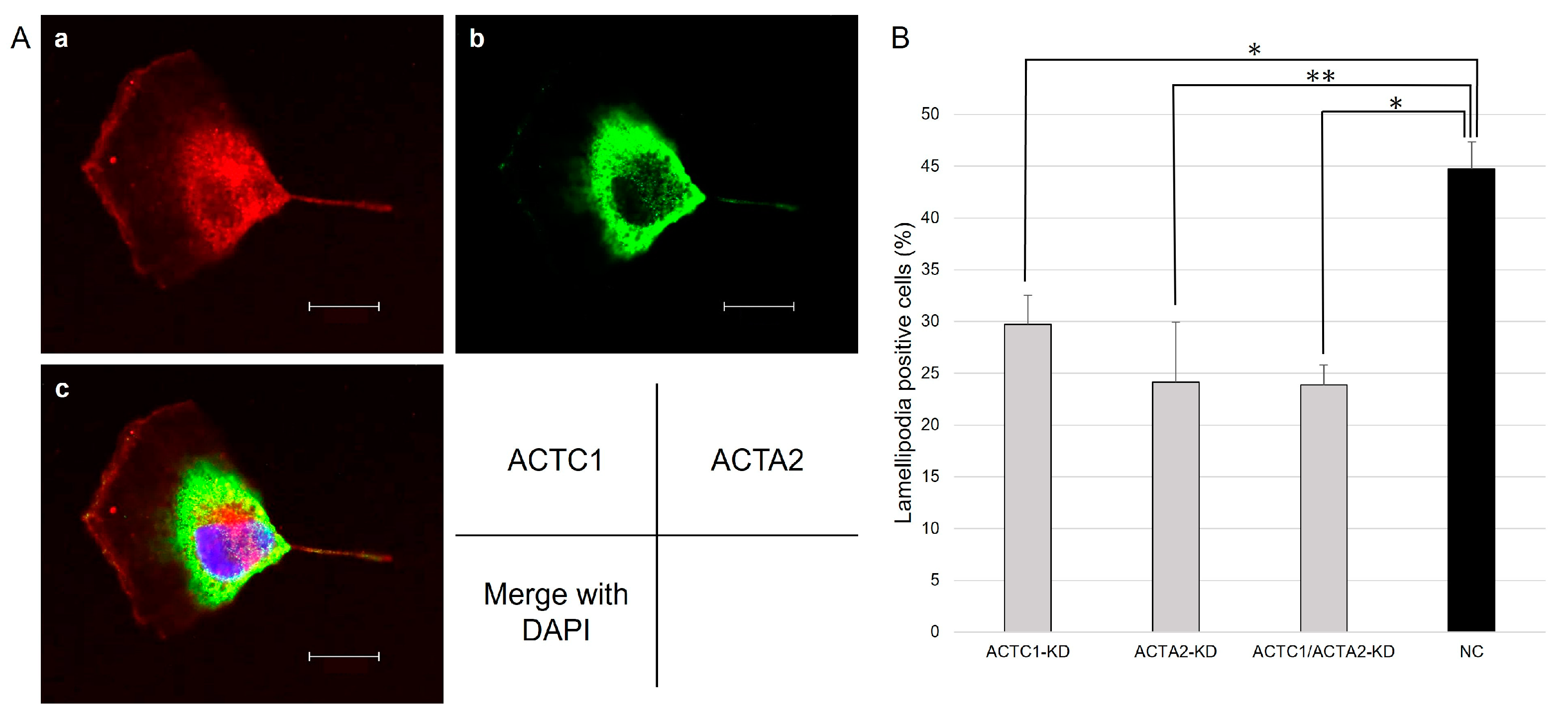
3.4. Immunohistochemistry
3.5. Lamellipodia Formation of U251MG
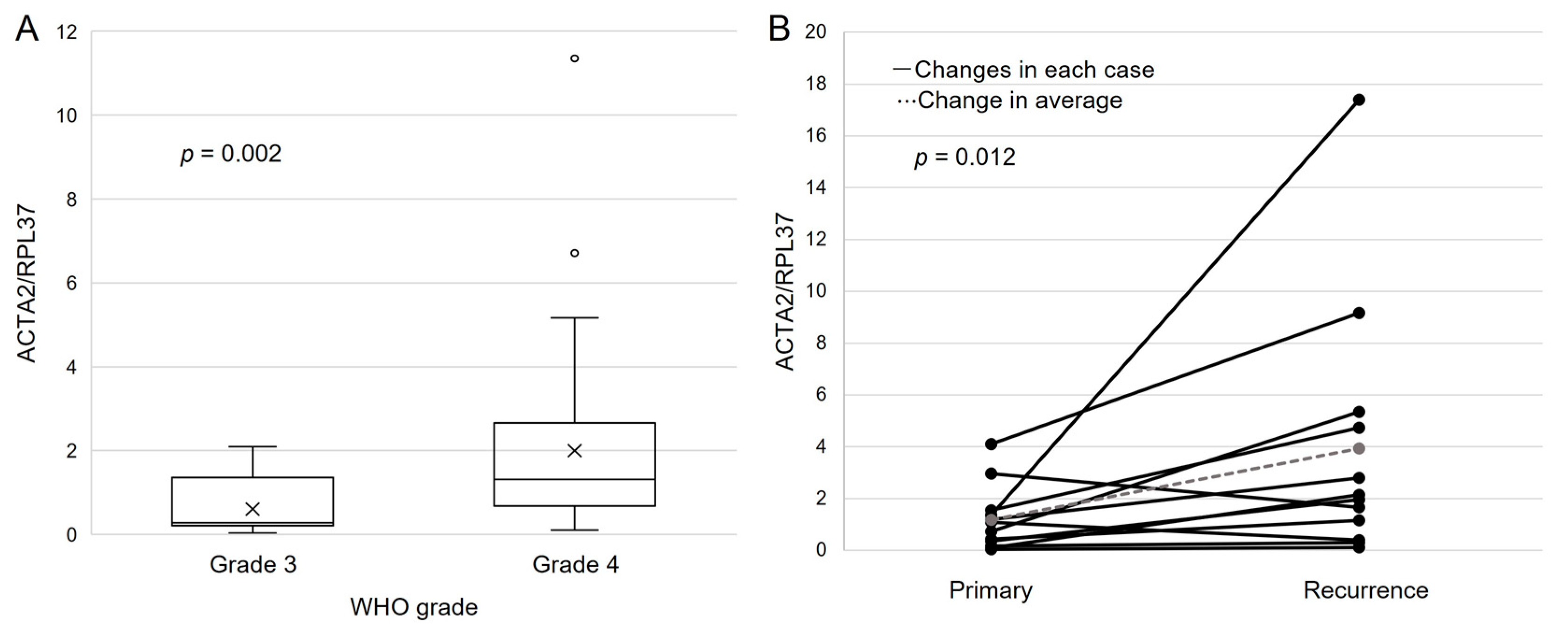
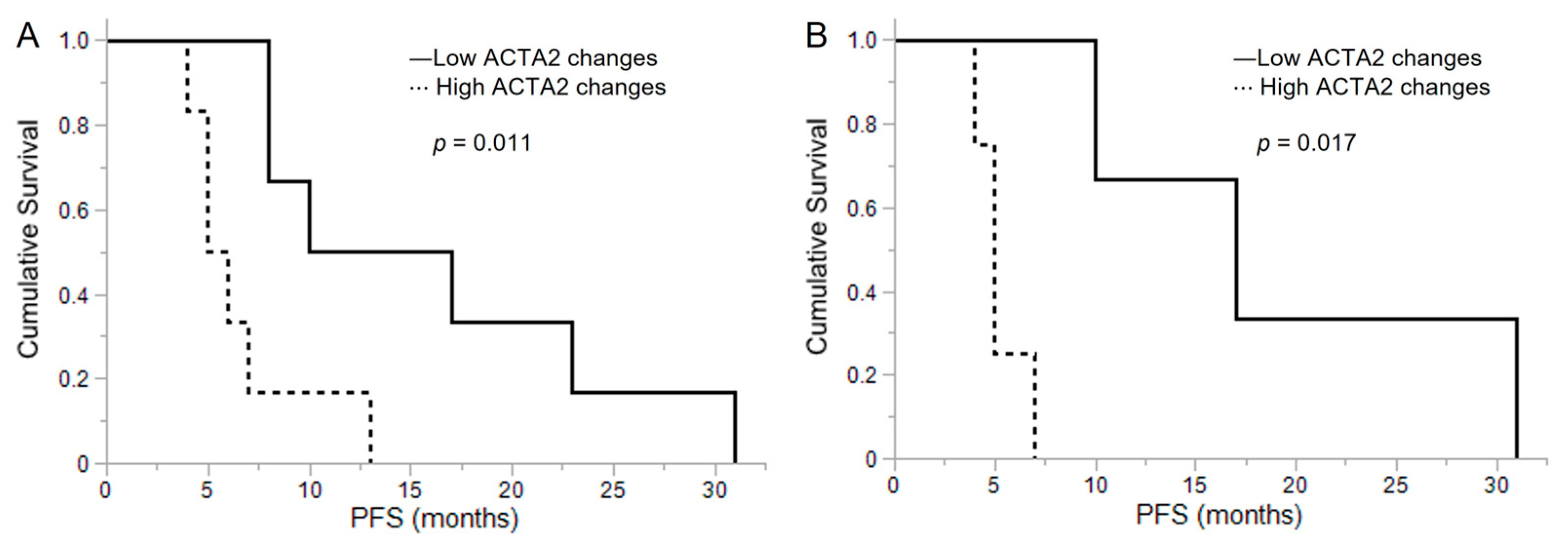
3.6. Clinical Data of Malignant Glioma Patients
3.7. ACTA2 Expression in Primary Malignant Glioma
3.8. ACTA2 Expression in Recurrent Malignant Glioma
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Cioffi, G.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS statistical report: Primary brain and other central nervous system tumors diagnosed in the United States in 2014–2018. Neuro Oncol. 2021, 23, iii1–iii105. [Google Scholar] [CrossRef] [PubMed]
- Narita, Y.; Shibui, S.; Committee of Brain Tumor Registry of Japan Supported by the Japan Neurosurgical Society. Trends and outcomes in the treatment of gliomas based on data during 2001–2004 from the Brain Tumor Registry of Japan. Neurol. Med. Chir. 2015, 55, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Claes, A.; Idema, A.J.; Wesseling, P. Diffuse glioma growth: A guerilla war. Acta Neuropathol. 2007, 114, 443–458. [Google Scholar] [CrossRef]
- de Vrij, J.; Maas, S.L.; Kwappenberg, K.M.; Schnoor, R.; Kleijn, A.; Dekker, L.; Luider, T.M.; de Witte, L.D.; Litjens, M.; van Strien, M.E.; et al. Glioblastoma-derived extracellular vesicles modify the phenotype of monocytic cells. Int. J. Cancer 2015, 137, 1630–1642. [Google Scholar] [CrossRef] [PubMed]
- Schiera, G.; Di Liegro, C.M.; Di Liegro, I. Extracellular Membrane Vesicles as Vehicles for Brain Cell-to-Cell Interactions in Physiological as well as Pathological Conditions. Biomed. Res. Int. 2015, 2015, 152926. [Google Scholar] [CrossRef] [PubMed]
- Schiera, G.; Di Liegro, C.M.; Di Liegro, I. Molecular Determinants of Malignant Brain Cancers: From Intracellular Alterations to Invasion Mediated by Extracellular Vesicles. Int. J. Mol. Sci. 2017, 18, 2774. [Google Scholar] [CrossRef]
- Pollard, T.D.; Cooper, J.A. Actin, a central player in cell shape and movement. Science 2009, 326, 1208–1212. [Google Scholar] [CrossRef]
- Suresh, R.; Diaz, R.J. The remodeling of actin composition as a hallmark of cancer. Transl. Oncol. 2021, 14, 101051. [Google Scholar] [CrossRef]
- Ohtaki, S.; Wanibuchi, M.; Kataoka-Sasaki, Y.; Sasaki, M.; Oka, S.; Noshiro, S.; Akiyama, Y.; Mikami, T.; Mikuni, N.; Kocsis, J.D.; et al. ACTC1 as an invasion and prognosis marker in glioma. J. Neurosurg. 2017, 126, 467–475. [Google Scholar] [CrossRef]
- Wanibuchi, M.; Ohtaki, S.; Ookawa, S.; Kataoka-Sasaki, Y.; Sasaki, M.; Oka, S.; Kimura, Y.; Akiyama, Y.; Mikami, T.; Mikuni, N.; et al. Actin, alpha, cardiac muscle 1 (ACTC1) knockdown inhibits the migration of glioblastoma cells in vitro. J. Neurol. Sci. 2018, 392, 117–121. [Google Scholar] [CrossRef]
- Yuan, S.M. α-smooth muscle actin and ACTA2 gene expressions in vasculopathies. Braz. J. Cardiovasc. Surg. 2015, 30, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Bowman, R.L.; Wang, Q.; Carro, A.; Verhaak, R.G.; Squatrito, M. GlioVis data portal for visualization and analysis of brain tumor expression datasets. Neuro Oncol. 2017, 19, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Small, J.V.; Stradal, T.; Vignal, E.; Rottner, K. The lamellipodium: Where motility begins. Trends Cell Biol. 2002, 12, 112–120. [Google Scholar] [CrossRef]
- Yamazaki, D.; Kurisu, S.; Takenawa, T. Regulation of cancer cell motility through actin reorganization. Cancer Sci. 2005, 96, 379–386. [Google Scholar] [CrossRef]
- Jeon, M.; You, D.; Bae, S.Y.; Kim, S.W.; Nam, S.J.; Kim, H.H.; Kim, S.; Lee, J.E. Dimerization of EGFR and HER2 induces breast cancer cell motility through STAT1-dependent ACTA2 induction. Oncotarget 2017, 8, 50570–50581. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Chen, Y.; Chen, M.; Wang, S.; Wen, X.; Zhang, S. Identification of key candidate genes and biological pathways in bladder cancer. PeerJ. 2018, 6, e6036. [Google Scholar] [CrossRef]
- Zhao, B.; Baloch, Z.; Ma, Y.; Wan, Z.; Huo, Y.; Li, F.; Zhao, Y. Identification of potential key genes and pathways in early-onset colorectal cancer through bioinformatics analysis. Cancer Control. 2019, 26, 1073274819831260. [Google Scholar] [CrossRef]
- Lee, H.W.; Seol, H.J.; Choi, Y.L.; Ju, H.J.; Joo, K.M.; Ko, Y.H.; Lee, J.I.; Nam, D.H. Genomic copy number alterations associated with the early brain metastasis of non-small cell lung cancer. Int. J. Oncol. 2012, 41, 2013–2020. [Google Scholar] [CrossRef]
- Lee, H.W.; Park, Y.M.; Lee, S.J.; Cho, H.J.; Kim, D.H.; Lee, J.I.; Kang, M.S.; Seol, H.J.; Shim, Y.M.; Nam, D.H.; et al. Alpha-smooth muscle actin (ACTA2) is required for metastatic potential of human lung adenocarcinoma. Clin. Cancer Res. 2013, 19, 5879–5889. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, X.; Zhang, C.; Zhong, J.; Fang, X.; Li, H.; Xie, F.; Huang, X.; Zhang, X.; Hu, Q.; et al. Actin alpha 2 (ACTA2) downregulation inhibits neural stem cell migration through rho GTPase activation. Stem Cells Int. 2020, 2020, 4764012. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Che, C.L.; Zhang, Y.M.; Zhang, H.H.; Sang, Y.L.; Lu, B.; Dong, F.S.; Zhang, L.J.; Lv, F.Z. DNA microarray reveals different pathways responding to paclitaxel and docetaxel in non-small cell lung cancer cell line. Int. J. Clin. Exp. Pathol. 2013, 6, 1538–1548. [Google Scholar] [PubMed]
- Yang, M.; Li, H.; Li, Y.; Ruan, Y.; Quan, C. Identification of genes and pathways associated with MDR in MCF-7/MDR breast cancer cells by RNA-seq analysis. Mol. Med. Rep. 2018, 17, 6211–6226. [Google Scholar] [CrossRef]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; van den Bent, M.J.; Taphoorn, M.J.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomized phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Marsh, D.; Suchak, K.; Moutasim, K.A.; Vallath, S.; Hopper, C.; Jerjes, W.; Upile, T.; Kalavrezos, N.; Violette, S.M.; Weinreb, P.H.; et al. Stromal features are predictive of disease mortality in oral cancer patients. J. Pathol. 2011, 223, 470–481. [Google Scholar] [CrossRef]
- Öhlund, D.; Handly-Santana, A.; Biffi, G.; Elyada, E.; Almeida, A.S.; Ponz-Sarvise, M.; Corbo, V.; Oni, T.E.; Hearn, S.A.; Lee, E.J.; et al. Distinct populations of inflammatory fibroblasts and myofibroblasts in pancreatic cancer. J. Exp. Med. 2017, 214, 579–596. [Google Scholar] [CrossRef]
- Lavie, D.; Ben-Shmuel, A.; Erez, N.; Scherz-Shouval, R. Cancer-associated fibroblasts in the single-cell era. Nat. Cancer 2022, 3, 793–807. [Google Scholar] [CrossRef]
- Vedula, P.; Kashina, A. The makings of the “actin code”: Regulation of actin’s biological function at the amino acid and nucleotide level. J. Cell Sci. 2018, 131, jcs215509. [Google Scholar] [CrossRef]
- Kumar, A.; Crawford, K.; Close, L.; Madison, M.; Lorenz, J.; Doetschman, T.; Pawlowski, S.; Duffy, J.; Neumann, J.; Robbins, J.; et al. Rescue of cardiac alpha-actin-deficient mice by enteric smooth muscle gamma-actin. Proc. Natl. Acad. Sci. USA 1997, 94, 4406–4411. [Google Scholar] [CrossRef]
- Nowak, K.J.; Ravenscroft, G.; Jackaman, C.; Filipovska, A.; Davies, S.M.; Lim, E.M.; Squire, S.E.; Potter, A.C.; Baker, E.; Clément, S.; et al. Rescue of skeletal muscle alpha-actin-null mice by cardiac (fetal) alpha-actin. J. Cell Biol. 2009, 185, 903–915. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Malignant Glioma (n = 50) | Recurrent Malignant Glioma (n = 12) | ||
|---|---|---|---|
| Age at diagnosis, median (range) | 64.5 (19–91) | 48 (30–80) | |
| Sex | |||
| Male | 32 (64%) | 9 (75%) | |
| Female | 18 (36%) | 3 (25%) | |
| WHO grade | |||
| 3 | 11 (22%) | 5 (42%) | |
| 4 | 39 (78%) | 7 (58%) | |
| PFS (month), median (range) | 9 (1–104) | 8 (4–31) | |
| OS (month), median (range) | 19 (1–104) | 31.5 (11–80) | |
| Postoperative therapy | |||
| Radiotherapy | 48 (96%) | 11 (92%) | |
| Chemotherapy | 48 (96%) | 12 (100%) | |
| ACTC1 Expression | ACTA2 Expression | Cell Proliferation | Cell Motility | Lamellipodia Formation | |
|---|---|---|---|---|---|
| ACTC1-KD | ↓ | ↑ | ↑ | ↓ | ↓ |
| ACTA2-KD | ↑ | ↓ | ↓ | ↓ | ↓ |
| ACTC1/ACTA2 double KD | ↓ | ↓ | ↓ | ↓ | ↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoshimaru, T.; Nonoguchi, N.; Kosaka, T.; Furuse, M.; Kawabata, S.; Yagi, R.; Kurisu, Y.; Kashiwagi, H.; Kameda, M.; Takami, T.; et al. Actin Alpha 2, Smooth Muscle (ACTA2) Is Involved in the Migratory Potential of Malignant Gliomas, and Its Increased Expression at Recurrence Is a Significant Adverse Prognostic Factor. Brain Sci. 2023, 13, 1477. https://doi.org/10.3390/brainsci13101477
Hoshimaru T, Nonoguchi N, Kosaka T, Furuse M, Kawabata S, Yagi R, Kurisu Y, Kashiwagi H, Kameda M, Takami T, et al. Actin Alpha 2, Smooth Muscle (ACTA2) Is Involved in the Migratory Potential of Malignant Gliomas, and Its Increased Expression at Recurrence Is a Significant Adverse Prognostic Factor. Brain Sciences. 2023; 13(10):1477. https://doi.org/10.3390/brainsci13101477
Chicago/Turabian StyleHoshimaru, Takumi, Naosuke Nonoguchi, Takuya Kosaka, Motomasa Furuse, Shinji Kawabata, Ryokichi Yagi, Yoshitaka Kurisu, Hideki Kashiwagi, Masahiro Kameda, Toshihiro Takami, and et al. 2023. "Actin Alpha 2, Smooth Muscle (ACTA2) Is Involved in the Migratory Potential of Malignant Gliomas, and Its Increased Expression at Recurrence Is a Significant Adverse Prognostic Factor" Brain Sciences 13, no. 10: 1477. https://doi.org/10.3390/brainsci13101477