
Differences and Changes in Cerebellar Functional Connectivity of Parkinson’s Patients with Visual Hallucinations
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Structure Magnetic Resonance Imaging Data Acquisition and Preprocessing
2.3. Functional Magnetic Resonance Imaging Data Acquisition and Preprocessing
2.4. Functional Connectivity Analysis
2.5. Feature Select
2.6. SVM Classification
3. Results
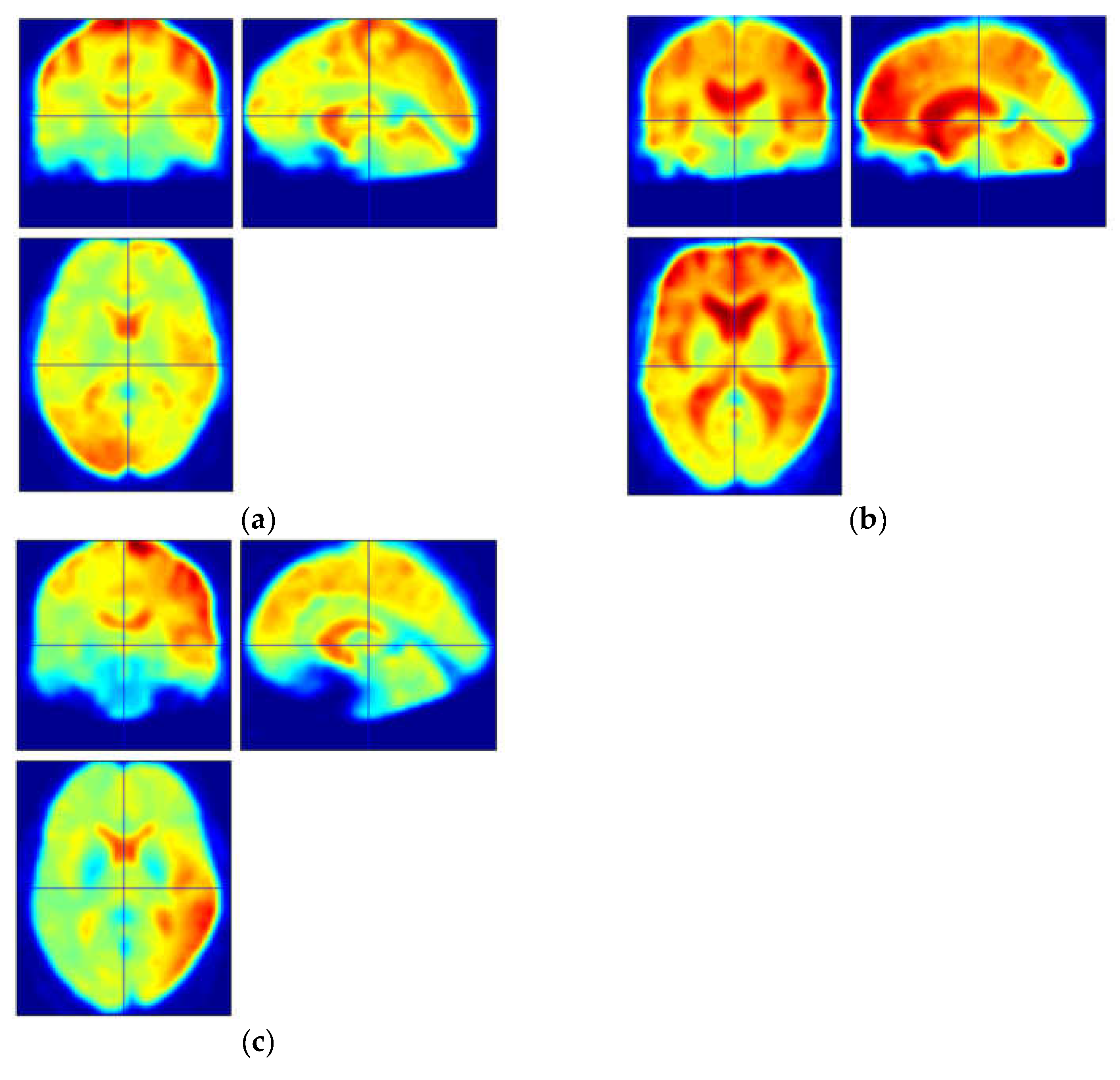
3.1. Functional Connectivity Changes of the Cerebellar
3.1.1. Functional Connectivity Changes of the Vermis Lobule I_II
3.1.2. Functional Connectivity Changes of the Vermis Lobule IV_V
3.2. Machine Learning Training Results
4. Discussion and Conclusions
4.1. Functional Connectivity Changes in the PD-H Patients
4.2. Functional Connectivity Changes in the PD-NH Patients
4.3. Functional Connectivity Changes in Classification Task
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sveinbjornsdottir, S. The clinical symptoms of Parkinson’s disease. J. Neurochem. 2016, 139 (Suppl. S1), 318–324. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, R.F. Non-motor symptoms in Parkinson’s disease. Park. Relat. Disord. 2016, 22 (Suppl. S1), S119–S122. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Martin, P.; Schapira, A.H.; Stocchi, F.; Sethi, K.; Odin, P.; MacPhee, G.; Brown, R.G.; Naidu, Y.; Clayton, L.; Abe, K.; et al. Prevalence of nonmotor symptoms in Parkinson’s disease in an international setting; study using nonmotor symptoms questionnaire in 545 patients. Mov. Disord. 2007, 22, 1623–1629. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, K.R.; Martinez-Martin, P.; Schapira, A.H.; Stocchi, F.; Sethi, K.; Odin, P.; Brown, R.G.; Koller, W.; Barone, P.; MacPhee, G.; et al. International multicenter pilot study of the first comprehensive self-completed nonmotor symptoms questionnaire for Parkinson’s disease: The NMSQuest study. Mov. Disord. 2006, 21, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Xu, W.; Liu, F.T.; Li, J.Q.; Cao, X.P.; Tan, L.; Wang, J.; Yu, J.T. Modifiable risk factors for cognitive impairment in Parkinson’s disease: A systematic review and meta-analysis of prospective cohort studies. Mov. Disord. 2019, 34, 876–883. [Google Scholar] [CrossRef] [PubMed]
- Dehsarvi, A. Classification of resting-state fMRI using evolutionary algorithms: Towards a brain imaging biomarker for Parkinson’s disease. arXiv 2018, arXiv:1910.05378. [Google Scholar]
- Engels, G.; Vlaar, A.; McCoy, B.; Scherder, E.; Douw, L. Dynamic Functional Connectivity and Symptoms of Parkinson’s Disease: A Resting-State fMRI Study. Front. Aging Neurosci. 2018, 10, 388. [Google Scholar] [CrossRef] [PubMed]
- Bejr-Kasem, H.; Pagonabarraga, J.; Martínez-Horta, S.; Sampedro, F.; Marín-Lahoz, J.; Horta-Barba, A.; Aracil-Bolaños, I.; Pérez-Pérez, J.; Botí, M.; Campolongo, A.; et al. Disruption of the default mode network and its intrinsic functional connectivity underlies minor hallucinations in Parkinson’s disease. Mov. Disord. 2019, 34, 78–86. [Google Scholar] [CrossRef]
- Holroyd, S.; Wooten, G.F. Preliminary FMRI evidence of visual system dysfunction in Parkinson’s disease patients with visual hallucinations. Neuropsychiatry Clin. Neurosci. 2006, 18, 402–404. [Google Scholar] [CrossRef]
- Yao, N.; Shek-Kwan Chang, R.; Cheung, C.; Pang, S.; Lau, K.K.; Suckling, J.; Rowe, J.B.; Yu, K.; Ka-Fung Mak, H.; Chua, S.E.; et al. The default mode network is disrupted in Parkinson’s disease with visual hallucinations. Hum. Brain Mapp. 2014, 35, 5658–5666. [Google Scholar] [CrossRef]
- Rapoport, M.; van Reekum, R.; Mayberg, H. The role of the cerebellum in cognition and behavior: A selective review. Neuropsychiatr. Clin. Neurosci. 2000, 12, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D.; Caplan, D. Cognition, emotion and the cerebellum. Brain 2006, 129 Pt 2, 290–292. [Google Scholar] [CrossRef] [PubMed]
- Balsters, J.H.; Whelan, C.D.; Robertson, I.H.; Ramnani, N. Cerebellum and cognition: Evidence for the encoding of higher order rules. Cereb. Cortex 2013, 23, 1433–1443. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Hu, J.; Ma, W.; Wang, D.; Yao, Q.; Shi, J. Altered baseline activity and connectivity associated with cognitive impairment following acute cerebellar infarction: A resting-state fMRI study. Neurosci. Lett. 2019, 692, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yao, Q.; Yu, M.; Xiao, C.; Fan, L.; Lin, X.; Zhu, D.; Tian, M.; Shi, J. Topological disruption of structural brain networks in patients with cognitive impairment following cerebellar infarction. Front. Neurol. 2019, 10, 759. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D.; Sherman, J.C. The cerebellar cognitive affective syndrome. Brain 1998, 121 Pt 4, 561–579. [Google Scholar] [CrossRef] [PubMed]
- Hoche, F.; Guell, X.; Vangel, M.G.; Sherman, J.C.; Schmahmann, J.D. The cerebellar cognitive affective/Schmahmann syndrome scale. Brain 2018, 141, 248–270. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, N.; van Baarsen, K.; van Zandvoort, M.; Robe, P.A. The cerebellar cognitive affective syndrome-a meta-analysis. Cerebellum 2019, 18, 941–950. [Google Scholar] [CrossRef]
- Sereno, M.I.; Diedrichsen, J.; Tachrount, M.; Testa-Silva, G.; d’Arceuil, H.; De Zeeuw, C. The human cerebellum has almost 80% of the surface area of the neocortex. Proc. Natl. Acad. Sci. USA 2020, 117, 19538–19543. [Google Scholar] [CrossRef]
- Buckner, R.L.; Krienen, F.M.; Castellanos, A.; Diaz, J.C.; Yeo, B.T. The organization of the human cerebellum estimated by intrinsic functional connectivity. J. Neurophysiol. 2011, 106, 2322–2345. [Google Scholar] [CrossRef]
- Toniolo, S.; Serra, L.; Olivito, G.; Caltagirone, C.; Mercuri, N.B.; Marra, C.; Cercignani, M.; Bozzali, M. Cerebellar white matter disruption in Alzheimer’s disease patients: A diffusion tensor imaging study. J. Alzheimer’s Dis. 2020, 74, 615–624. [Google Scholar] [CrossRef]
- Zhang, L.; Ni, H.; Yu, Z.; Wang, J.; Qin, J.; Hou, F.; Yang, A.; Alzheimer’s Disease Neuroimaging Initiative (ADNI). Investigation on the alteration of brain functional network and its role in the identification of mild cognitive impairment. Front. Neurosci. 2020, 14, 558434. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Liao, W.; Watson, D.R.; Shi, Y.; Yuan, Y.; Cohen, A.D.; Xie, C.; Wang, Y.; Yue, C.; Teng, Y.; et al. Mapping the altered patterns of cerebellar resting-state function in longitudinal amnestic mild cognitive impairment patients. J. Alzheimers Dis. 2011, 23, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Sveljo, O.; Culic, M.; Koprivsek, K.; Lucic, M. The functional neuroimaging evidence of cerebellar involvement in the simple cognitive task. Brain Imaging Behav. 2014, 8, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Küper, M.; Kaschani, P.; Thürling, M.; Stefanescu, M.R.; Burciu, R.G.; Göricke, S.; Maderwald, S.; Ladd, M.E.; Hautzel, H.; Timmann, D. Cerebellar fMRI activation increases with increasing working memory demands. Cerebellum 2016, 15, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D. Cerebellum in Alzheimer’s disease and frontotemporal dementia: Not a silent bystander. Brain 2016, 139 Pt 5, 1314–1318. [Google Scholar] [CrossRef]
- Stoodley, C.J.; Schmahmann, J.D. Functional topography in the human cerebellum: A meta analysis of neuroimaging studies. Neuroimage 2009, 44, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, A.A.; Miall, R.C.; Ivry, R.B. The cerebellum: Adaptive prediction for movement and cognition. Trends Cogn. Sci. 2017, 21, 313–332. [Google Scholar] [CrossRef]
- Guell, X.; Gabrieli, J.D.E.; Schmahmann, J.D. Triple representation of language, working memory, social and emotion processing in the cerebellum: Convergent evidence from task and seed-based restingstate fMRI analyses in a single large cohort. Neuroimage 2018, 172, 437–449. [Google Scholar] [CrossRef]
- Shipman, M.L.; Green, J.T. Cerebellum and cognition: Does the rodent cerebellum participate in cognitive functions? Neurobiol. Learn. Mem. 2019, 170, 106996. [Google Scholar] [CrossRef]
- Gao, Z.; Liu, X.; Zhang, D.; Liu, M.; Hao, N. The indispensable role of the cerebellum in visual divergent thinking. Sci. Rep. 2020, 10, 16552. [Google Scholar] [CrossRef] [PubMed]
- Lawn, T.; Ffytche, D. Cerebellar Correlates of Visual Hallucinations in Parkinson’s Disease and Charles Bonnet Syndrome. Cortex 2020, 135, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Nasreddine, Z.S.; Phillips, N.A.; Bédirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H. The Montreal Cognitive Assessment, MoCA: A Brief Screening Tool For Mild Cognitive Impairment. J. Am. Geriatr. Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Folstein, M.F.; Folstein, S.E.; Mchugh, P.R. ”Mini-mental state”: A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.G.; Wang, X.D.; Zuo, X.N.; Zang, Y.F. DPABI: Data processing and analysis for (Resting-State) brain imaging. Neuroinformatics 2016, 14, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Hepp, D.H.; Foncke, E.M.J.; Dubbelink, K.T.E.O.; van de Berg, W.D.J.; Berendse, H.W.; Schoonheim, M.M. Loss of functional connectivity in patients with parkinson disease and visual hallucinations. Radiology 2017, 285, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E. The brain’s default mode network. Annu. Rev. Neurosci. 2015, 38, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Davey, C.G.; Pujol, J.; Harrison, B.J. Mapping the self in the brain’s default mode network. Neuroimage 2016, 132, 390–397. [Google Scholar] [CrossRef]
- Davey, C.G.; Harrison, B.J. The brain’s center of gravity: How the default mode network helps us to understand the self. World Psychiatr. 2018, 17, 278–279. [Google Scholar] [CrossRef]
- Habas, C.; Kamdar, N.; Nguyen, D.; Prater, K.; Beckmann, C.F.; Menon, V.; Greicius, M.D. Distinct cerebellar contributions to intrinsic connectivity networks. J. Neurosci. 2009, 29, 8586–8594. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeong, G.W.; Kang, H.K.; Lee, M.S.; Park, T.J. The Effect of Memory Load on Maintenance in Face and Spatial Working Memory: An Event-Related fMRI Study. Korean J. Cogn. Sci. 2010, 21, 359–386. [Google Scholar]
- Tino, P. Structural and Functional Brain Patterns of Non-Motor Syndromes in Parkinson’s Disease. Front. Neurol. 2018, 9, 138. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC | PD-H | PD-NH | p | P1 | P2 | P3 | |
|---|---|---|---|---|---|---|---|
| Age (years) | 59.12 ± 6.289 | 63.24 ± 6.582 | 59.00 ± 6.338 | 0.091 | |||
| Education (years) | 8.788 ± 3.287 | 8.294 ± 4.832 | 7.923 ± 4.609 | 0.814 | |||
| Sex (male) | 46.2% | 41.2% | 38.5 | 0.285 | |||
| Moca (scores) | 25.38 ± 3.601 | 19.76 ± 4.191 | 21.85 ± 5.398 | 0.000 | 0.017 | 0.000 | 0.599 |
| Course (years) | 2.294 ± 1.105 | 1.577 ± 1.718 | - | - | 0.176 |
| PD-H vs. HC | PD-H vs. PD-NH | ||
|---|---|---|---|
| Node-a | Node-b | Node-a | Node-b |
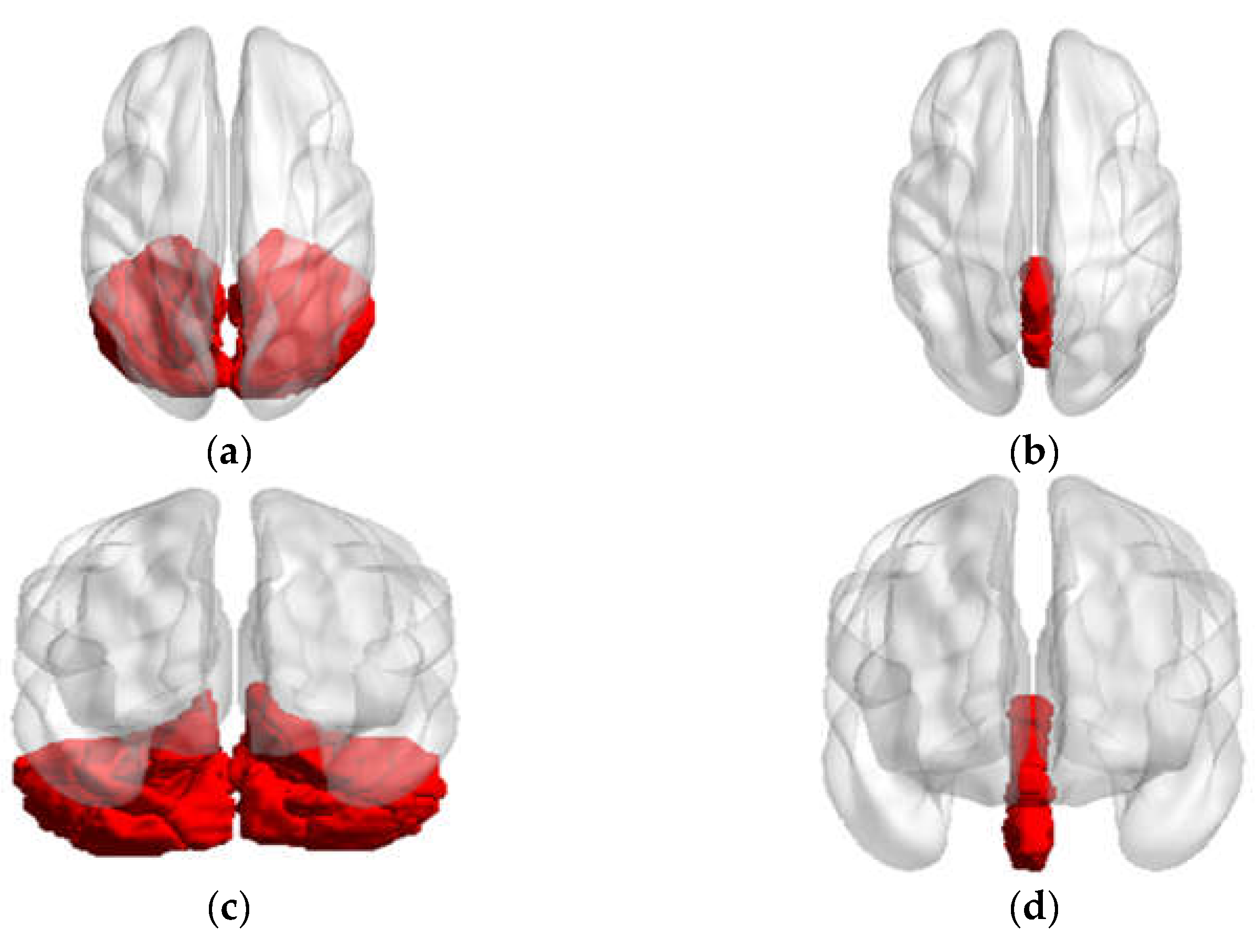
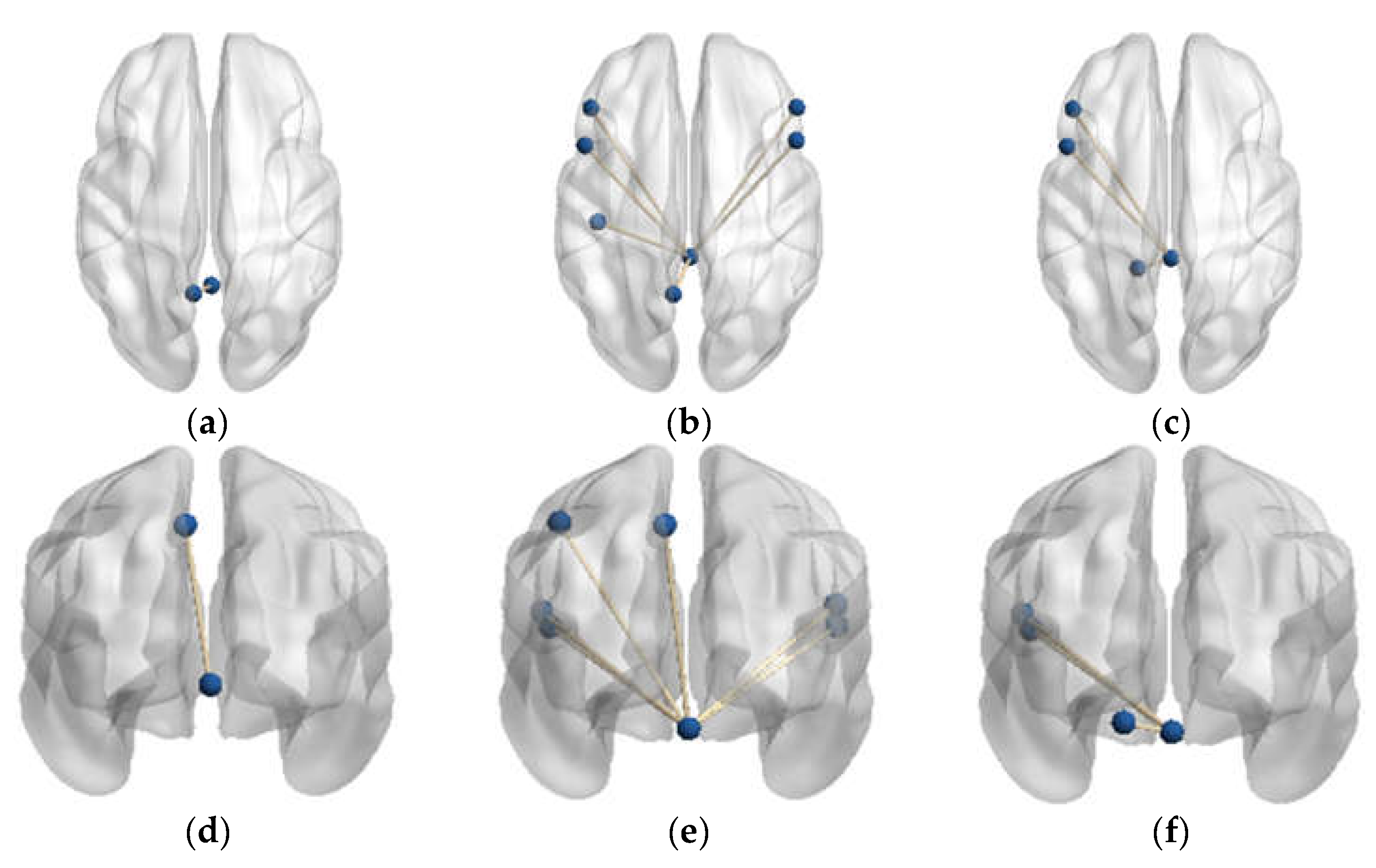
| Vermis_4_5 | Precuneus_L | Vermis_1_2 | Frontal_Inf_Oper_L Frontal_Inf_Oper_R Frontal_Inf_Tri_L Frontal_Inf_Tri_R Postcentral_L Parietal_Inf_L Precuneus_L |
| PD-NH vs. HC | |||
| Vermis_1_2 | Frontal_Inf_Oper_L Frontal_Inf_Tri_L Cerebelum_4_5_L | ||
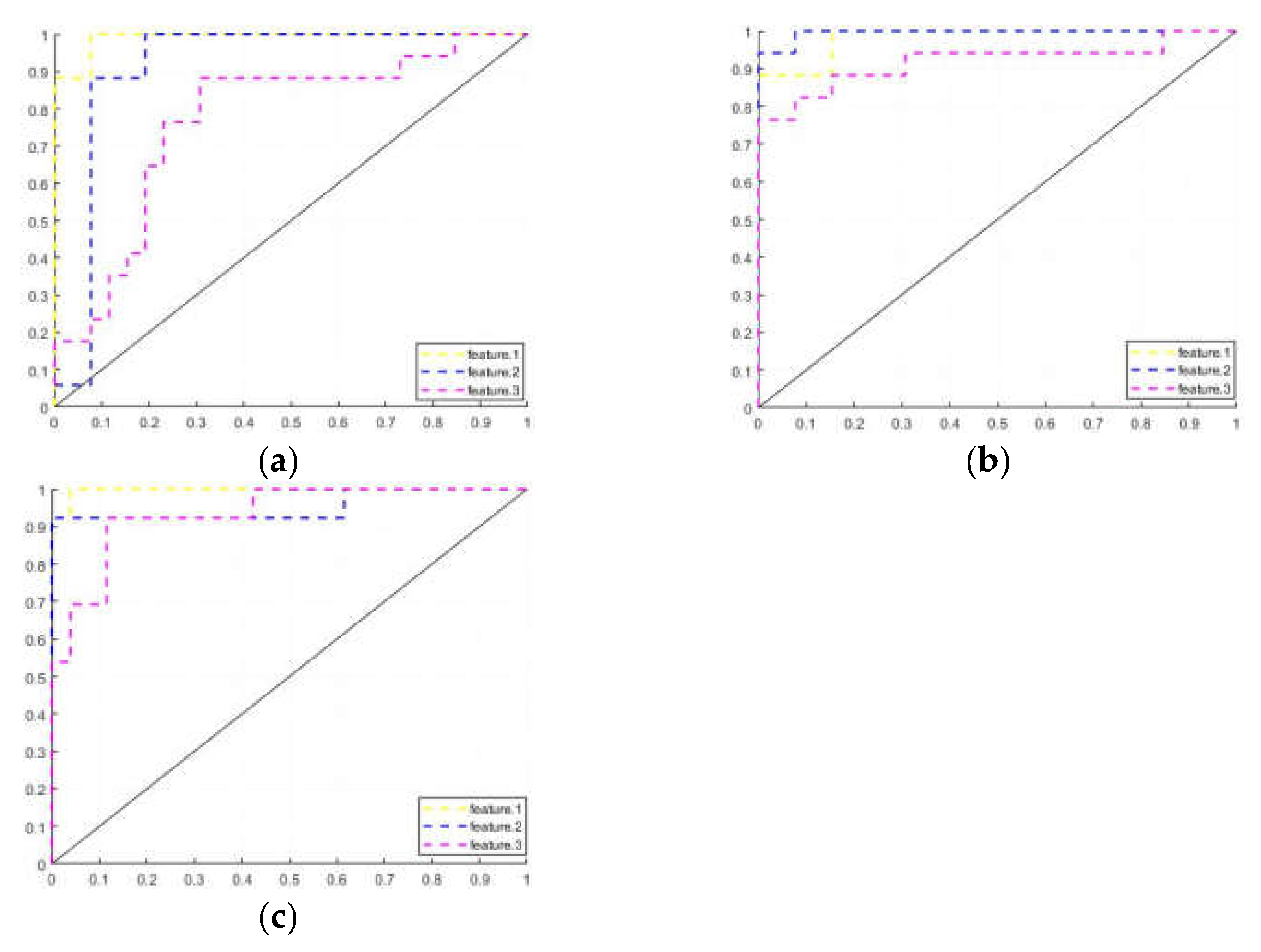
| ACC | SEN | SPE | AUC | |
|---|---|---|---|---|
| Feature 1 | ||||
| PD-H:HC | 93.02 [80.94,98.54] | 88.24 [63.56,98.54] | 96.15 [80.36,99.90] | 0.9910 |
| PD-H:PD-NH | 86.67 [69.28,96.24] | 88.24 [63.56,98.54] | 84.62 [54.55,98.08] | 0.9819 |
| PD-NH:HC | 92.31 [79.13,98.38] | 84.62 [54.55,98.08] | 96.15 [80.36,99.90] | 0.9970 |
| Ferutre 2 | ||||
| PD-H:HC | 88.37 [74.92,96.11] | 82.35 [56.57,92.30] | 92.31 [74.87,99.05] | 0.9140 |
| PD-H:PD-NH | 93.33 [77.93,99.18] | 94.12 [71.31,99.85] | 92.31 [63.97,99.81] | 0.9955 |
| PD-NH:HC | 89.74 [75.78,97.13] | 92.31 [63.97,99.81] | 88.46 [69.85,97.55] | 0.9527 |
| Feature 3 | ||||
| PD-H:HC | 74.42 [58.83,86.48] | 70.59 [44.04,84.69] | 76.92 [56.35,91.03] | 0.7715 |
| PD-H:PD-NH | 80.00 [61.43,92.29] | 88.24 [63.56,98.54] | 69.23 [38.57,90.91] | 0.9186 |
| PD-NH:HC | 89.47 [75.78,97.13] | 92.31 [63.97,99.81] | 88.46 [69.85,97.55] | 0.9349 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, L.; Liu, C.; Cao, Y.; Shi, J.; Yin, K.; Liu, W. Differences and Changes in Cerebellar Functional Connectivity of Parkinson’s Patients with Visual Hallucinations. Brain Sci. 2023, 13, 1458. https://doi.org/10.3390/brainsci13101458
Qu L, Liu C, Cao Y, Shi J, Yin K, Liu W. Differences and Changes in Cerebellar Functional Connectivity of Parkinson’s Patients with Visual Hallucinations. Brain Sciences. 2023; 13(10):1458. https://doi.org/10.3390/brainsci13101458
Chicago/Turabian StyleQu, Liangcheng, Chuan Liu, Yiting Cao, Jingping Shi, Kuiying Yin, and Weiguo Liu. 2023. "Differences and Changes in Cerebellar Functional Connectivity of Parkinson’s Patients with Visual Hallucinations" Brain Sciences 13, no. 10: 1458. https://doi.org/10.3390/brainsci13101458