
Dissociating Language and Thought in Human Reasoning †
 ,
,  and
and
Abstract
:1. Dissociating Language and Thought in Human Reasoning
2. Methods
2.1. Participants
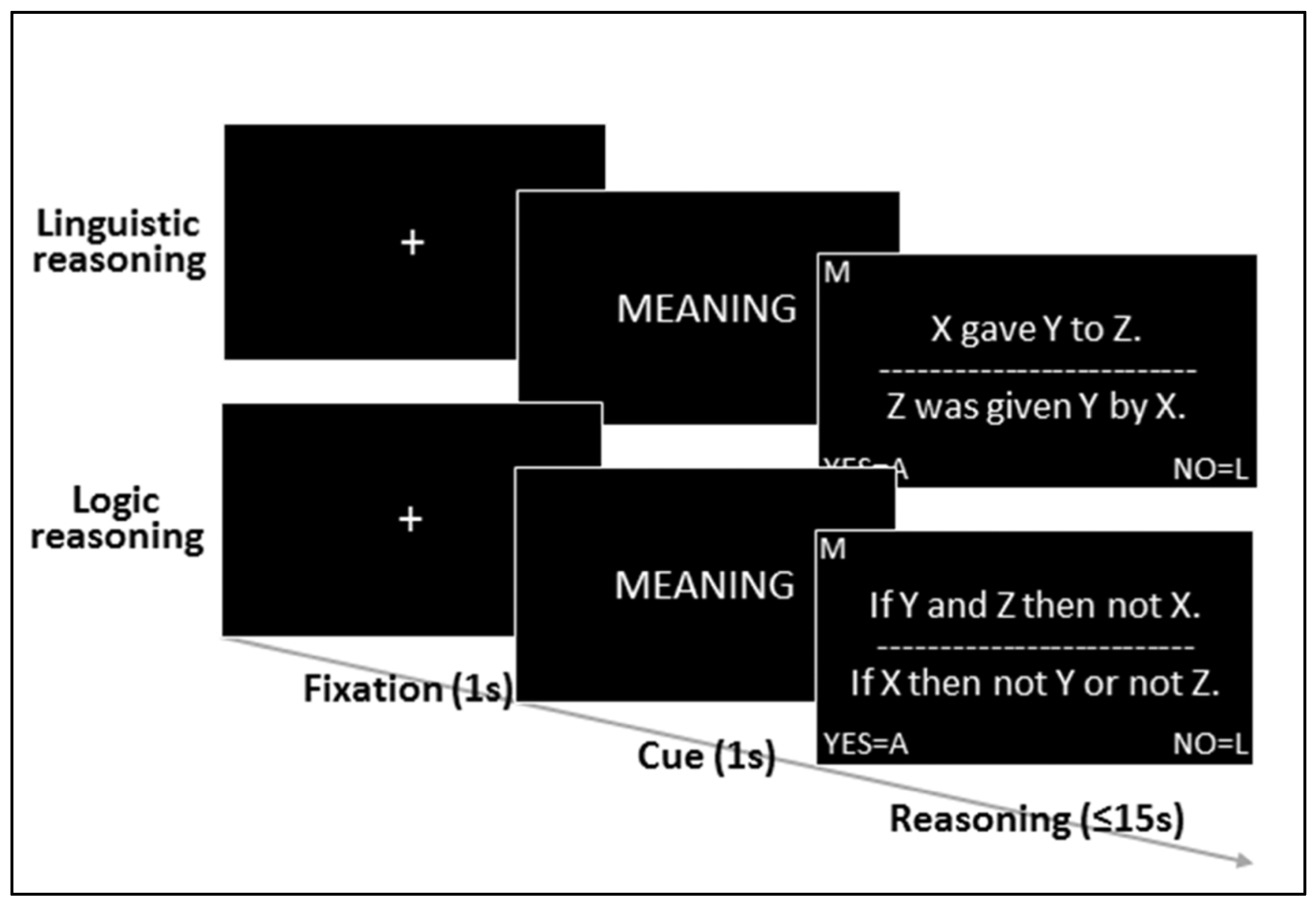
2.2. Task and Stimuli
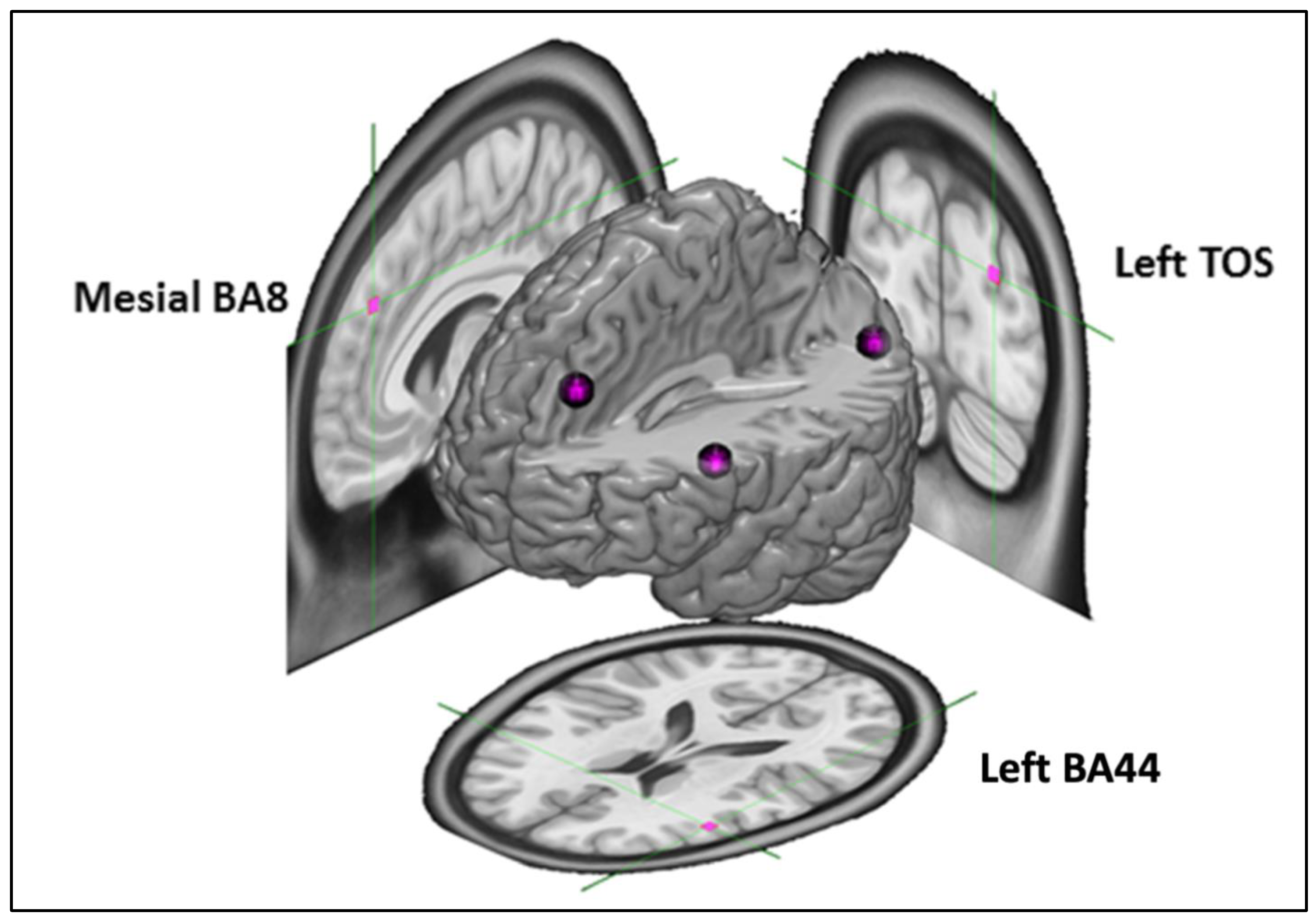
2.3. Targets

2.4. Experimental Sessions
2.5. Continuous Theta Burst Stimulation
2.6. Analysis
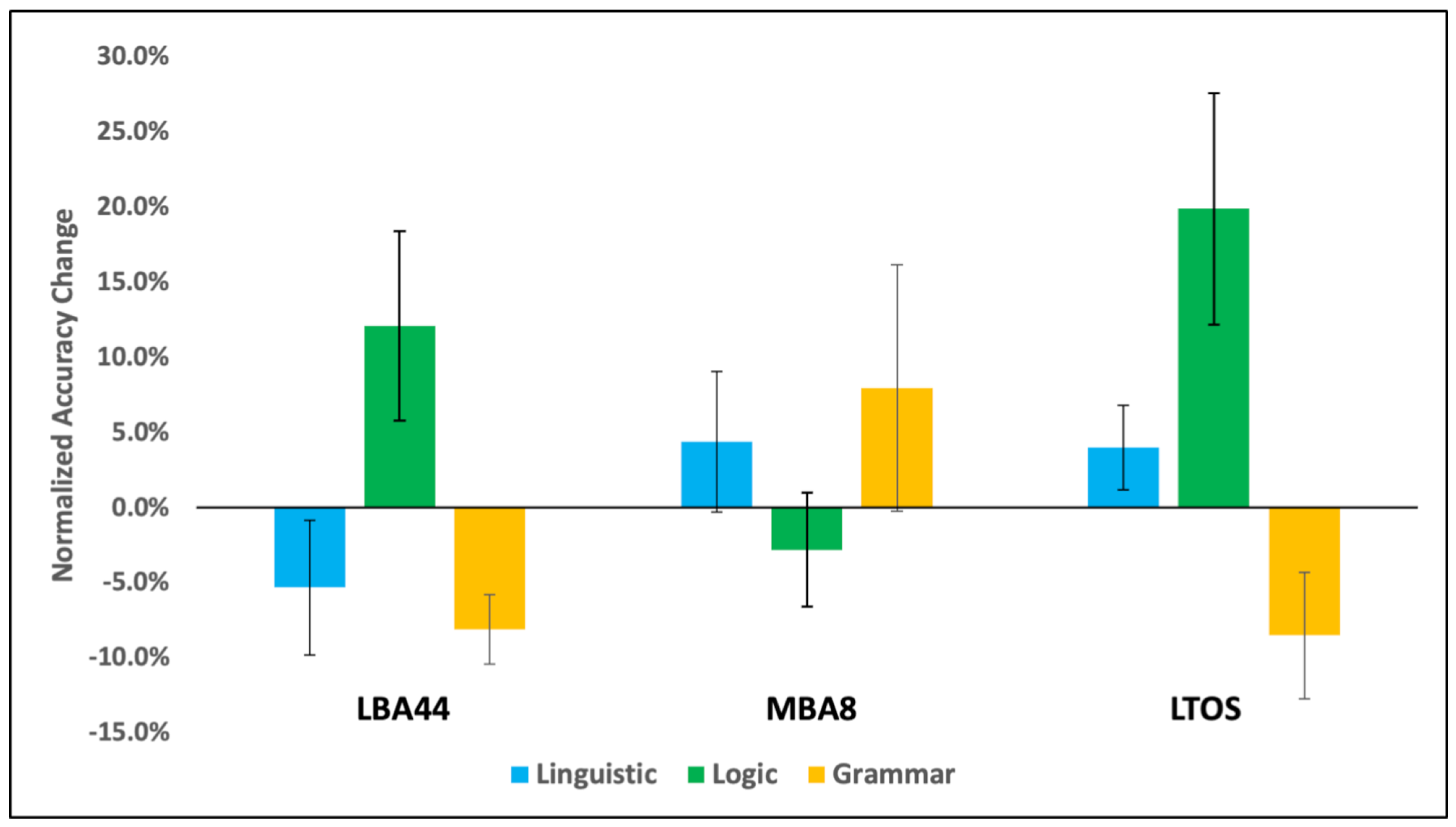
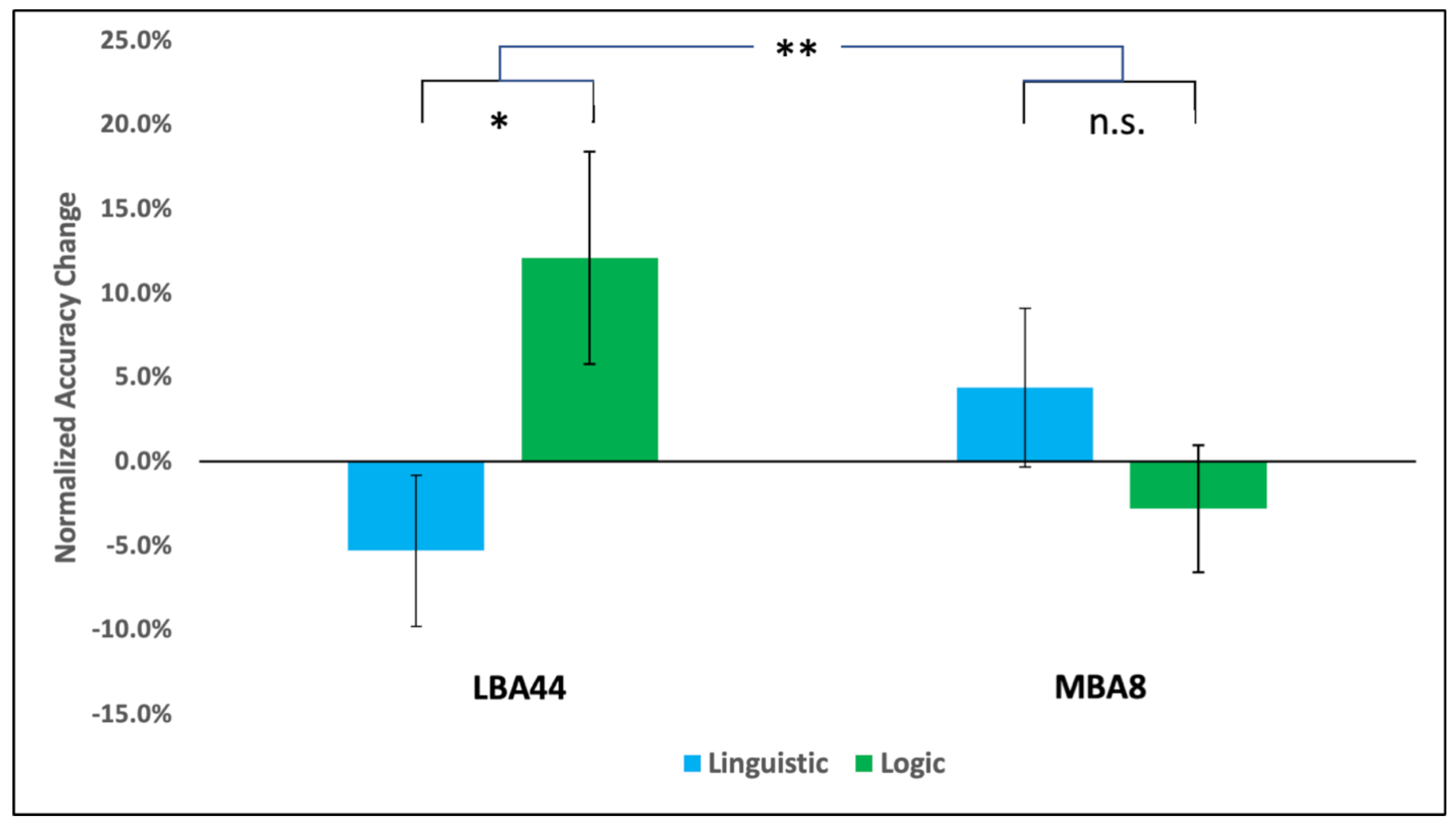
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fodor, J.A. The Language of Thought; Harvard University Press: Cambridge, MA, USA, 1975; Volume 5. [Google Scholar]
- Pinker, S. The Language Instinct: How the Mind Creates Language, 1st ed.; Harper Perennial Modern Classics: New York, NY, USA, 2007; ISBN 978-0-06-203252-2. [Google Scholar]
- Sapir, E. The Status of Linguistics as a Science. Language 1929, 5, 207–214. [Google Scholar] [CrossRef]
- Vygotsky, L.S. Thought and Language; MIT Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Whorf, B.L. Science and Linguistics; Bobbs-Merrill: Indianapolis, IN, USA, 1940. [Google Scholar]
- Carruthers, P.; Boucher, J. Language and Thought: Interdisciplinary Themes; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Sapir, E. Conceptual Categories in Primitive Languages. Science 1931, 74, 578. [Google Scholar]
- Von Humboldt, W.; von Humboldt, W.F. Humboldt: ’On Language’: On the Diversity of Human Language Construction and Its Influence on the Mental Development of the Human Species; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Carruthers, P. The Cognitive Functions of Language. Behav. Brain Sci. 2002, 25, 657–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locke, J. The Works of John Locke: In Nine Volumes; C. and J. Rivington: London, UK, 1824; Volume 7. [Google Scholar]
- Monti, M.M. The Role of Language in Structure-Dependent Cognition. In Neural Mechanisms of Language; Moody, M., Ed.; Springer: New York, NY, USA, 2015. [Google Scholar]
- Pinker, S. Language Learnability and Language Development: With New Commentary by the Author; Harvard University Press: Cambridge, MA, USA, 2009; Volume 7. [Google Scholar]
- Soja, N.N.; Carey, S.; Spelke, E.S. Ontological Categories Guide Young Children’s Inductions of Word Meaning: Object Terms and Substance Terms. Cognition 1991, 38, 179–211. [Google Scholar] [CrossRef] [PubMed]
- Regier, T.; Kay, P. Language, Thought, and Color: Whorf Was Half Right. Trends Cogn. Sci. 2009, 13, 439–446. [Google Scholar] [CrossRef]
- Gibson, E.; Futrell, R.; Jara-Ettinger, J.; Mahowald, K.; Bergen, L.; Ratnasingam, S.; Gibson, M.; Piantadosi, S.T.; Conway, B.R. Color Naming across Languages Reflects Color Use. Proc. Natl. Acad. Sci. USA 2017, 114, 10785–10790. [Google Scholar] [CrossRef] [Green Version]
- De Cruz, H. An Enhanced Argument for Innate Elementary Geometric Knowledge and Its Philosophical Implications. In New Perspectives on Mathematical Practices; World Scientific: Brussels, Belgium, 2008; pp. 185–206. [Google Scholar]
- Hermer-Vazquez, L.; Spelke, E.S.; Katsnelson, A.S. Sources of Flexibility in Human Cognition: Dual-Task Studies of Space and Language. Cognit. Psychol. 1999, 39, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Spelke, E.S.; Tsivkin, S. Language and Number: A Bilingual Training Study. Cognition 2001, 78, 45–88. [Google Scholar] [CrossRef]
- Varley, R.A.; Klessinger, N.J.C.; Romanowski, C.A.J.; Siegal, M. Agrammatic but Numerate. Proc. Natl. Acad. Sci. USA 2005, 102, 3519–3524. [Google Scholar] [CrossRef] [Green Version]
- Papafragou, A.; Massey, C.; Gleitman, L. Shake, Rattle, ‘n’ Roll: The Representation of Motion in Language and Cognition. Cognition 2002, 84, 189–219. [Google Scholar] [CrossRef] [Green Version]
- Peretz, I.; Vuvan, D.; Lagrois, M.-É.; Armony, J.L. Neural Overlap in Processing Music and Speech. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140090. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.N.; Rosenberg, M.H.; Bufford, C.A.; Stephens, D.; Lysy, A.; Monti, M.M. The Language of Music: Common Neural Codes for Structured Sequences in Music and Natural Language. Brain Lang. 2018, 185, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Papafragou, A.; Cassidy, K.; Gleitman, L. When We Think about Thinking: The Acquisition of Belief Verbs. Cognition 2007, 105, 125–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Villiers, J. The Interface of Language and Theory of Mind. Lingua 2007, 117, 1858–1878. [Google Scholar] [CrossRef] [Green Version]
- Friederici, A.D. Processing Local Transitions versus Long-Distance Syntactic Hierarchies. Trends Cogn. Sci. 2004, 8, 245–247. [Google Scholar] [CrossRef]
- Friederici, A.D. The Neuroanatomical Pathway Model of Language. In Neurobiology of Language; Elsevier: Amsterdam, The Netherlands, 2016; pp. 349–356. ISBN 978-0-12-407794-2. [Google Scholar]
- Grodzinsky, Y.; Santi, A. The Battle for Broca’s Region. Trends Cogn. Sci. 2008, 12, 474–480. [Google Scholar] [CrossRef]
- Bornkessel, I.; Zysset, S.; Friederici, A.D.; von Cramon, D.Y.; Schlesewsky, M. Who Did What to Whom? The Neural Basis of Argument Hierarchies during Language Comprehension. NeuroImage 2005, 26, 221–233. [Google Scholar] [CrossRef]
- Ben-Shachar, M.; Hendler, T.; Kahn, I.; Ben-Bashat, D.; Grodzinsky, Y. The Neural Reality of Syntactic Transformations Evidence From Functional Magnetic Resonance Imaging. Psychol. Sci. 2003, 14, 433–440. [Google Scholar] [CrossRef]
- Fadiga, L.; Craighero, L.; D’Ausilio, A. Broca’s Area in Language, Action, and Music. Ann. N. Y. Acad. Sci. 2009, 1169, 448–458. [Google Scholar] [CrossRef]
- Fitch, W.T.; Martins, M.D. Hierarchical Processing in Music, Language, and Action: Lashley Revisited. Ann. N. Y. Acad. Sci. 2014, 1316, 87–104. [Google Scholar] [CrossRef] [Green Version]
- Tettamanti, M.; Weniger, D. Broca’s Area: A Supramodal Hierarchical Processor? Cortex 2006, 42, 491–494. [Google Scholar] [CrossRef] [PubMed]
- Clark, H.H. Linguistic Processes in Deductive Reasoning. Psychol. Rev. 1969, 76, 387–404. [Google Scholar] [CrossRef] [Green Version]
- Krumnack, A.; Bucher, L.; Nejasmic, J.; Nebel, B.; Knauff, M. A Model for Relational Reasoning as Verbal Reasoning. Cogn. Syst. Res. 2011, 12, 377–392. [Google Scholar] [CrossRef]
- Montague, R. Formal Philosophy: Selected Papers of Richard Montague; Edited and with an Introduction by Richmond H. Thomason; Yale University Press: New Haven, CT, USA, 1974. [Google Scholar]
- Partee, B.H.; Hendriks, H.L.W. Montague Grammar. In Handbook of Logic and Language; Elsevier: Amsterdam, The Netherlands, 1997; pp. 5–91. ISBN 978-0-444-81714-3. [Google Scholar]
- Rips, L.J. The Psychology of Proof: Deductive Reasoning in Human Thinking; MIT Press: Cambridge, MA, USA, 1994; ISBN 978-0-262-18153-2. [Google Scholar]
- Bonatti, L.L.; Cherubini, P.; Reverberi, C. Nothing New under the Sun, or the Moon, or Both. Front. Hum. Neurosci. 2015, 9, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coetzee, J.P.; Monti, M.M. At the Core of Reasoning: Dissociating Deductive and Non-Deductive Load. Hum. Brain Mapp. 2018, 39, 1850–1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fangmeier, T.; Knauff, M.; Ruff, C.C.; Sloutsky, V. FMRI Evidence for a Three-Stage Model of Deductive Reasoning. J. Cogn. Neurosci. 2006, 18, 320–334. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, A.A.; Srikant, S.; Sueoka, Y.; Kean, H.H.; Dhamala, R.; O’Reilly, U.-M.; Bers, M.U.; Fedorenko, E. Comprehension of Computer Code Relies Primarily on Domain-General Executive Brain Regions. eLife 2020, 9, e58906. [Google Scholar] [CrossRef]
- Monti, M.M.; Osherson, D.N. Logic, Language and the Brain. Brain Res. 2012, 1428, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Prado, J. The Relationship between Deductive Reasoning and the Syntax of Language in Broca’s Area: A Review of the Neuroimaging Literature. L’Année Psychol. 2018, 118, 289–315. [Google Scholar] [CrossRef] [Green Version]
- Prado, J.; Chadha, A.; Booth, J.R. The Brain Network for Deductive Reasoning: A Quantitative Meta-Analysis of 28 Neuroimaging Studies. J. Cogn. Neurosci. 2011, 23, 3483–3497. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, M.; Zou, F.; Wu, X.; Wang, Y. Deductive-reasoning Brain Networks: A Coordinate-based Meta-analysis of the Neural Signatures in Deductive Reasoning. Brain Behav. 2020, 10, e01853. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, C.; Cherubini, P.; Rapisarda, A.; Rigamonti, E.; Caltagirone, C.; Frackowiak, R.S.J.J.; Macaluso, E.; Paulesu, E. Neural Basis of Generation of Conclusions in Elementary Deduction. NeuroImage 2007, 38, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, C.; Cherubini, P.; Frackowiak, R.S.J.; Caltagirone, C.; Paulesu, E.; Macaluso, E. Conditional and Syllogistic Deductive Tasks Dissociate Functionally during Premise Integration. Hum. Brain Mapp. 2010, 31, 1430–1445. [Google Scholar] [CrossRef]
- Goel, V.; Gold, B.; Kapur, S.; Houle, S. The Seats of Reason? An Imaging Study of Deductive and Inductive Reasoning. NeuroReport 1997, 8, 1305–1310. [Google Scholar] [CrossRef]
- Goel, V.; Gold, B.; Kapur, S.; Houle, S. Neuroanatomical Correlates of Human Reasoning. J. Cogn. Neurosci. 1998, 10, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Goel, V.; Dolan, R.J. Differential Involvement of Left Prefrontal Cortexin Inductive and Deductive Reasoning. Cognition 2004, 93, B109–B121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prado, J.; Der Henst, J.-B.V.; Noveck, I.A. Recomposing a Fragmented Literature: How Conditional and Relational Arguments Engage Different Neural Systems for Deductive Reasoning. NeuroImage 2010, 51, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.M.; Parsons, L.M.; Osherson, D.N. The Boundaries of Language and Thought in Deductive Inference. Proc. Natl. Acad. Sci. USA 2009, 106, 12554–12559. [Google Scholar] [CrossRef] [Green Version]
- Monti, M.M.; Osherson, D.N.; Martinez, M.J.; Parsons, L.M. Functional Neuroanatomy of Deductive Inference: A Language-Independent Distributed Network. NeuroImage 2007, 37, 1005–1016. [Google Scholar] [CrossRef]
- Rodriguez-Moreno, D.; Hirsch, J. The Dynamics of Deductive Reasoning: An FMRI Investigation. Neuropsychologia 2009, 47, 949–961. [Google Scholar] [CrossRef]
- Canessa, N.; Gorini, A.; Cappa, S.F.; Piattelli-Palmarini, M.; Danna, M.; Fazio, F.; Perani, D. The Effect of Social Content on Deductive Reasoning: An FMRI Study. Hum. Brain Mapp. 2005, 26, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Goel, V.; Dolan, R.J. Functional Neuroanatomy of Three-Term Relational Reasoning. Neuropsychologia 2001, 39, 901–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knauff, M.; Mulack, T.; Kassubek, J.; Salih, H.R.; Greenlee, M.W. Spatial Imagery in Deductive Reasoning: A Functional MRI Study. Cogn. Brain Res. 2002, 13, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noveck, I.A.; Goel, V.; Smith, K.W. The Neural Basis of Conditional Reasoning with Arbitrary Content. Cortex 2004, 40, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Parsons, L.M.; Osherson, D. New Evidence for Distinct Right and Left Brain Systems for Deductive versus Probabilistic Reasoning. Cereb. Cortex 2001, 11, 954–965. [Google Scholar] [CrossRef]
- Prado, J.; Noveck, I.A. Overcoming Perceptual Features in Logical Reasoning: A Parametric Functional Magnetic Resonance Imaging Study. J. Cogn. Neurosci. 2007, 19, 642–657. [Google Scholar] [CrossRef]
- Polk, T.A.; Newell, A. Deduction as Verbal Reasoning. Psychol. Rev. 1995, 102, 533–566. [Google Scholar] [CrossRef]
- Evans, J.S.B.T. Heuristic and Analytic Processes in Reasoning. Br. J. Psychol. 1984, 75, 451–468. [Google Scholar] [CrossRef]
- Bestmann, S.; Feredoes, E. Combined Neurostimulation and Neuroimaging in Cognitive Neuroscience: Past, Present, and Future. Ann. N. Y. Acad. Sci. 2013, 1296, 11–30. [Google Scholar] [CrossRef]
- Silvanto, J.; Pascual-Leone, A. Why the Assessment of Causality in Brain–Behavior Relations Requires Brain Stimulation. J. Cogn. Neurosci. 2012, 24, 775–777. [Google Scholar] [CrossRef]
- Huang, Y.-Z.; Edwards, M.J.; Rounis, E.; Bhatia, K.P.; Rothwell, J.C. Theta Burst Stimulation of the Human Motor Cortex. Neuron 2005, 45, 201–206. [Google Scholar] [CrossRef]
- Ojala, K.E.; Staib, M.; Gerster, S.; Ruff, C.C.; Bach, D.R. Inhibiting Human Aversive Memory by Transcranial Theta-Burst Stimulation to the Primary Sensory Cortex. Biol. Psychiatry 2022, 92, 149–157. [Google Scholar] [CrossRef]
- Sandrini, M.; Umiltà, C.; Rusconi, E. The Use of Transcranial Magnetic Stimulation in Cognitive Neuroscience: A New Synthesis of Methodological Issues. Neurosci. Biobehav. Rev. 2011, 35, 516–536. [Google Scholar] [CrossRef]
- Suppa, A.; Huang, Y.-Z.; Funke, K.; Ridding, M.C.; Cheeran, B.; Di Lazzaro, V.; Ziemann, U.; Rothwell, J.C. Ten Years of Theta Burst Stimulation in Humans: Established Knowledge, Unknowns and Prospects. Brain Stimulat. 2016, 9, 323–335. [Google Scholar] [CrossRef]
- Cattaneo, Z.; Devlin, J.T.; Vecchi, T.; Silvanto, J. Dissociable Neural Representations of Grammatical Gender in Broca’s Area Investigated by the Combination of Satiation and TMS. NeuroImage 2009, 47, 700–704. [Google Scholar] [CrossRef]
- Kuhnke, P.; Meyer, L.; Friederici, A.D.; Hartwigsen, G. Left Posterior Inferior Frontal Gyrus Is Causally Involved in Reordering during Sentence Processing. NeuroImage 2017, 148, 254–263. [Google Scholar] [CrossRef]
- Sakai, K.L.; Noguchi, Y.; Takeuchi, T.; Watanabe, E. Selective Priming of Syntactic Processing by Event-Related Transcranial Magnetic Stimulation of Broca’s Area. Neuron 2002, 35, 1177–1182. [Google Scholar] [CrossRef] [Green Version]
- Ragni, M.; Franzmeier, I.; Maier, S.; Knauff, M. Uncertain Relational Reasoning in the Parietal Cortex. Brain Cogn. 2016, 104, 72–81. [Google Scholar] [CrossRef]
- Tsujii, T.; Sakatani, K.; Masuda, S.; Akiyama, T.; Watanabe, S. Evaluating the Roles of the Inferior Frontal Gyrus and Superior Parietal Lobule in Deductive Reasoning: An RTMS Study. NeuroImage 2011, 58, 640–646. [Google Scholar] [CrossRef]
- Tsujii, T.; Watanabe, S. Neural Correlates of Belief-Bias Reasoning under Time Pressure: A near-Infrared Spectroscopy Study. NeuroImage 2010, 50, 1320–1326. [Google Scholar] [CrossRef]
- Bettencourt, K.C.; Xu, Y. The Role of Transverse Occipital Sulcus in Scene Perception and Its Relationship to Object Individuation in Inferior Intraparietal Sulcus. J. Cogn. Neurosci. 2013, 25, 1711–1722. [Google Scholar] [CrossRef]
- Ganaden, R.E.; Mullin, C.R.; Steeves, J.K.E. Transcranial Magnetic Stimulation to the Transverse Occipital Sulcus Affects Scene but Not Object Processing. J. Cogn. Neurosci. 2013, 25, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Welbourne, L.E.; Jonnalagadda, A.; Giesbrecht, B.; Eckstein, M.P. The Transverse Occipital Sulcus and Intraparietal Sulcus Show Neural Selectivity to Object-Scene Size Relationships. Commun. Biol. 2021, 4, 768. [Google Scholar] [CrossRef]
- Goel, V. Anatomy of Deductive Reasoning. Trends Cogn. Sci. 2007, 11, 435–441. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A Flexible Statistical Power Analysis Program for the Social, Behavioral, and Biomedical Sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Alamia, A.; Solopchuk, O.; D’Ausilio, A.; Van Bever, V.; Fadiga, L.; Olivier, E.; Zénon, A. Disruption of Broca’s Area Alters Higher-Order Chunking Processing during Perceptual Sequence Learning. J. Cogn. Neurosci. 2016, 28, 402–417. [Google Scholar] [CrossRef]
- Clerget, E.; Badets, A.; Duqué, J.; Olivier, E. Role of Broca’s Area in Motor Sequence Programming: A Ctbs Study. NeuroReport 2011, 22, 965–969. [Google Scholar] [CrossRef]
- Clerget, E.; Poncin, W.; Fadiga, L.; Olivier, E. Role of Broca’s Area in Implicit Motor Skill Learning: Evidence from Continuous Theta-Burst Magnetic Stimulation. J. Cogn. Neurosci. 2012, 24, 80–92. [Google Scholar] [CrossRef]
- Clerget, E.; Andres, M.; Olivier, E. Deficit in Complex Sequence Processing after a Virtual Lesion of Left BA45. PLoS ONE 2013, 8, e63722. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A. Safety, Ethical Considerations, and Application Guidelines for the Use of Transcranial Magnetic Stimulation in Clinical Practice and Research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [Green Version]
- Goodstein, R.L. Boolean Algebra; Dover Books on Mathematics, 1. Publ.; Dover Publications: Mineola, NY, USA, 2007; ISBN 978-0-486-45894-6. [Google Scholar]
- Hurley, P.J. A Concise Introduction to Logic, 12th ed.; Cengage Learning: Melbourne, Australia, 2015; ISBN 978-1-285-19654-1. [Google Scholar]
- Halford, G.S.; Wilson, W.H.; Phillips, S. Relational Knowledge: The Foundation of Higher Cognition. Trends Cogn. Sci. 2010, 14, 497–505. [Google Scholar] [CrossRef]
- Peirce, J.; Gray, J.R.; Simpson, S.; MacAskill, M.; Höchenberger, R.; Sogo, H.; Kastman, E.; Lindeløv, J.K. PsychoPy2: Experiments in Behavior Made Easy. Behav. Res. Methods 2019, 51, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Iaria, G.; Petrides, M. Occipital Sulci of the Human Brain: Variability and Probability Maps. J. Comp. Neurol. 2007, 501, 243–259. [Google Scholar] [CrossRef]
- Mayka, M.A.; Corcos, D.M.; Leurgans, S.E.; Vaillancourt, D.E. Three-Dimensional Locations and Boundaries of Motor and Premotor Cortices as Defined by Functional Brain Imaging: A Meta-Analysis. NeuroImage 2006, 31, 1453–1474. [Google Scholar] [CrossRef] [Green Version]
- Niskanen, E.; Julkunen, P.; Säisänen, L.; Vanninen, R.; Karjalainen, P.; Könönen, M. Group-Level Variations in Motor Representation Areas of Thenar and Anterior Tibial Muscles: Navigated Transcranial Magnetic Stimulation Study. Hum. Brain Mapp. 2010, 31, 1272–1280. [Google Scholar] [CrossRef]
- Sarfeld, A.S.; Diekhoff, S.; Wang, L.E.; Liuzzi, G.; Uludaǧ, K.; Eickhoff, S.B.; Fink, G.R.; Grefkes, C. Convergence of Human Brain Mapping Tools: Neuronavigated TMS Parameters and FMRI Activity in the Hand Motor Area. Hum. Brain Mapp. 2012, 33, 1107–1123. [Google Scholar] [CrossRef]
- Smith, S.M.; Jenkinson, M.; Woolrich, M.W.; Beckmann, C.F.; Behrens, T.E.J.; Johansen-Berg, H.; Bannister, P.R.; De Luca, M.; Drobnjak, I.; Flitney, D.E.; et al. Advances in Functional and Structural MR Image Analysis and Implementation as FSL. NeuroImage 2004, 23 (Suppl. S1), S208–S219. [Google Scholar] [CrossRef] [Green Version]
- Oberman, L.; Edwards, D.; Eldaief, M.; Pascual-Leone, A. Safety of Theta Burst Transcranial Magnetic Stimulation: A Systematic Review of the Literature. J. Clin. Neurophysiol. 2011, 28, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Christov-Moore, L.; Iacoboni, M. Self-Other Resonance, Its Control and Prosocial Inclinations: Brain-Behavior Relationships. Hum. Brain Mapp. 2016, 37, 1544–1558. [Google Scholar] [CrossRef] [Green Version]
- Klucharev, V.; Munneke, M.A.M.; Smidts, A.; Fernandez, G. Downregulation of the Posterior Medial Frontal Cortex Prevents Social Conformity. J. Neurosci. 2011, 31, 11934–11940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christov-Moore, L.; Sugiyama, T.; Grigaityte, K.; Iacoboni, M. Increasing Generosity by Disrupting Prefrontal Cortex. Soc. Neurosci. 2017, 12, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Deblieck, C.; Thompson, B.; Iacoboni, M.; Wu, A.D. Correlation between Motor and Phosphene Thresholds: A Transcranial Magnetic Stimulation Study. Hum. Brain Mapp. 2008, 29, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, P.B.; Fountain, S.; Daskalakis, Z.J. A Comprehensive Review of the Effects of RTMS on Motor Cortical Excitability and Inhibition. Clin. Neurophysiol. 2006, 117, 2584–2596. [Google Scholar] [CrossRef]
- Stokes, M.G.; Chambers, C.D.; Gould, I.C.; Henderson, T.R.; Janko, N.E.; Allen, N.B.; Mattingley, J.B. Simple Metric for Scaling Motor Threshold Based on Scalp-Cortex Distance: Application to Studies Using Transcranial Magnetic Stimulation. J. Neurophysiol. 2005, 94, 4520–4527. [Google Scholar] [CrossRef] [Green Version]
- Fox, N.C.; Schott, J.M. Imaging Cerebral Atrophy: Normal Ageing to Alzheimer’s Disease. Lancet 2004, 363, 392–394. [Google Scholar] [CrossRef] [PubMed]
- Lashley, K.S. The Problem of Serial Order in Behavior. In Cerebral Mechanisms in Behavior: The Hixon Symposium; Wiley: Hoboken, NJ, USA, 1951. [Google Scholar]
- Chomsky, N. Syntactic Structures; Mouton: The Hague, The Netherlands, 1957. [Google Scholar]
- Chomsky, N. Aspects of the Theory of Syntax; MIT Press: Cambridge, MA, USA, 1965. [Google Scholar]
- Ernest, P. A Model of The Cognitive Meaning of Mathematical Expressions. Br. J. Educ. Psychol. 1987, 57, 343–370. [Google Scholar] [CrossRef]
- Monti, M.M.; Parsons, L.M.; Osherson, D.N. Thought Beyond Language Neural Dissociation of Algebra and Natural Language. Psychol. Sci. 2012, 23, 914–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amalric, M.; Dehaene, S. Origins of the Brain Networks for Advanced Mathematics in Expert Mathematicians. Proc. Natl. Acad. Sci. USA 2016, 113, 4909–4917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koelsch, S.; Rohrmeier, M.; Torrecuso, R.; Jentschke, S. Processing of Hierarchical Syntactic Structure in Music. Proc. Natl. Acad. Sci. USA 2013, 110, 15443–15448. [Google Scholar] [CrossRef] [Green Version]
- Lerdahl, F.; Jackendoff, R. A Generative Theory of Tonal Music; MIT Press: Cambridge, MA, USA, 2010; ISBN 978-0-262-62107-6. [Google Scholar]
- Patel, A.D. Language, Music, Syntax and the Brain. Nat. Neurosci. 2003, 6, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Botvinick, M.M. Hierarchical Models of Behavior and Prefrontal Function. Trends Cogn. Sci. 2008, 12, 201–208. [Google Scholar] [CrossRef]
- Reverberi, C.; Shallice, T.; D’Agostini, S.; Skrap, M.; Bonatti, L.L. Cortical Bases of Elementary Deductive Reasoning: Inference, Memory, and Metadeduction. Neuropsychologia 2009, 47, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, C.; Bonatti, L.L.; Frackowiak, R.S.J.; Paulesu, E.; Cherubini, P.; Macaluso, E. Large Scale Brain Activations Predict Reasoning Profiles. NeuroImage 2012, 59, 1752–1764. [Google Scholar] [CrossRef] [PubMed]
- Moayedi, M.; Salomons, T.V.; Dunlop, K.A.M.; Downar, J.; Davis, K.D. Connectivity-Based Parcellation of the Human Frontal Polar Cortex. Brain Struct. Funct. 2015, 220, 2603–2616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prado, J.; Mutreja, R.; Booth, J.R. Fractionating the Neural Substrates of Transitive Reasoning: Task-Dependent Contributions of Spatial and Verbal Representations. Cereb. Cortex 2013, 23, 499–507. [Google Scholar] [CrossRef] [Green Version]
- Knauff, M.; Fangmeier, T.; Ruff, C.C.; Johnson-Laird, P.N. Reasoning, Models, and Images: Behavioral Measures and Cortical Activity. J. Cogn. Neurosci. 2003, 15, 559–573. [Google Scholar] [CrossRef] [Green Version]
- Prado, J.; Spotorno, N.; Koun, E.; Hewitt, E.; Van der Henst, J.-B.; Sperber, D.; Noveck, I.A. Neural Interaction between Logical Reasoning and Pragmatic Processing in Narrative Discourse. J. Cogn. Neurosci. 2015, 27, 692–704. [Google Scholar] [CrossRef]
- Schwartz, F.; Epinat-Duclos, J.; Noveck, I.; Prado, J. The Neural Development of Pragmatic Inference-Making in Natural Discourse. Dev. Sci. 2018, 21, e12678. [Google Scholar] [CrossRef] [Green Version]
- Cesana-Arlotti, A.N.; Martín, A.; Téglás, E. Precursors of Logical Reasoning in Preverbal Infants. Science 2018, 1266, 25–27. [Google Scholar] [CrossRef] [Green Version]
- Kroger, J.K.; Nystrom, L.E.; Cohen, J.D.; Johnson-Laird, P.N. Distinct Neural Substrates for Deductive and Mathematical Processing. Brain Res. 2008, 1243, 86–103. [Google Scholar] [CrossRef]
- Rogalsky, C.; Hickok, G. The Role of Broca’s Area in Sentence Comprehension. J. Cogn. Neurosci. 2010, 23, 1664–1680. [Google Scholar] [CrossRef] [PubMed]
- Boeckx, C. Language in Cognition: Uncovering Mental Structures and the Rules behind Them; Wiley-Blackwell: Malden, MA, USA, 2010. [Google Scholar]
- Spelke, E. What Makes Us Smart? Core Knowledge and Natural Language. In Language in Mind: Advances in the Study of Language and Thought; Gentner, D., Goldin-Meadow, S., Eds.; MIT Press: Cambridge, MA, USA, 2003. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reasoning Task | |||
|---|---|---|---|
| Type | Terms | Matching | Non-Matching |
| Log | 3 | If both X and Z then not Y. | If either Y or Z then not X. |
| If Y then either not X or not Z. | If X then both Y and Z. | ||
| Log | 4 | If both X and not Z then either Y or not W. | If both not Y and not Q then both Z and X. |
| If both W and not Y then either Z or not X. | If both Z and X then both not Y and not W. | ||
| Ling | 3 | It was X that Y saw Z take. | It was Y that Z thought X said. |
| Z was seen by Y taking X. | Z was thought by Y to have said X. | ||
| Ling | 4 | It was X that W heard Y saw Z take. | What W knew that Y gave Z was X. |
| W heard that Z was seen by Y taking X. | It was X that W knew was given to Y by Z. | ||
| Grammaticality Judgment Task | |||
| Type | Terms | Grammatical | Non-Grammatical |
| Log | 3 | If either Y or X then not Z. | If not Y then Z both and X. |
| If Y then either X or Z. | If either not Z or not X then not Y. | ||
| Log | 4 | If either X or W then both Y and Z. | If both Z and not Y then either X or not W. |
| If both not Y and not W then both Z and X. | If both W and Y then either not X not or Z. | ||
| Ling | 3 | Z was thought by Y to have said X. | It was to Y that from Z told X. |
| It was Y that X thought Z said. | What Z told Y was X. | ||
| Ling | 4 | Z knows that X is given by Y to W. | Z will be seen by Y taking X is what W will hear. |
| If either W or X then both Y and not Z. | It was X that W heard Y take Z saw. | ||
| Stimulation Site | ||||||
|---|---|---|---|---|---|---|
| LBA44 | MBA8 | LTOS | ||||
| Pre-cTBS | Post-cTBS | Pre-cTBS | Post-cTBS | Pre-cTBS | Post-cTBS | |
| Linguistic Reasoning | 94% | 87% | 82% | 85% | 85% | 87% |
| Logic Reasoning | 72% | 76% | 78% | 75% | 69% | 78% |
| Grammaticality Judgment | 90% | 83% | 83% | 85% | 80% | 85% |
| Contrast | Task | Stim. Site & Contrast Weights | F | Signature |
|---|---|---|---|---|
| 1 | Linguistic | LBA44(−1) vs. LTOS(+0.5) & MBA8(+0.5) | 8.207 | 0.012 * |
| 2 | LBA44(+0.5) & LTOS(+0.5) vs. MBA8(−1) | 1.456 | 0.248 | |
| 3 | Logic | LBA44(−1) vs. LTOS(+0.5) & MBA8(+0.5) | 0.176 | 0.681 |
| 4 | LBA44(+0.5) & LTOS(+0.5) vs. MBA8(−1) | 10.652 | 0.006 * | |
| 5 | Grammar | LBA44(−1) vs. LTOS(+0.5) & MBA8(+0.5) | 8.708 | 0.011 * |
| 6 | LBA44(+0.5) & LTOS(+0.5) vs. MBA8(−1) | 0.994 | 0.336 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coetzee, J.P.; Johnson, M.A.; Lee, Y.; Wu, A.D.; Iacoboni, M.; Monti, M.M. Dissociating Language and Thought in Human Reasoning. Brain Sci. 2023, 13, 67. https://doi.org/10.3390/brainsci13010067
Coetzee JP, Johnson MA, Lee Y, Wu AD, Iacoboni M, Monti MM. Dissociating Language and Thought in Human Reasoning. Brain Sciences. 2023; 13(1):67. https://doi.org/10.3390/brainsci13010067
Chicago/Turabian StyleCoetzee, John P., Micah A. Johnson, Youngzie Lee, Allan D. Wu, Marco Iacoboni, and Martin M. Monti. 2023. "Dissociating Language and Thought in Human Reasoning" Brain Sciences 13, no. 1: 67. https://doi.org/10.3390/brainsci13010067