
Arousal Regulation by the External Globus Pallidus: A New Node for the Mesocircuit Hypothesis
{kind=link}
Abstract
:1. Introduction
2. GPe in Arousal/Sleep-Wake Regulation
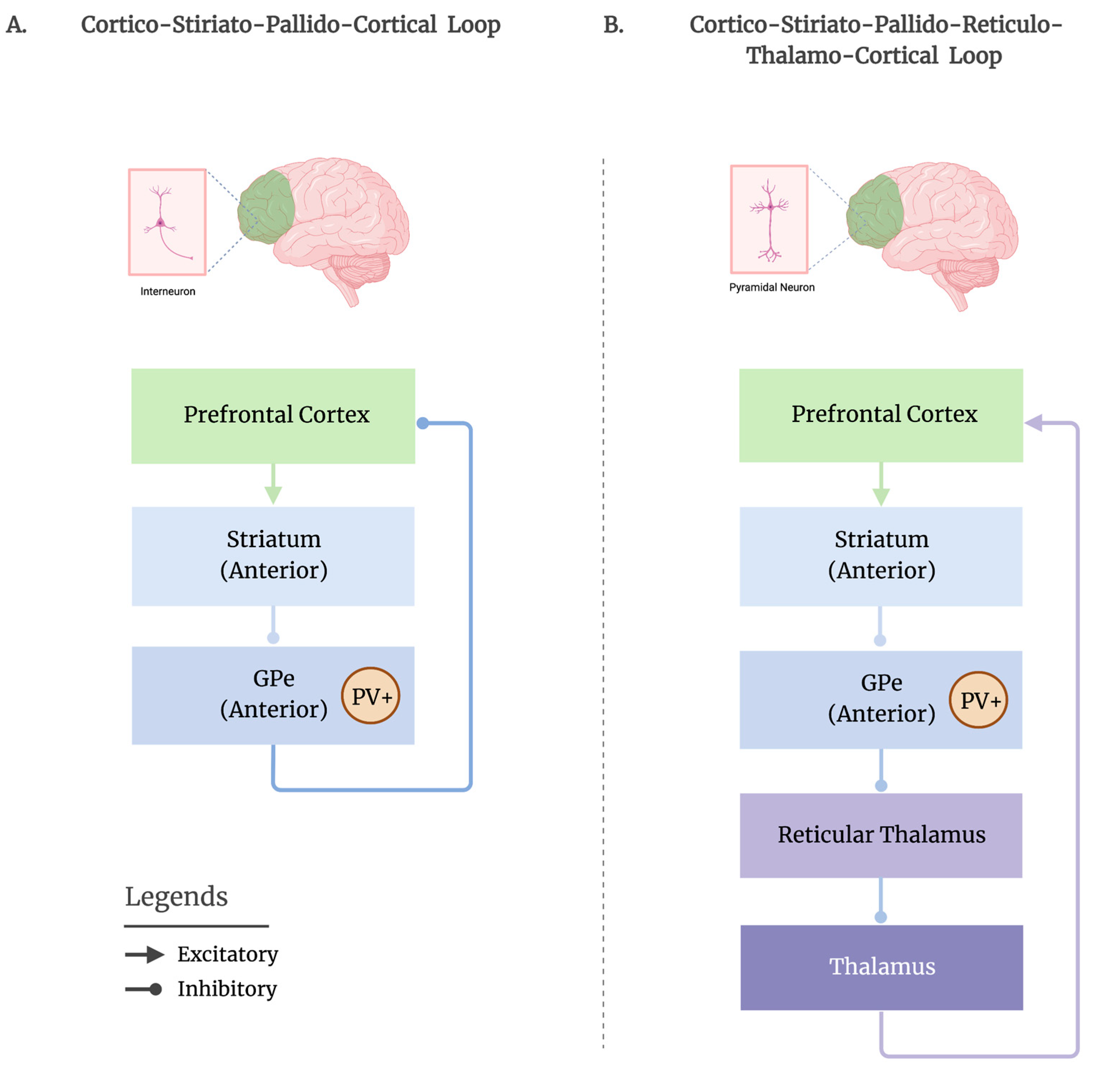
3. Proposed Striato-Pallidal Circuitries for Promoting Sleep
3.1. Cortico-Striato-Pallido-Cortical Route
3.2. Cortico-Striato-Pallido-Reticulo-Thalamo-Cortical Route
4. Globus Pallidus in Consciousness Disruption
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Llinás, R.; Ribary, U.; Contreras, D.; Pedroarena, C. The Neuronal Basis for Consciousness. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1998, 353, 1841–1849. [Google Scholar] [PubMed] [Green Version]
- Kostopoulos, G.K. Involvement of the Thalamocortical System in Epileptic Loss of Consciousness. Epilepsia 2001, 42 (Suppl. 3), 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumenfeld, H. Impaired Consciousness in Epilepsy. Lancet Neurol. 2012, 11, 814–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCafferty, C.; David, F.; Venzi, M.; Lőrincz, M.L.; Delicata, F.; Atherton, Z.; Recchia, G.; Orban, G.; Lambert, R.C.; Di Giovanni, G.; et al. Cortical Drive and Thalamic Feed-Forward Inhibition Control Thalamic Output Synchrony during Absence Seizures. Nat. Neurosci. 2018, 21, 744–756. [Google Scholar] [CrossRef] [PubMed]
- Schiff, N.D. Recovery of Consciousness after Brain Injury: A Mesocircuit Hypothesis. Trends Neurosci. 2010, 33, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Scannell, J.W.; Burns, G.A.P.C.; Hilgetag, C.C.; O’Neil, M.A.; Young, M.P. The Connectional Organization of the Cortico-Thalamic System of the Cat. Cereb. Cortex 1999, 9, 277–299. [Google Scholar] [CrossRef] [Green Version]
- Schiff, N.D. Mesocircuit Mechanisms Underlying Recovery of Consciousness Following Severe Brain Injuries: Model and Predictions. In Brain Function and Responsiveness in Disorders of Consciousness; Monti, M.M., Sannita, W.G., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 195–204. ISBN 978-3-319-21425-2. [Google Scholar]
- Qiu, M.-H.; Vetrivelan, R.; Fuller, P.M.; Lu, J. Basal Ganglia Control of Sleep-Wake Behavior and Cortical Activation. Eur. J. Neurosci. 2010, 31, 499–507. [Google Scholar] [CrossRef]
- Qiu, M.H.; Chen, M.C.; Wu, J.; Nelson, D.; Lu, J. Deep Brain Stimulation in the Globus Pallidus Externa Promotes Sleep. Neuroscience 2016, 322, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Qiu, M.-H.; Yao, Q.-L.; Vetrivelan, R.; Chen, M.C.; Lu, J. Nigrostriatal Dopamine Acting on Globus Pallidus Regulates Sleep. Cereb. Cortex 2016, 26, 1430–1439. [Google Scholar] [CrossRef] [Green Version]
- Vetrivelan, R.; Qiu, M.-H.; Chang, C.; Lu, J. Role of Basal Ganglia in Sleep-Wake Regulation: Neural Circuitry and Clinical Significance. Front. Neuroanat. 2010, 4, 145. [Google Scholar] [CrossRef]
- Yuan, X.-S.; Wang, L.; Dong, H.; Qu, W.-M.; Yang, S.-R.; Cherasse, Y.; Lazarus, M.; Schiffmann, S.N.; de d’Exaerde, A.K.; Li, R.-X.; et al. Striatal Adenosine A2A Receptor Neurons Control Active-Period Sleep via Parvalbumin Neurons in External Globus Pallidus. eLife 2017, 6, e29055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.C.; Ferrari, L.; Sacchet, M.D.; Foland-Ross, L.C.; Qiu, M.-H.; Gotlib, I.H.; Fuller, P.M.; Arrigoni, E.; Lu, J. Identification of a Direct GABAergic Pallidocortical Pathway in Rodents. Eur. J. Neurosci. 2015, 41, 748–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, A.; Oldenburg, I.A.; Berezovskii, V.K.; Johnson, C.A.; Kingery, N.D.; Elliott, H.L.; Xie, T.; Gerfen, C.R.; Sabatini, B.L. A Direct GABAergic Output from the Basal Ganglia to Frontal Cortex. Nature 2015, 521, 85–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.S.; Monti, M.M. Cortical and Thalamic Connections of the Human Globus Pallidus: Implications for Disorders of Consciousness. Front. Neuroanat. 2022, 16, 960439. [Google Scholar] [CrossRef] [PubMed]
- Cornwall, J.; Cooper, J.D.; Phillipson, O.T. Projections to the Rostral Reticular Thalamic Nucleus in the Rat. Exp. Brain Res. 1990, 80, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Gandia, J.; De Las Heras, S.; García, M.; Giménez-Amaya, J.M. Afferent Projections to the Reticular Thalamic Nucleus from the Globus Pallidus and the Substantia Nigra in the Rat. Brain Res. Bull. 1993, 32, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Hazrati, L.N.; Parent, A. Projection from the External Pallidum to the Reticular Thalamic Nucleus in the Squirrel Monkey. Brain Res. 1991, 550, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.H. The Basal Ganglia in Action. Neuroscientist 2017, 23, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Saga, Y.; Hoshi, E.; Tremblay, L. Roles of Multiple Globus Pallidus Territories of Monkeys and Humans in Motivation, Cognition and Action: An Anatomical, Physiological and Pathophysiological Review. Front. Neuroanat. 2017, 11, 30. [Google Scholar] [CrossRef] [Green Version]
- DeLong, M.R. Primate Models of Movement Disorders of Basal Ganglia Origin. Trends Neurosci. 1990, 13, 281–285. [Google Scholar] [CrossRef]
- Castillo, P.R.; Middlebrooks, E.H.; Grewal, S.S.; Okromelidze, L.; Meschia, J.F.; Quinones-Hinojosa, A.; Uitti, R.J.; Wharen, R.E. Globus Pallidus Externus Deep Brain Stimulation Treats Insomnia in a Patient With Parkinson Disease. Mayo Clin. Proc. 2020, 95, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Bjorness, T.E.; Greene, R.W. Adenosine and Sleep. Curr. Neuropharmacol. 2009, 7, 238–245. [Google Scholar] [CrossRef] [PubMed]
- de Lera Ruiz, M.; Lim, Y.-H.; Zheng, J. Adenosine A2A Receptor as a Drug Discovery Target. J. Med. Chem. 2014, 57, 3623–3650. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros-Yáñez, I.; Castillo, C.A.; Merighi, S.; Gessi, S. The Role of Adenosine Receptors in Psychostimulant Addiction. Front. Pharmacol. 2018, 8, 985. [Google Scholar] [CrossRef] [Green Version]
- Monti, J.M.; Monti, D. The Involvement of Dopamine in the Modulation of Sleep and Waking. Sleep Med. Rev. 2007, 11, 113–133. [Google Scholar] [CrossRef]
- Arnulf, I.; Nielsen, J.; Lohmann, E.; Schiefer, J.; Schieffer, J.; Wild, E.; Jennum, P.; Konofal, E.; Walker, M.; Oudiette, D.; et al. Rapid Eye Movement Sleep Disturbances in Huntington Disease. Arch. Neurol. 2008, 65, 482–488. [Google Scholar] [CrossRef] [Green Version]
- Cooper, A.J.; Stanford, I.M. Dopamine D2 Receptor Mediated Presynaptic Inhibition of Striatopallidal GABA(A) IPSCs in Vitro. Neuropharmacology 2001, 41, 62–71. [Google Scholar] [CrossRef]
- Querejeta, E.; Delgado, A.; Valdiosera, R.; Erlij, D.; Aceves, J. Intrapallidal D2 Dopamine Receptors Control Globus Pallidus Neuron Activity in the Rat. Neurosci. Lett. 2001, 300, 79–82. [Google Scholar] [CrossRef]
- Beggiato, S.; Antonelli, T.; Tomasini, M.C.; Borelli, A.C.; Agnati, L.F.; Tanganelli, S.; Fuxe, K.; Ferraro, L. Adenosine A2A-D2 Receptor-Receptor Interactions in Putative Heteromers in the Regulation of the Striato-Pallidal Gaba Pathway: Possible Relevance for Parkinson’s Disease and Its Treatment. Curr. Protein Pept. Sci. 2014, 15, 673–680. [Google Scholar] [CrossRef]
- Ferré, S.; Diamond, I.; Goldberg, S.R.; Yao, L.; Hourani, S.M.O.; Huang, Z.L.; Urade, Y.; Kitchen, I. Adenosine A2A Receptors in Ventral Striatum, Hypothalamus and Nociceptive Circuitry: Implications for Drug Addiction, Sleep and Pain. Prog. Neurobiol. 2007, 83, 332–347. [Google Scholar] [CrossRef]
- Fuxe, K.; Agnati, L.F.; Jacobsen, K.; Hillion, J.; Canals, M.; Torvinen, M.; Tinner-Staines, B.; Staines, W.; Rosin, D.; Terasmaa, A.; et al. Receptor Heteromerization in Adenosine A2A Receptor Signaling: Relevance for Striatal Function and Parkinson’s Disease. Neurology 2003, 61, S19–S23. [Google Scholar] [CrossRef]
- Villar-Menéndez, I.; Porta, S.; Buira, S.P.; Pereira-Veiga, T.; Díaz-Sánchez, S.; Albasanz, J.L.; Ferrer, I.; Martín, M.; Barrachina, M. Increased Striatal Adenosine A2A Receptor Levels Is an Early Event in Parkinson’s Disease-Related Pathology and It Is Potentially Regulated by MiR-34b. Neurobiol. Dis. 2014, 69, 206–214. [Google Scholar] [CrossRef]
- Mitchell, I.J.; Clarke, C.E.; Boyce, S.; Robertson, R.G.; Peggs, D.; Sambrook, M.A.; Crossman, A.R. Neural Mechanisms Underlying Parkinsonian Symptoms Based upon Regional Uptake of 2-Deoxyglucose in Monkeys Exposed to 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine. Neuroscience 1989, 32, 213–226. [Google Scholar] [CrossRef]
- Gerfen, C.R.; Engber, T.M.; Mahan, L.C.; Susel, Z.; Chase, T.N.; Monsma, F.J.; Sibley, D.R. D1 and D2 Dopamine Receptor-Regulated Gene Expression of Striatonigral and Striatopallidal Neurons. Science 1990, 250, 1429–1432. [Google Scholar] [CrossRef] [PubMed]
- Akkal, D.; Dum, R.P.; Strick, P.L. Supplementary Motor Area and Presupplementary Motor Area: Targets of Basal Ganglia and Cerebellar Output. J. Neurosci. 2007, 27, 10659–10673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abecassis, Z.A.; Berceau, B.L.; Win, P.H.; García, D.; Xenias, H.S.; Cui, Q.; Pamukcu, A.; Cherian, S.; Hernández, V.M.; Chon, U.; et al. Npas1+-Nkx2.1+ Neurons Are an Integral Part of the Cortico-Pallido-Cortical Loop. J. Neurosci. 2020, 40, 743–768. [Google Scholar] [CrossRef]
- Castillo, P.R.; Benarroch, E.E. How Could the Basal Ganglia Control Sleep? Neurology 2020, 95, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B. Organization of Cerebral Cortical Afferent Systems in the Rat. II. Magnocellular Basal Nucleus. J. Comp. Neurol. 1984, 222, 313–342. [Google Scholar] [CrossRef]
- Sanchez-Vives, M.V.; McCormick, D.A. Cellular and Network Mechanisms of Rhythmic Recurrent Activity in Neocortex. Nat. Neurosci. 2000, 3, 1027–1034. [Google Scholar] [CrossRef]
- Naka, A.; Adesnik, H. Inhibitory Circuits in Cortical Layer 5. Front. Neural Circuits 2016, 10, 35. [Google Scholar] [CrossRef]
- Borbély, A.A.; Daan, S.; Wirz-Justice, A.; Deboer, T. The Two-Process Model of Sleep Regulation: A Reappraisal. J. Sleep Res. 2016, 25, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Siclari, F.; Tononi, G. Local Aspects of Sleep and Wakefulness. Curr. Opin. Neurobiol. 2017, 44, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Pinault, D. The Thalamic Reticular Nucleus: Structure, Function and Concept. Brain Res. Brain Res. Rev. 2004, 46, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Kayahara, T.; Nakano, K. The Globus Pallidus Sends Axons to the Thalamic Reticular Nucleus Neurons Projecting to the Centromedian Nucleus of the Thalamus: A Light and Electron Microscope Study in the Cat. Brain Res. Bull. 1998, 45, 623–630. [Google Scholar] [CrossRef]
- Pazo, J.H.; Barceló, A.C.; Bellantonio, E.; Pazo, V.C.; Almarares, N. Electrophysiologic Study of Globus Pallidus Projections to the Thalamic Reticular Nucleus. Brain Res. Bull. 2013, 94, 82–89. [Google Scholar] [CrossRef]
- Villalobos, N.; Oviedo-Chávez, A.; Alatorre, A.; Ríos, A.; Barrientos, R.; Delgado, A.; Querejeta, E. Striatum and Globus Pallidus Control the Electrical Activity of Reticular Thalamic Nuclei. Brain Res. 2016, 1644, 258–266. [Google Scholar] [CrossRef]
- Steriade, M.; Llinás, R.R. The Functional States of the Thalamus and the Associated Neuronal Interplay. Physiol. Rev. 1988, 68, 649–742. [Google Scholar] [CrossRef]
- Crunelli, V.; Lőrincz, M.L.; Connelly, W.M.; David, F.; Hughes, S.W.; Lambert, R.C.; Leresche, N.; Errington, A.C. Dual Function of Thalamic Low-Vigilance State Oscillations: Rhythm-Regulation and Plasticity. Nat. Rev. Neurosci. 2018, 19, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.G.; Cadavieco, M.C.; Jego, S.; Ponomarenko, A.; Korotkova, T.; Adamantidis, A. Hypothalamic Feedforward Inhibition of Thalamocortical Network Controls Arousal and Consciousness. Nat. Neurosci. 2016, 19, 290–298. [Google Scholar] [CrossRef] [Green Version]
- Ni, K.-M.; Hou, X.-J.; Yang, C.-H.; Dong, P.; Li, Y.; Zhang, Y.; Jiang, P.; Berg, D.K.; Duan, S.; Li, X.-M. Selectively Driving Cholinergic Fibers Optically in the Thalamic Reticular Nucleus Promotes Sleep. eLife 2016, 5, e10382. [Google Scholar] [CrossRef]
- Vantomme, G.; Osorio-Forero, A.; Lüthi, A.; Fernandez, L.M.J. Regulation of Local Sleep by the Thalamic Reticular Nucleus. Front. Neurosci. 2019, 13, 576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, L.M.; Vantomme, G.; Osorio-Forero, A.; Cardis, R.; Béard, E.; Lüthi, A. Thalamic Reticular Control of Local Sleep in Mouse Sensory Cortex. eLife 2018, 7, e39111. [Google Scholar] [CrossRef] [PubMed]
- McCormick, D.A.; Bal, T. Sensory Gating Mechanisms of the Thalamus. Curr. Opin. Neurobiol. 1994, 4, 550–556. [Google Scholar] [CrossRef]
- Nakajima, M.; Schmitt, L.I.; Halassa, M.M. Prefrontal Cortex Regulates Sensory Filtering through a Basal Ganglia-to-Thalamus Pathway. Neuron 2019, 103, 445–458.e10. [Google Scholar] [CrossRef] [PubMed]
- Crone, J.S.; Lutkenhoff, E.S.; Bio, B.J.; Laureys, S.; Monti, M.M. Testing Proposed Neuronal Models of Effective Connectivity Within the Cortico-Basal Ganglia-Thalamo-Cortical Loop During Loss of Consciousness. Cereb. Cortex 2017, 27, 2727–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutkenhoff, E.S.; Chiang, J.; Tshibanda, L.; Kamau, E.; Kirsch, M.; Pickard, J.D.; Laureys, S.; Owen, A.M.; Monti, M.M. Thalamic and Extrathalamic Mechanisms of Consciousness after Severe Brain Injury. Ann. Neurol. 2015, 78, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Lutkenhoff, E.S.; Johnson, M.A.; Casarotto, S.; Massimini, M.; Monti, M.M. Subcortical Atrophy Correlates with the Perturbational Complexity Index in Patients with Disorders of Consciousness. Brain Stimul. 2020, 13, 1426–1435. [Google Scholar] [CrossRef]
- Lutkenhoff, E.S.; Nigri, A.; Sebastiano, D.R.; Sattin, D.; Visani, E.; Rosazza, C.; D’Incerti, L.; Bruzzone, M.G.; Franceschetti, S.; Leonardi, M.; et al. EEG Power Spectra and Subcortical Pathology in Chronic Disorders of Consciousness. Psychol. Med. 2022, 52, 1491–1500. [Google Scholar] [CrossRef]
- Swainston Harrison, T.; Keating, G.M. Zolpidem: A Review of Its Use in the Management of Insomnia. CNS Drugs 2005, 19, 65–89. [Google Scholar] [CrossRef]
- Tassinari, C.A.; Rubboli, G.; Gardella, E.; Cantalupo, G.; Calandra-Buonaura, G.; Vedovello, M.; Alessandria, M.; Gandini, G.; Cinotti, S.; Zamponi, N.; et al. Central Pattern Generators for a Common Semiology in Fronto-Limbic Seizures and in Parasomnias. A Neuroethologic Approach. Neurol. Sci. 2005, 26, s225–s232. [Google Scholar] [CrossRef]
- Xue, Y.; Han, X.-H.; Chen, L. Effects of Pharmacological Block of GABAA Receptors on Pallidal Neurons in Normal and Parkinsonian State. Front. Cell. Neurosci. 2010, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiff, N.D.; Posner, J.B. Another “Awakenings”. Ann. Neurol. 2007, 62, 5–7. [Google Scholar] [CrossRef]
- Laplane, D.; Levasseur, M.; Pillon, B.; Dubois, B.; Baulac, M.; Mazoyer, B.; Tran Dinh, S.; Sette, G.; Danze, F.; Baron, J.C. Obsessive-Compulsive and Other Behavioural Changes with Bilateral Basal Ganglia Lesions. A Neuropsychological, Magnetic Resonance Imaging and Positron Tomography Study. Brain 1989, 112 Pt 3, 699–725. [Google Scholar] [CrossRef]
- Brefel-Courbon, C.; Payoux, P.; Ory, F.; Sommet, A.; Slaoui, T.; Raboyeau, G.; Lemesle, B.; Puel, M.; Montastruc, J.-L.; Demonet, J.-F.; et al. Clinical and Imaging Evidence of Zolpidem Effect in Hypoxic Encephalopathy. Ann. Neurol. 2007, 62, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Clauss, R.P.; Güldenpfennig, W.M.; Nel, H.W.; Sathekge, M.M.; Venkannagari, R.R. Extraordinary Arousal from Semi-Comatose State on Zolpidem. A Case Report. S. Afr. Med. J. 2000, 90, 68–72. [Google Scholar] [PubMed]
- Williams, S.T.; Conte, M.M.; Goldfine, A.M.; Noirhomme, Q.; Gosseries, O.; Thonnard, M.; Beattie, B.; Hersh, J.; Katz, D.I.; Victor, J.D.; et al. Common Resting Brain Dynamics Indicate a Possible Mechanism Underlying Zolpidem Response in Severe Brain Injury. Elife 2013, 2, e01157. [Google Scholar] [CrossRef]
- Chatelle, C.; Thibaut, A.; Gosseries, O.; Bruno, M.-A.; Demertzi, A.; Bernard, C.; Hustinx, R.; Tshibanda, L.; Bahri, M.A.; Laureys, S. Changes in Cerebral Metabolism in Patients with a Minimally Conscious State Responding to Zolpidem. Front. Hum. Neurosci. 2014, 8, 917. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Z.S.; Reggente, N.; Monti, M.M. Arousal Regulation by the External Globus Pallidus: A New Node for the Mesocircuit Hypothesis. Brain Sci. 2023, 13, 146. https://doi.org/10.3390/brainsci13010146
Zheng ZS, Reggente N, Monti MM. Arousal Regulation by the External Globus Pallidus: A New Node for the Mesocircuit Hypothesis. Brain Sciences. 2023; 13(1):146. https://doi.org/10.3390/brainsci13010146
Chicago/Turabian StyleZheng, Zhong Sheng, Nicco Reggente, and Martin M. Monti. 2023. "Arousal Regulation by the External Globus Pallidus: A New Node for the Mesocircuit Hypothesis" Brain Sciences 13, no. 1: 146. https://doi.org/10.3390/brainsci13010146