Consecutive Injection of High-Dose Lipopolysaccharide Modulates Microglia Polarization via TREM2 to Alter Status of Septic Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Cell Line
2.2. Murine Sepsis Score Test
2.3. Cell Culture and Transfection
2.4. Lentiviral Particles Preparation
2.5. Treatments of Mice Models
2.6. Novel Object Recognition (NOR)
2.7. Y-Maze
2.8. Hematoxylin and Eosin (H&E) Staining
2.9. Immunofluorescence
2.10. Western Blot
2.11. ELISA Assay
2.12. Quantitative RT-PCR
2.13. Statistical Analysis
3. Results
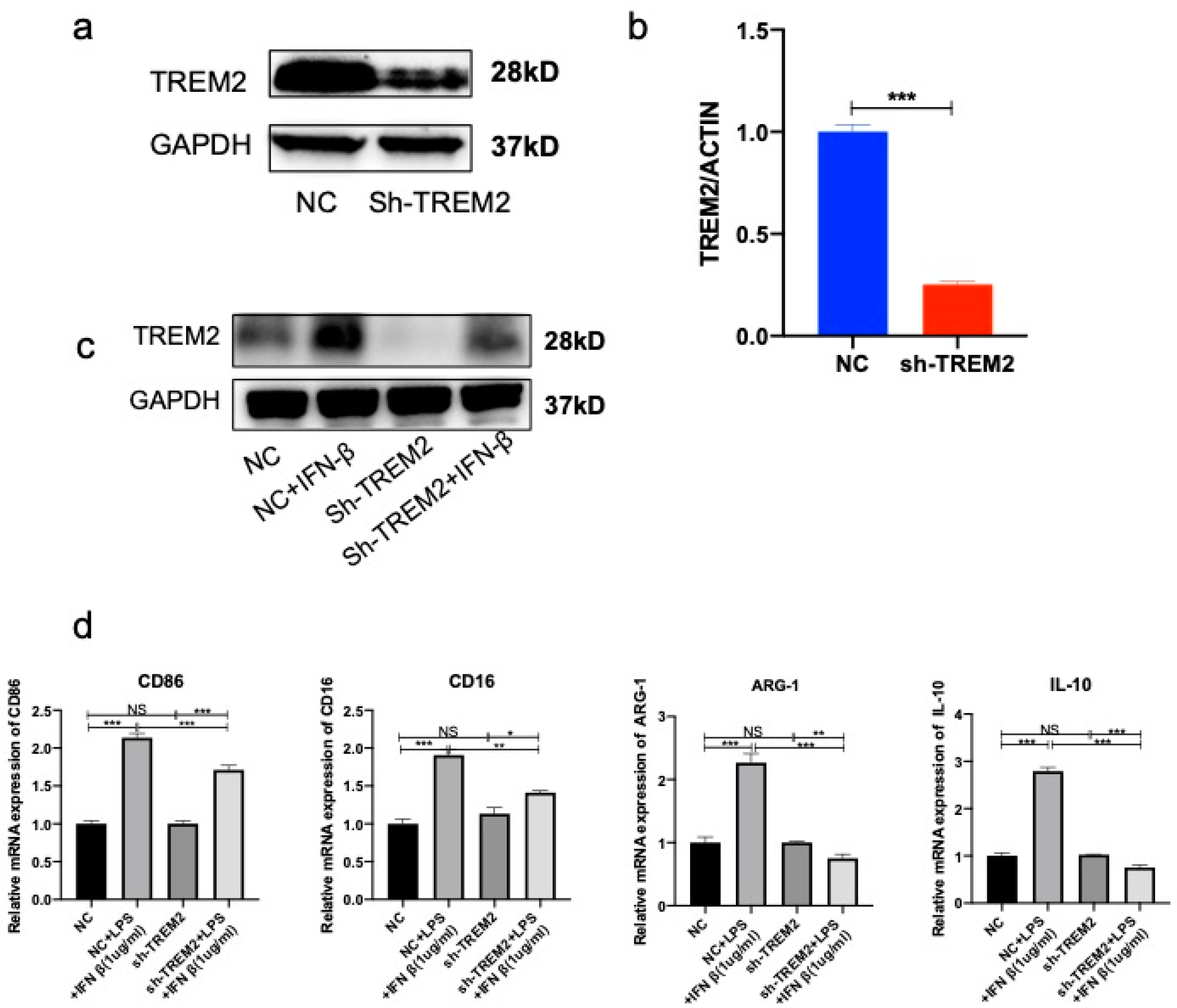
3.1. Reduction of TREM2 in Septic Model In Vivo and In Vitro
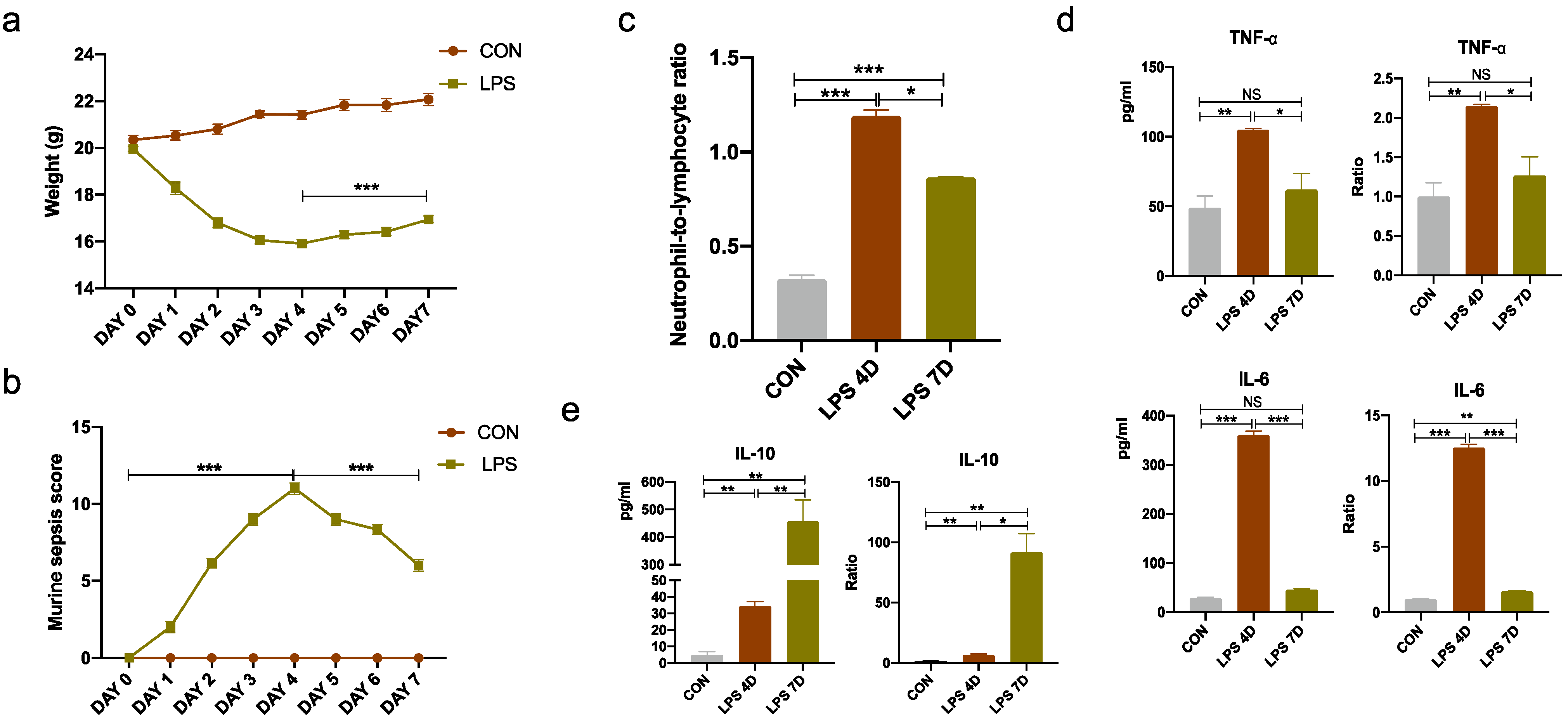
3.2. Evaluation of Inflammation in Septic Mouse Model
3.3. IFN-β Enhanced TREM2 Expression Triggering Conversion of Microglia from M1 to M2
3.4. Knockdown of TREM2 in Microglia Inhibited Conversion of Microglia from M1 Phenotype to M2 Phenotype
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A. The blood-brain barrier in neuroimmunology: Tales of separation and assimilation. Brain Behav. Immun. 2015, 44, 1312–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelon, C.; Michels, M.; Abatti, M.; Vieira, A.; Borges, H.; Dominguini, D.; Barichello, T.; Dal-Pizzol, F. The Role of Secretase Pathway in Long-term Brain Inflammation and Cognitive Impairment in an Animal Model of Severe Sepsis. Mol. Neurobiol. 2020, 57, 1159–1169. [Google Scholar] [CrossRef]
- Michels, M.; Vieira, A.S.; Vuolo, F.; Zapelini, H.G.; Mendonça, B.; Mina, F.; Dominguini, D.; Steckert, A.; Schuck, P.F.; Quevedo, J.; et al. The role of microglia activation in the development of sepsis-induced long-term cognitive impairment. Brain Behav. Immun. 2015, 43, 54–59. [Google Scholar] [CrossRef]
- Subhramanyam, C.S.; Wang, C.; Hu, Q.; Dheen, S.T. Microglia-mediated neuroinflammation in neurodegenerative diseases. Semin. Cell Dev. Biol. 2019, 94, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Bermel, R.A.; Rudick, R.A. Interferon-β treatment for multiple sclerosis. Neurotherapeutics 2007, 4, 633–646. [Google Scholar] [CrossRef]
- Teige, I.; Treschow, A.; Teige, A.; Mattsson, R.; Navikas, V.; Leanderson, T.; Holmdahl, R.; Issazadeh-Navikas, S. IFN-β gene deletion leads to augmented and chronic demyelinating experimental autoimmune encephalomyelitis. J. Immunol. 2003, 170, 4776–4784. [Google Scholar] [CrossRef] [Green Version]
- Touil, T.; Fitzgerald, D.; Zhang, G.X.; Rostami, A.; Gran, B. Cutting Edge: TLR3 stimulation suppresses experimental autoimmune encephalomyelitis by inducing endogenous IFN-β. J. Immunol. 2006, 177, 7505–7509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuohy, V.K.; Yu, M.; Yin, L.; Mathisen, P.M.; Johnson, J.M.; Kawczak, J.A. Modulation of the IL-10/IL-12 cytokine circuit by interferon-β inhibits the development of epitope spreading and disease progression in murine autoimmune encephalomyelitis. J. Neuroimmunol. 2000, 111, 55–63. [Google Scholar] [CrossRef]
- Floris, S.; Ruuls, S.R.; Wierinckx, A.; van der Pol, S.M.; Döpp, E.; van der Meide, P.H.; Dijkstra, C.D.; De Vries, H.E. Interferon-β directly influences monocyte infiltration into the central nervous system. J. Neuroimmunol. 2002, 127, 69–79. [Google Scholar] [CrossRef]
- Mimouna, S.; Rollins, D.A.; Shibu, G.; Tharmalingam, B.; Deochand, D.K.; Chen, X.; Oliver, D.; Chinenov, Y.; Rogatsky, I. Transcription cofactor GRIP1 differentially affects myeloid cell–driven neuroinflammation and response to IFN-β therapy. J. Exp. Med. 2021, 218, e20192386. [Google Scholar] [CrossRef]
- Mathur, V.; Burai, R.; Vest, R.T.; Bonanno, L.N.; Lehallier, B.; Zardeneta, M.E.; Mistry, K.N.; Do, D.; Marsh, S.E.; Abud, E.M.; et al. Activation of the STING-Dependent Type I Interferon Response Reduces Microglial Reactivity and Neuroinflammation. Neuron 2017, 96, 1290–1302. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Taso, O.; Wang, R.; Bayram, S.; Graham, A.C.; Garcia-Reitboeck, P.; Mallach, A.; Andrews, W.D.; Piers, T.M.; Botia, J.A.; et al. Trem2 promotes anti-inflammatory responses in microglia and is suppressed under pro-inflammatory conditions. Hum. Mol. Genet. 2020, 29, 3224–3248. [Google Scholar] [CrossRef]
- Ulland, T.K.; Colonna, M. TREM2—a key player in microglial biology and Alzheimer disease. Nat. Rev. Neurol. 2018, 14, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zheng, Y.; Luo, Y.; Du, Y.; Zhang, X.; Fu, J. Curcumin inhibits LPS-induced neuroinflammation by promoting microglial M2 polarization via TREM2/ TLR4/ NF-κB pathways in BV2 cells. Mol. Immunol. 2019, 116, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Gawish, R.; Martins, R.; Böhm, B.; Wimberger, T.; Sharif, O.; Lakovits, K.; Schmidt, M.; Knapp, S. Triggering receptor expressed on myeloid cells-2 fine-tunes inflammatory responses in murine Gram-negative sepsis. FASEB J. 2015, 29, 1247–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, L.-S.; Xu, Y.; Wang, Q.-S. YY1 Promotes Microglia M2 Polarization through The MIR-130A-3P/TREM-2 AXIS to Alleviate Sepsis-Associated Encephalopathy. Shock 2022, 58, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Zhai, Q.; Fang, P.; Yang, S.; Sun, Y.; Wu, S.; Huang, R.; Chen, Q.; Fang, X. Triggering receptor expressed on myeloid Cells-2 (TREM2) inhibits steroidogenesis in adrenocortical cell by macrophage-derived exosomes in lipopolysaccharide-induced septic shock. Mol. Cell. Endocrinol. 2021, 525, 111178. [Google Scholar] [CrossRef]
- Wang, Y.; Cella, M.; Mallinson, K.; Ulrich, J.D.; Young, K.L.; Robinette, M.L.; Gilfillan, S.; Krishnan, G.M.; Sudhakar, S.; Zinselmeyer, B.H.; et al. TREM2 Lipid Sensing Sustains the Microglial Response in an Alzheimer’s Disease Model. Cell 2015, 160, 1061–1071. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Peng, J.; Sherchan, P.; Ma, Y.; Xiang, S.; Yan, F.; Zhao, H.; Jiang, Y.; Wang, N.; Zhang, J.H.; et al. TREM2 activation attenuates neuroinflammation and neuronal apoptosis via PI3K/Akt pathway after intracerebral hemorrhage in mice. J. Neuroinflammation 2020, 17, 168. [Google Scholar] [CrossRef]
- Raha, A.A.; Henderson, J.W.; Stott, S.R.; Vuono, R.; Foscarin, S.; Friedland, R.P.; Zaman, S.H.; Raha-Chowdhury, R. Neuroprotective Effect of TREM-2 in Aging and Alzheimer’s Disease Model. J. Alzheimer’s Dis. 2017, 55, 199–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Prinz, M.; Stagi, M.; Chechneva, O.; Neumann, H. TREM2-Transduced Myeloid Precursors Mediate Nervous Tissue Debris Clearance and Facilitate Recovery in an Animal Model of Multiple Sclerosis. PLoS Med. 2007, 4, e124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakao, T.; Ono, Y.; Dai, H.; Nakano, R.; Perez-Gutierrez, A.; Camirand, G.; Huang, H.; Geller, D.A.; Thomson, A.W. DNAX Activating Protein of 12 kDa/Triggering Receptor Expressed on Myeloid Cells 2 Expression by Mouse and Human Liver Dendritic Cells: Functional Implications and Regulation of Liver Ischemia-Reperfusion Injury. Hepatology 2019, 70, 696–710. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, K.; Jin, Y.; Zhu, T.; Cheng, B.; Shu, Q.; Fang, X. Triggering Receptor Expressed on Myeloid Cells-2 Protects against Polymicrobial Sepsis by Enhancing Bacterial Clearance. Am. J. Respir. Crit. Care Med. 2013, 188, 201–212. [Google Scholar] [CrossRef]
- Tan, P.-H.; Ji, J.; Yeh, C.-C.; Ji, R.-R. Interferons in Pain and Infections: Emerging Roles in Neuro-Immune and Neuro-Glial Interactions. Front. Immunol. 2021, 12, 783725. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Meilandt, W.J.; Xie, L.; Gandham, V.D.; Ngu, H.; Barck, K.H.; Rezzonico, M.G.; Imperio, J.; Lalehzadeh, G.; Huntley, M.A.; et al. Trem2 restrains the enhancement of tau accumulation and neurodegeneration by β-amyloid pathology. Neuron 2021, 109, 1283–1301. [Google Scholar] [CrossRef]
- Wu, M.; Liao, M.; Huang, R.; Chen, C.; Tian, T.; Wang, H.; Li, J.; Li, J.; Sun, Y.; Wu, C.; et al. Hippocampal overexpression of TREM2 ameliorates high fat diet induced cognitive impairment and modulates phenotypic polarization of the microglia. Genes Dis. 2020, 9, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Xu, W.; Cheng, H.; Yuan, H.; Tan, X. Efficacy and mechanism of cGAMP to suppress Alzheimer’s disease by elevating TREM2. Brain Behav. Immun. 2019, 81, 495–508. [Google Scholar] [CrossRef]
- Guo, Y.; Wei, X.; Yan, H.; Qin, Y.; Yan, S.; Liu, J.; Zhao, Y.; Jiang, F.; Lou, H. TREM2 deficiency aggravates α-synuclein-induced neurodegeneration and neuroinflammation in Parkinson’s disease models. FASEB J. 2019, 33, 12164–12174. [Google Scholar] [CrossRef] [Green Version]
- Filipello, F.; Morini, R.; Corradini, I.; Zerbi, V.; Canzi, A.; Michalski, B.; Erreni, M.; Markicevic, M.; Starvaggi-Cucuzza, C.; Otero, K.; et al. The Microglial Innate Immune Receptor TREM2 Is Required for Synapse Elimination and Normal Brain Connectivity. Immunity 2018, 48, 979–991. [Google Scholar] [CrossRef]
- Cignarella, F.; Filipello, F.; Bollman, B.; Cantoni, C.; Locca, A.; Mikesell, R.; Manis, M.; Ibrahim, A.; Deng, L.; Benitez, B.A.; et al. TREM2 activation on microglia promotes myelin debris clearance and remyelination in a model of multiple sclerosis. Acta Neuropathol. 2020, 140, 513–534. [Google Scholar] [CrossRef] [PubMed]
- Gratuze, M.; Chen, Y.; Parhizkar, S.; Jain, N.; Strickland, M.R.; Serrano, J.R.; Colonna, M.; Ulrich, J.D.; Holtzman, D.M. Activated microglia mitigate Aβ-associated tau seeding and spreading. J. Exp. Med. 2021, 218, e20210542. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences |
|---|---|
| IL-6 | F: CTGCAAGAGACTTCCATCCAG |
| R: AGTGGTATAGACAGGTCTGTTGG | |
| IL-4 | F: GGTCTCAACCCCCAGCTAGT |
| R: GCCGATGATCTCTCTCAAGTGAT | |
| IL-10 | F: CTTACTGACTGGCATGAGGATCA |
| R: GCAGCTCTAGGAGCATGTGG | |
| TNF-α | F: CAGGCGGTGCCTATGTCTC |
| R: CGATCACCCCGAAGTTCAGTAG | |
| TREM2 | F: CTGGAACCGTCACCATCACTC |
| R: CGAAACTCGATGACTCCTCGG | |
| iNOS | F: GTTCTCAGCCCAACAATACAAGA |
| R: GTGGACGGGTCGATGTCAC | |
| CD86 | F: TCAATGGGACTGCATATCTGCC |
| R: GCCAAAATACTACCAGCTCACT | |
| CD206 | F: CTCTGTTCAGCTATTGGACGC |
| R: TGGCACTCCCAAACATAATTTGA | |
| CD16 | F: AATGCACACTCTGGAAGCCAA |
| R: CACTCTGCCTGTCTGCAAAAG | |
| ARG-1 | F: CTCCAAGCCAAAGTCCTTAGAG |
| R: GGAGCTGTCATTAGGGACATCA | |
| IFN-β | F: AGCTCCAAGAAAGGACGAACA |
| R: GCCCTGTAGGTGAGGTTGAT | |
| GAPDH | F: AGGTCGGTGTGAACGGATTTG |
| R: GGGGTCGTTGATGGCAACA | |
| β-Actin | F: GTGACGTTGACATCCGTAAAGA |
| R: GTGACGTTGACATCCGTAAAGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Z.; Wang, H.; Qu, M.; Zhu, S.; Zhang, H.; Liao, Q.; Miao, C. Consecutive Injection of High-Dose Lipopolysaccharide Modulates Microglia Polarization via TREM2 to Alter Status of Septic Mice. Brain Sci. 2023, 13, 126. https://doi.org/10.3390/brainsci13010126
Qiu Z, Wang H, Qu M, Zhu S, Zhang H, Liao Q, Miao C. Consecutive Injection of High-Dose Lipopolysaccharide Modulates Microglia Polarization via TREM2 to Alter Status of Septic Mice. Brain Sciences. 2023; 13(1):126. https://doi.org/10.3390/brainsci13010126
Chicago/Turabian StyleQiu, Zhiyun, Huilin Wang, Mengdi Qu, Shuainan Zhu, Hao Zhang, Qingwu Liao, and Changhong Miao. 2023. "Consecutive Injection of High-Dose Lipopolysaccharide Modulates Microglia Polarization via TREM2 to Alter Status of Septic Mice" Brain Sciences 13, no. 1: 126. https://doi.org/10.3390/brainsci13010126