
Neuronal Cytoglobin in the Auditory Brainstem of Rat and Mouse: Distribution, Cochlear Projection, and Nitric Oxide Production


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Olivocochlear Neuronal Tracing
2.3. Tissue Processing and Immunofluorescence Incubations
2.4. Double Immunofluorescence Incubation of Mouse Brainstem Sections
2.5. Cell Quantifications and Statistical Analysis
2.6. Image Analysis
2.7. Control Studies
3. Results
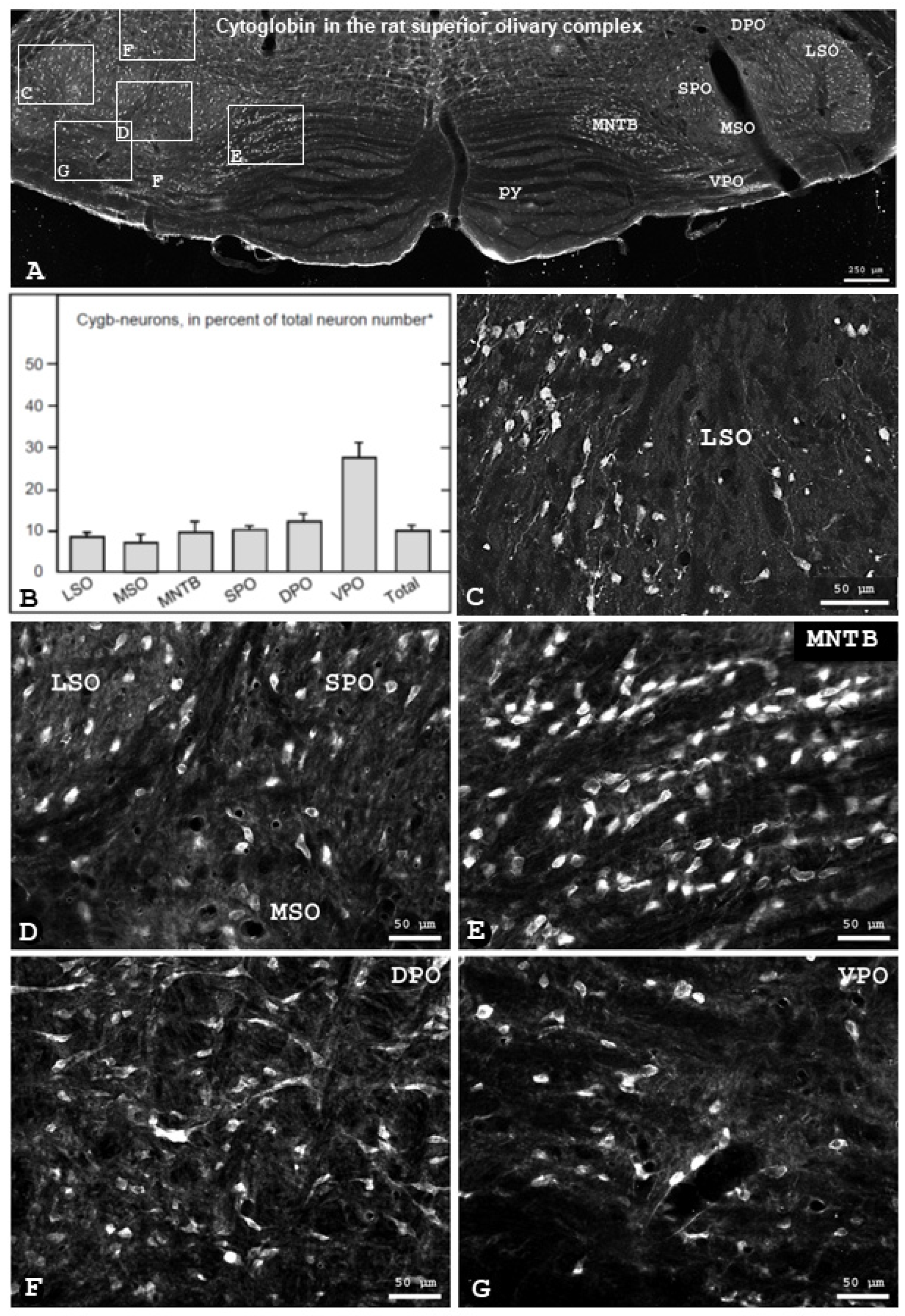
3.1. Localization of Cytoglobin Protein in Rat Brain
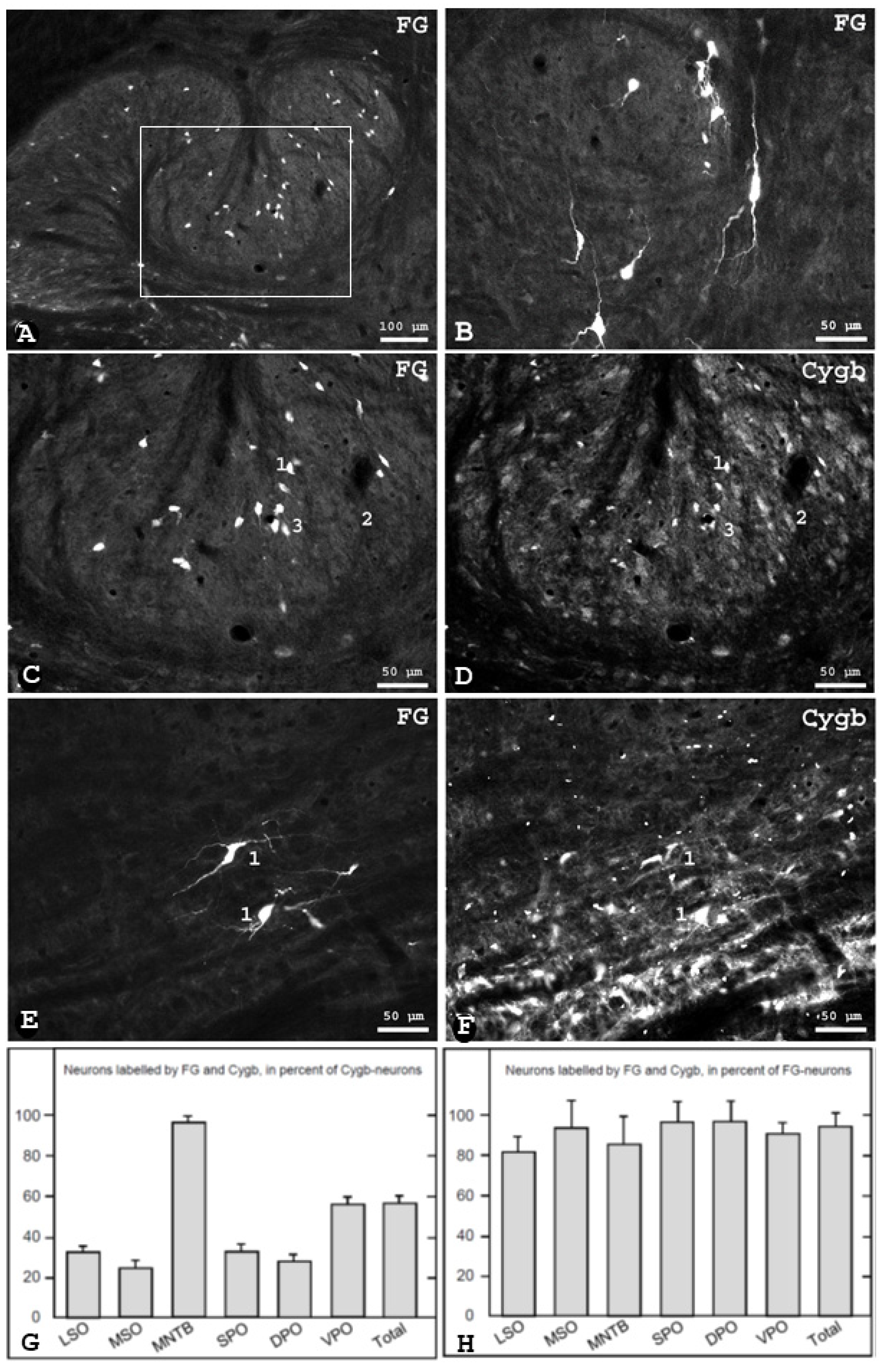
3.2. Cygb-Immunoreactivity in Identified Olivocochlear Neurons in the Rat
3.3. Cytoglobin in the Superior Olivary Complex of the Mouse
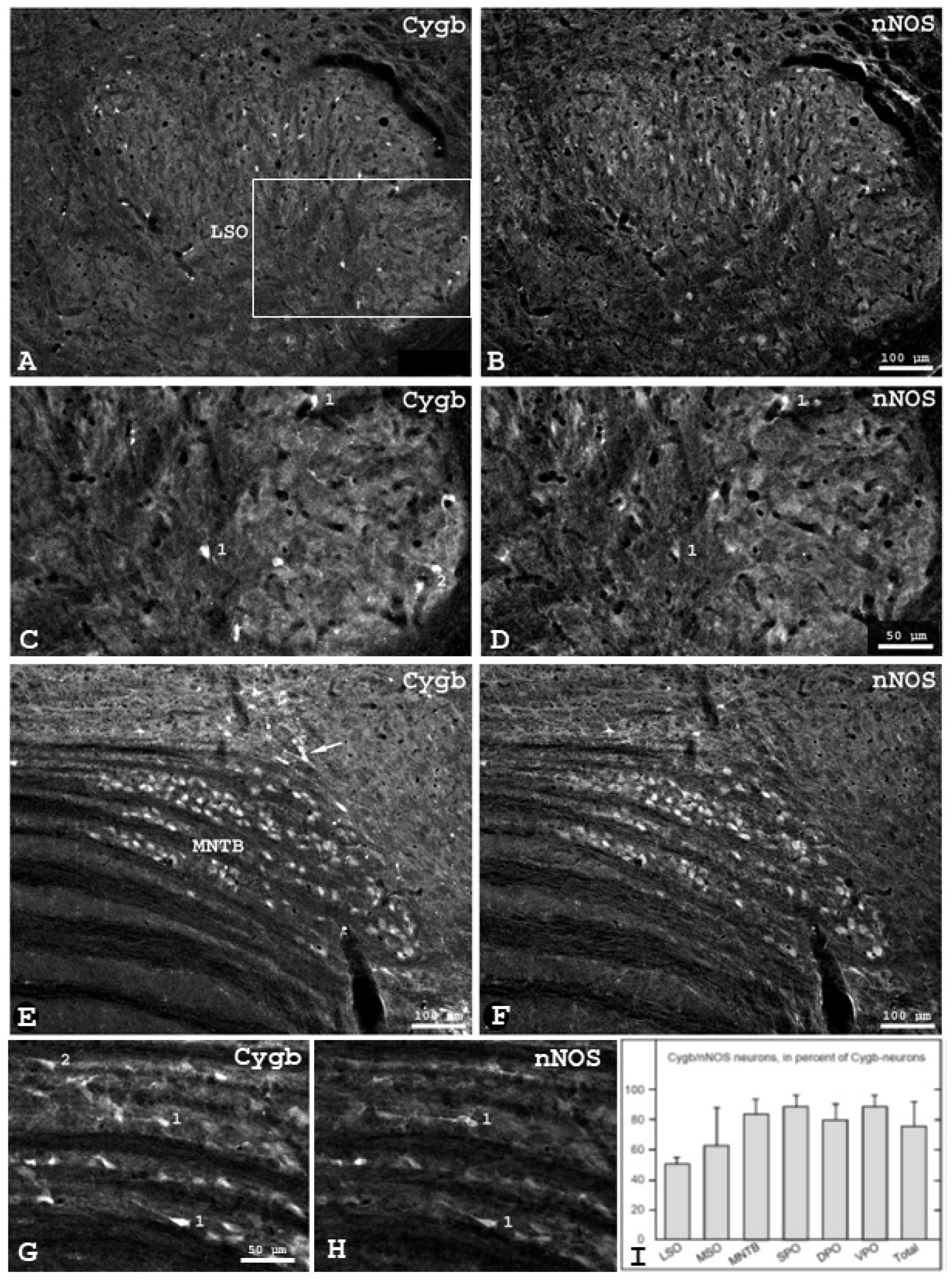
3.4. Colocalization of Cytoglobin and Neuronal Nitric Oxide Synthase in the Mouse SOC
4. Discussion
4.1. Distribution of Cytoglobin in the Superior Olivary Complex
4.2. Cytoglobin Expression by Rat Olivocochlear Neurons
4.3. Distribution of Cytoglobin in the Mouse Superior Olivary Complex
4.4. Colocalization of Cytoglobin and Neuronal Nitric Oxide-Synthase in the Mouse Superior Olivary Complex
4.5. Possible Function of Cytoglobin in Auditory Brainstem
5. Limitations of the Study
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Kawada, N.; Kristensen, D.B.; Asahina, K.; Nakatani, K.; Minamiyama, Y.; Seki, S.; Yoshizato, K. Characterization of a stellate cell activation-associated protein (STAP) with peroxidase activity found in rat hepatic stellate cells. J. Biol. Chem. 2001, 276, 25318–25323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmester, T.; Ebner, B.; Weich, B.; Hankeln, T. Cytoglobin: A novel globin type ubiquitously expressed in vertebrate tissues. Mol. Biol. Evol. 2002, 19, 416–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trent, J.T.; Hargrove, M.S. A ubiquitously expressed human hexacoordinate hemoglobin. J. Biol. Chem. 2002, 277, 19538–19545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, H.; Makino, M.; Sawai, H.; Kawada, N.; Yoshizato, K.; Shiro, Y. Structural basis of human cytoglobin for ligand binding. J. Mol. Biol. 2004, 339, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, K.; Okuyama, H.; Shimahara, Y.; Saeki, S.; Kim, D.H.; Nakajima, Y.; Seki, S.; Kawada, N.; Yoshizato, K. Cytoglobin/STAP, its unique localization in splanchnic fibroblast- like cells and function in organ fibrogenesis. Lab. Investig. 2004, 84, 91–101. [Google Scholar] [CrossRef]
- Schmidt, M.; Gerlach, F.; Avivi, A.; Laufs, T.; Wystub, S.; Simpson, J.C.; Nevo, E.; Saaler-Reinhardt, S.; Reuss, S.; Hankeln, T.; et al. Cytoglobin Is a Respiratory Protein in Connective Tissue and Neurons, Which Is Up-regulated by Hypoxia. J. Biol. Chem. 2004, 279, 8063–8069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keppner, A.; Maric, D.; Correia, M.; Koay, T.W.; Orlando, I.M.C.; Vinogradov, S.N.; Hoogewijs, D. Lessons from the post-genomic era: Globin diversity beyond oxygen binding and transport. Redox Biol. 2020, 37, 101687. [Google Scholar] [CrossRef]
- Burmester, T.; Hankeln, T. Function and evolution of vertebrate globins. Acta Physiol. 2014, 211, 501–514. [Google Scholar] [CrossRef]
- Ascenzi, P.; Gustincich, S.; Marino, M. Mammalian nerve globins in search of functions. IUBMB Life 2014, 66, 268–276. [Google Scholar] [CrossRef]
- Xie, L.K.; Yang, S.H. Brain globins in physiology and pathology. Med. Gas Res. 2016, 6, 154–163. [Google Scholar] [CrossRef]
- Avivi, A.; Gerlach, F.; Joel, A.; Reuss, S.; Burmester, T.; Nevo, E.; Hankeln, T. Neuroglobin, cytoglobin, and myoglobin contribute to hypoxia adaptation of the subterranean mole rat Spalax. Proc. Natl. Acad. Sci. USA 2010, 107, 21570–21575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hundahl, C.A.; Elfving, B.; Muller, H.K.; Hay-Schmidt, A.; Wegener, G. A gene-environment study of cytoglobin in the human and rat hippocampus. PLoS ONE 2013, 8, e63288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hundahl, C.A.; Fahrenkrug, J.; Hannibal, J. Neurochemical phenotype of cytoglobin-expressing neurons in the rat hippocampus. Biomed. Rep. 2014, 2, 620–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.; Laufs, T.; Reuss, S.; Hankeln, T.; Burmester, T. Divergent distribution of cytoglobin and neuroglobin in the murine eye. Neurosci. Lett. 2005, 374, 207–211. [Google Scholar] [CrossRef]
- Mammen, P.P.; Shelton, J.M.; Ye, Q.; Kanatous, S.B.; McGrath, A.J.; Richardson, J.A.; Garry, D.J. Cytoglobin is a stress-responsive hemoprotein expressed in the developing and adult brain. J. Histochem. Cytochem. 2006, 54, 1349–1361. [Google Scholar] [CrossRef] [Green Version]
- Hundahl, C.A.; Allen, G.C.; Hannibal, J.; Kjaer, K.; Rehfeld, J.F.; Dewilde, S.; Nyengaard, J.R.; Kelsen, J.; Hay-Schmidt, A. Anatomical characterization of cytoglobin and neuroglobin mRNA and protein expression in the mouse brain. Brain Res. 2010, 1331, 58–73. [Google Scholar] [CrossRef]
- Reuss, S.; Wystub, S.; Disque-Kaiser, U.; Hankeln, T.; Burmester, T. Distribution of Cytoglobin in the Mouse Brain. Front. Neuroanat. 2016, 10, 47. [Google Scholar] [CrossRef] [Green Version]
- Guinan, J.J., Jr. Olivocochlear efferents: Their action, effects, measurement and uses, and the impact of the new conception of cochlear mechanical responses. Hear. Res. 2018, 362, 38–47. [Google Scholar] [CrossRef]
- Moore, D.R. Anatomy and physiology of binaural hearing. Audiol. Off. Organ Int. Soc. Audiol. 1991, 30, 125–134. [Google Scholar] [CrossRef]
- McLean, I.W.; Nakane, P.K. Periodate-lysine-paraformaldehyde fixative. A new fixation for immunoelectron microscopy. J. Histochem. Cytochem. 1974, 22, 1077–1083. [Google Scholar] [CrossRef]
- Reuss, S.; Kühn, I.; Windoffer, R.; Riemann, R. Neurochemistry of identified motoneurons of the tensor tympani muscle in rat middle ear. Hear. Res. 2009, 248, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Abercrombie, M. Estimation of nuclear population from microtome sections. Anat. Rec. 1946, 94, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 7th ed.; Academic Press: San Diego, CA, USA, 2013. [Google Scholar]
- Malmierca, M.S.; Ryugo, D.K. Auditory System. In The Mouse Nervous System, 1st ed.; Watson, C., Paxinos, G., Puelles, L., Eds.; Academic Press: Amsterdam, The Netherlands, 2012; pp. 607–645. [Google Scholar]
- Malmierca, M.S. Auditory System. In The Rat Nervous System, 4th ed.; Paxinos, G., Ed.; Academic Press: Amsterdam, The Netherlands, 2015; pp. 863–946. [Google Scholar]
- Riemann, R.; Reuss, S. Nitric oxide synthase in identified olivocochlear projection neurons in rat and guinea pig. Hear. Res. 1999, 135, 181–189. [Google Scholar] [CrossRef]
- Vetter, D.E.; Mugnaini, E. Distribution and dendritic features of three groups of rat olivocochlear neurons. A study with two retrograde cholera toxin tracers. Anat. Embryol. 1992, 185, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mitz, S.A.; Reuss, S.; Folkow, L.P.; Blix, A.S.; Ramirez, J.M.; Hankeln, T.; Burmester, T. When the brain goes diving: Glial oxidative metabolism may confer hypoxia tolerance to the seal brain. Neuroscience 2009, 163, 552–560. [Google Scholar] [CrossRef]
- Li, R.C.; Lee, S.K.; Pouranfar, F.; Brittian, K.R.; Clair, H.B.; Row, B.W.; Wang, Y.; Gozal, D. Hypoxia differentially regulates the expression of neuroglobin and cytoglobin in rat brain. Brain Res. 2006, 1096, 173–179. [Google Scholar] [CrossRef]
- Itoh, R.; Fujita, K.; Mu, A.; Kim, D.H.; Tai, T.T.; Sagami, I.; Taketani, S. Imaging of heme/hemeproteins in nucleus of the living cells expressing heme-binding nuclear receptors. FEBS Lett. 2013, 587, 2131–2136. [Google Scholar] [CrossRef] [Green Version]
- Reuss, S.; Disque-Kaiser, U.; Antoniou-Lipfert, P.; Najaf Gholi, M.; Riemann, E.; Riemann, R. Neurochemistry of olivocochlear neurons in the hamster. Anat. Rec. 2009, 292, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Reuss, S.; Banica, O.; Elgurt, M.; Mitz, S.; Disque-Kaiser, U.; Riemann, R.; Hill, M.; Jaquish, D.V.; Koehrn, F.J.; Burmester, T.; et al. Neuroglobin expression in the mammalian auditory system. Mol. Neurobiol. 2016, 53, 1461–1477. [Google Scholar] [CrossRef] [Green Version]
- Robertson, D.; Harvey, A.R.; Cole, K.S. Postnatal development of the efferent innervation of the rat cochlea. Dev. Brain Res. 1989, 47, 197–207. [Google Scholar] [CrossRef]
- Aschoff, A.; Ostwald, J. Distribution of cochlear efferents and olivo-collicular neurons in the brainstem of rat and guinea pig. A double labeling study with fluorescent tracers. Exp. Brain Res. 1988, 71, 241–251. [Google Scholar] [CrossRef] [PubMed]
- White, J.S.; Warr, W.B. The dual origins of the olivocochlear bundle in the albino rat. J. Comp. Neurol. 1983, 219, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Ruel, J.; Wang, J.; Rebillard, G.; Eybalin, M.; Lloyd, R.; Pujol, R.; Puel, J.L. Physiology, pharmacology and plasticity at the inner hair cell synaptic complex. Hear. Res. 2007, 227, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Schofield, B.R.; Beebe, N.L. The Efferent Auditory System: Central Pathways That Modulate Peripheral Input. In The Senses: A Comprehensive Reference, 2nd ed.; Grothe, B., Fritzsch, B., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 2, pp. 501–516. [Google Scholar]
- Warr, W.B.; Boche, J.B.; Neely, S.T. Efferent innervation of the inner hair cell region: Origins and terminations of two lateral olivocochlear systems. Hear. Res. 1997, 108, 89–111. [Google Scholar] [CrossRef]
- Sánchez-González, M.A.; Warr, W.B.; López, D.E. Anatomy of olivocochlear neurons in the hamster studied with FluoroGold. Hear. Res. 2003, 185, 65–76. [Google Scholar] [CrossRef]
- Hill, J.C.; Prasher, D.K.; Luxon, L.M. Evidence for efferent effects on auditory afferent activity, and their functional relevance. Clin. Otolaryngol. Allied Sci. 1997, 22, 394–402. [Google Scholar] [CrossRef]
- Tanaka, C.; Coling, D.E.; Manohar, S.; Chen, G.D.; Hu, B.H.; Salvi, R.; Henderson, D. Expression pattern of oxidative stress and antioxidant defense-related genes in the aging Fischer 344/NHsd rat cochlea. Neurobiol Aging 2012, 33, 1842.e1–1842.e14. [Google Scholar] [CrossRef] [Green Version]
- Dalla Costa, I.; Buchanan, C.N.; Zdradzinski, M.D.; Sahoo, P.K.; Smith, T.P.; Thames, E.; Kar, A.N.; Twiss, J.L. The functional organization of axonal mRNA transport and translation. Nat. Rev. Neurosci. 2021, 22, 77–91. [Google Scholar] [CrossRef]
- Mink, J.W.; Blumenschine, R.J.; Adams, D.B. Ratio of central nervous system to body metabolism in vertebrates: Its constancy and functional basis. Am. J. Physiol. 1981, 241, R203–R212. [Google Scholar] [CrossRef]
- Jackson Laboratory Resource Manual. Available online: https://phenome.jax.org/measureset/22701 (accessed on 26 December 2022).
- Kulesza, R.J.; Viñuela, A.; Saldaña, E.; Berrebi, A.S. Unbiased stereological estimates of neuron number in subcortical auditory nuclei of the rat. Hear. Res. 2002, 168, 12–24. [Google Scholar] [CrossRef]
- Burmester, T.; Hankeln, T. Neuroglobin and other nerve haemoglobins. In Dioxygen Binding and Sensing Proteins. Protein Reviews; Bolognesi, M., di Prisco, G., Verde, C., Eds.; Springer: Milano, Italy, 2008; pp. 211–222. [Google Scholar]
- Fessenden, J.D.; Coling, D.E.; Schacht, J. Detection and characterization of nitric oxide synthase in the mammalian cochlea. Brain Res. 1994, 668, 9–15. [Google Scholar] [CrossRef]
- Rodrigo, J.; Springall, D.R.; Uttenthal, O.; Bentura, M.L.; Abadia-Molina, F.; Riveros-Moreno, V.; Martínez-Murillo, R.; Polak, J.M.; Moncada, S. Localization of nitric oxide synthase in the adult rat brain. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 1994, 345, 175–221. [Google Scholar] [CrossRef]
- Reuss, S.; Riemann, R. Distribution and projections of nitric oxide synthase neurons in the rodent superior olivary complex. Microsc. Res. Tech. 2000, 51, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, U.R.; Helling, K. Nitric oxide—A versatile key player in cochlear function and hearing disorders. Nitric Oxide 2012, 27, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Halligan, K.E.; Jourd’heuil, F.L.; Jourd’heuil, D. Cytoglobin is expressed in the vasculature and regulates cell respiration and proliferation via nitric oxide dioxygenation. J. Biol. Chem. 2009, 284, 8539–8547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, A.M.; Cook, M.R.; Gardner, P.R. Nitric-oxide dioxygenase function of human cytoglobin with cellular reductants and in rat hepatocytes. J. Biol. Chem. 2010, 285, 23850–23857. [Google Scholar] [CrossRef] [Green Version]
- Luyckx, E.; Van Acker, Z.P.; Ponsaerts, P.; Dewilde, S. Neuroglobin Expression Models as a Tool to Study Its Function. Oxidative Med. Cell. Longev. 2019, 2019, 5728129. [Google Scholar] [CrossRef] [Green Version]
- Nowotny, M.; Kiefer, L.; Andre, D.; Fabrizius, A.; Hankeln, T.; Reuss, S. Hearing Without Neuroglobin. Neuroscience 2017, 366, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.A. Novel roles for collagens in wiring the vertebrate nervous system. Curr. Opin. Cell Biol. 2008, 20, 508–513. [Google Scholar] [CrossRef]
- Wakabayashi, T. Transmembrane Collagens in Neuromuscular Development and Disorders. Front. Mol. Neurosci. 2020, 13, 635375. [Google Scholar] [CrossRef]
- Chang, D.; Brown, Q.; Tsui, G.; He, Y.; Liu, J.; Shi, L.; Rodríguez-Contreras, A. Distinct Cellular Profiles of Hif1a and Vegf mRNA Localization in Microglia, Astrocytes and Neurons during a Period of Vascular Maturation in the Auditory Brainstem of Neonate Rats. Brain Sci. 2021, 11, 944. [Google Scholar] [CrossRef] [PubMed]
- Canis, M.; Bertlich, M. Cochlear Capillary Pericytes. Adv. Exp. Med. Biol. 2019, 1122, 115–123. [Google Scholar] [CrossRef] [PubMed]
- White, P.M. Perspectives on Human Hearing Loss, Cochlear Regeneration, and the Potential for Hearing Restoration Therapies. Brain Sci. 2020, 10, 756. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reuss, S.; Balmaceda, D.; Elgurt, M.; Riemann, R. Neuronal Cytoglobin in the Auditory Brainstem of Rat and Mouse: Distribution, Cochlear Projection, and Nitric Oxide Production. Brain Sci. 2023, 13, 107. https://doi.org/10.3390/brainsci13010107
Reuss S, Balmaceda D, Elgurt M, Riemann R. Neuronal Cytoglobin in the Auditory Brainstem of Rat and Mouse: Distribution, Cochlear Projection, and Nitric Oxide Production. Brain Sciences. 2023; 13(1):107. https://doi.org/10.3390/brainsci13010107
Chicago/Turabian StyleReuss, Stefan, Daniel Balmaceda, Mirra Elgurt, and Randolf Riemann. 2023. "Neuronal Cytoglobin in the Auditory Brainstem of Rat and Mouse: Distribution, Cochlear Projection, and Nitric Oxide Production" Brain Sciences 13, no. 1: 107. https://doi.org/10.3390/brainsci13010107