
Effect of Electroacupuncture at Fengchi on Facial Allodynia, Microglial Activation, and Microglia–Neuron Interaction in a Rat Model of Migraine


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Dura Mater Electrode Implantation
2.3. Experimental Design
2.3.1. Experiment 1
2.3.2. Experiment 2
2.4. Establishment of Recurrent Migraine Rat Model
2.5. EA Intervention
2.6. Facial Mechanical Threshold by Electronic von Frey
2.7. Immunofluorescence and Image Analyses
2.8. Western Blot
2.9. Real-Time PCR
| β-actin | Forward primer 5, -CCCATCTATGAGGGTTACGC-3; |
| Reverse primer 5, -TTTAATGTCACGCACGATTTC-3; | |
| Iba-1 | Forward primer 5, -CTCCGAGGAGACGTTCAGTT-3; |
| Reverse primer 5, -TTGGCTTCTGGTGTTCTTTGT-3; | |
| TNF-α | Forward primer 5, -GGGCCACCACGCTCTTCTGT-3; |
| Reverse primer 5, -GGCTACGGGCTTGTCACTCG-3; | |
| IL-1β | Forward primer 5, -AGGCAGTGTCACTCATTGTGG-3; |
| Reverse primer 5, -TAGCAGGTCGTCATCATCCC-3; |
2.10. Antibodies
- Rabbit anti-Iba-1(Iba-1 as microglial marker, 1:400,019-19741, Wako Chemicals, VA, USA);
- Mouse anti-c-Fos (1:200, ab190289, Abcam, Cambridge, UK);
- Mouse anti-TNFRII (1:200, SC-8041, Santa Cruz, CA, USA);
- Mouse anti-IL-1RI (1:400, SC-393998, Santa Cruz, CA, USA);
- Rabbit anti-NeuN (neuronal marker, 1:400, ab236870, Abcam, Cambridge, UK).
- 2.
- Rabbit anti-TNF-α (1:500, bs-2081R, Bioss, Beijing, China);
- Mouse anti-TNF-αRII (1:500, Sc-8041, Santa Cruz, CA, USA);
- Rabbit anti-IL-1β(1:500, bs-6319R, Bioss, Beijing, China);
- Mouse anti-IL-1βRI (1:500, Sc-393998, Santa Cruz, CA, USA);
- Mouse anti-Iba-1 (microglial calcium-binding protein, 1:400, Sc-32725, Santa Cruz, CA, USA).
2.11. Statistical Analysis
3. Results
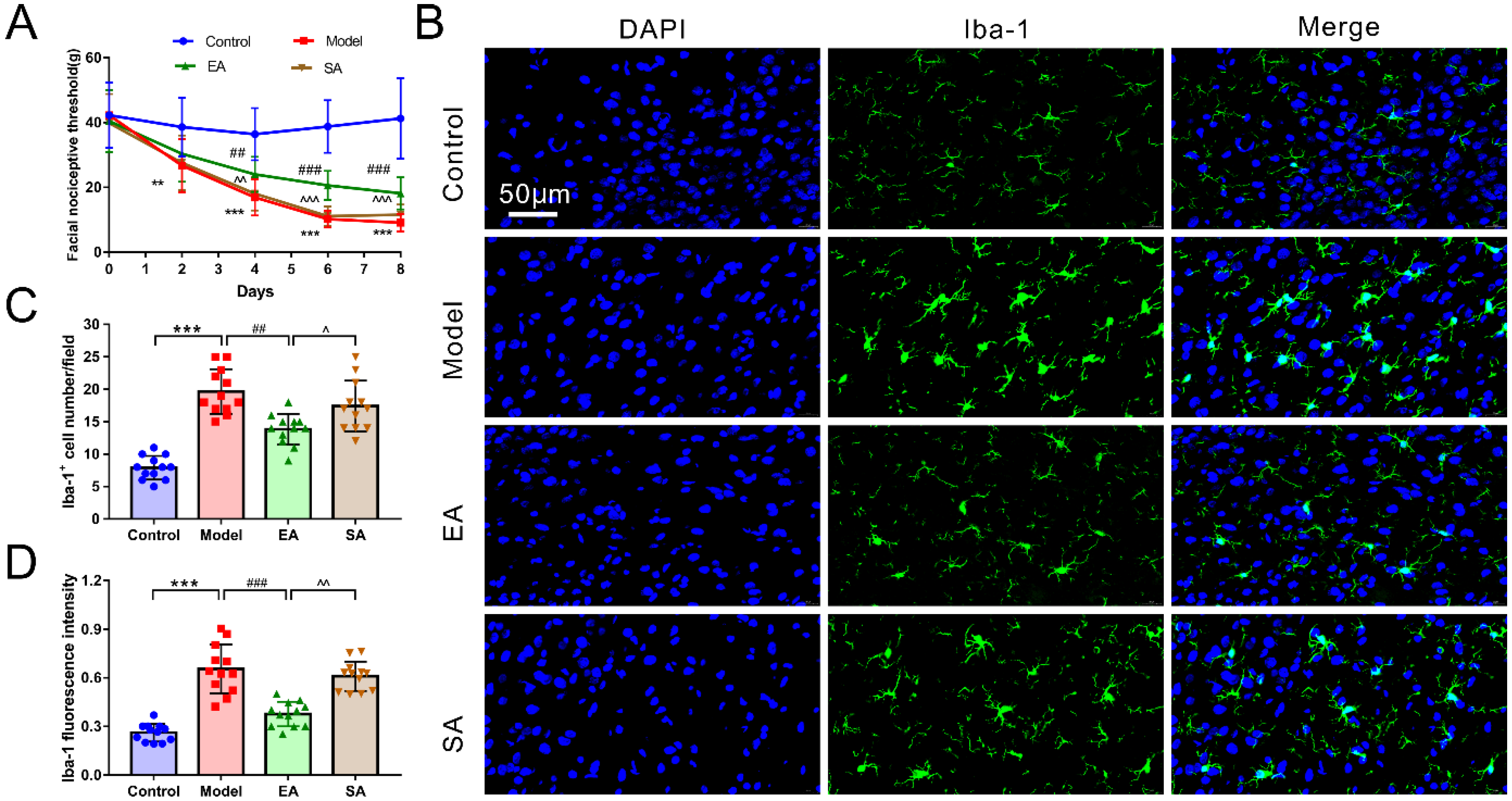
3.1. EA Attenuated DES-Induced Facial Mechanical Threshold
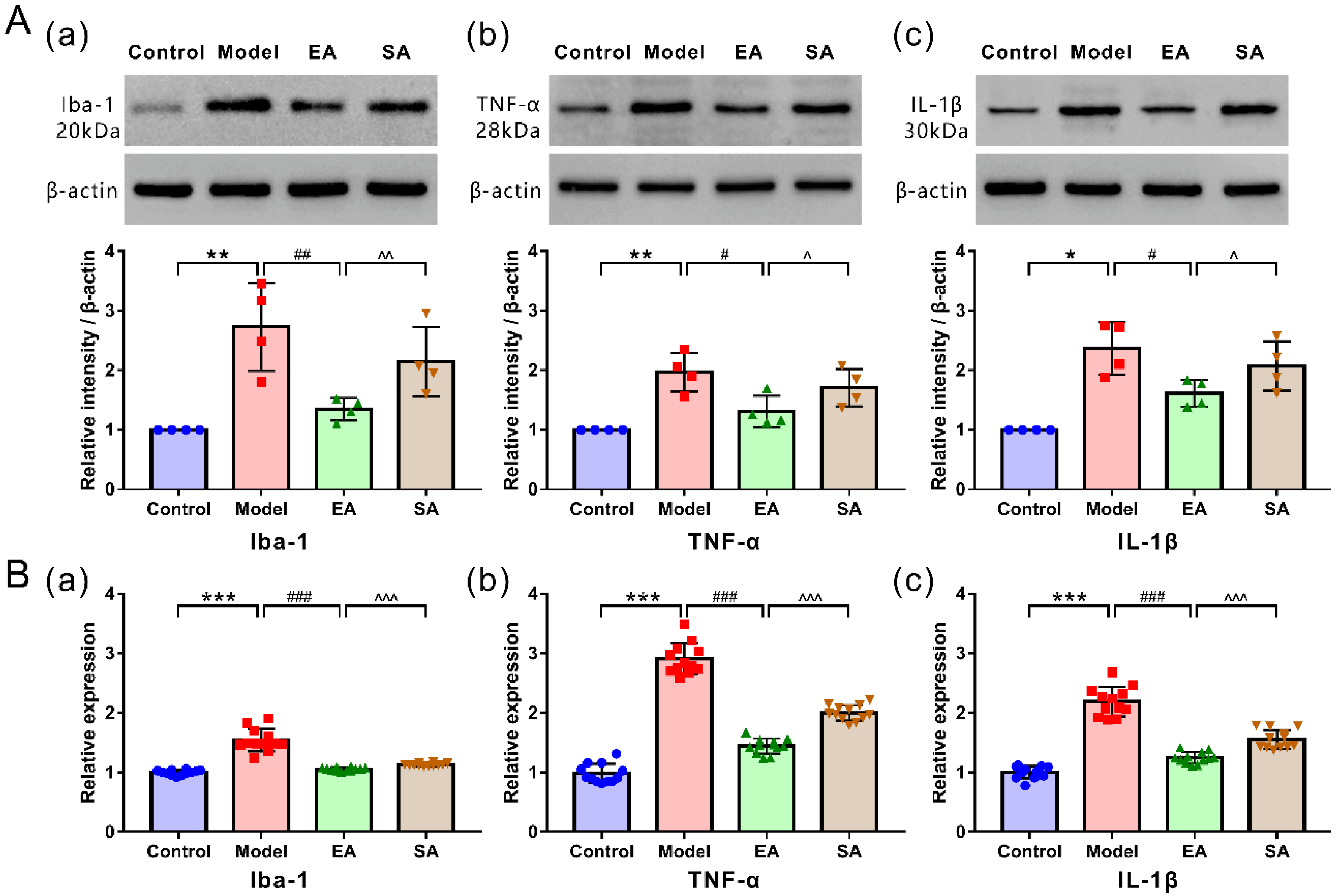
3.2. EA Attenuated DES-Induced Microglial Activation and Inflammatory Factor Release in the TNC
3.3. EA Attenuated DES-Induced High Expression of Inflammatory Factor Receptors in TNC Neurons
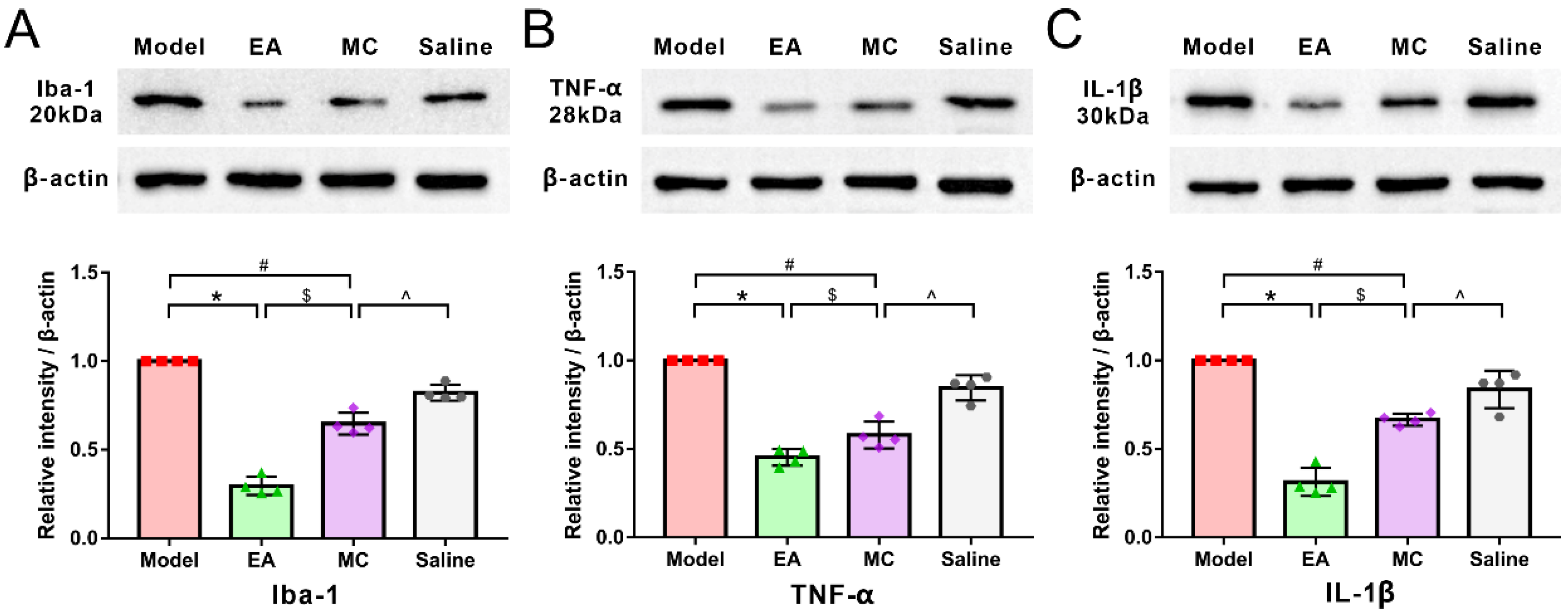
3.4. Minocycline Has a Partial Acupuncture-Like Effect in Migraine Central Sensitization: Relieving DES-Induced Facial Mechanical Threshold and TNC Activation
4. Discussion
4.1. EA at GB20 Alleviated Migraine Central Sensitization by Inhibiting Microglial Activation
4.2. GB20 Can Regulate the Expression of Inflammatory Factors Receptors on Neurons
4.3. Minocycline, a Microglia Inhibitor, Can Exert Part of Acupuncture-like Effects
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guven, H.; Cilliler, A.E.; Comoglu, S.S. Cutaneous allodynia in patients with episodic migraine. Neruol. Sci. 2013, 34, 1397–1402. [Google Scholar] [CrossRef] [PubMed]
- Louter, M.A.; Bosker, J.E.; van Oosterhout, W.P.; van Zwet, E.W.; Zitman, F.G.; Ferrari, M.D.; Terwindt, G.M. Cutaneous allodynia as a predictor of migraine chronification. Brain 2013, 136, 3489–3496. [Google Scholar] [CrossRef] [PubMed]
- Burstein, R.; Collins, B.; Jakubowski, M. Defeating migraine pain with triptans: A race against the development of cutaneous allodynia. Ann. Neurol. 2004, 55, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Lipton, R.B.; Bigal, M.E.; Ashina, S.; Burstein, R.; Silberstein, S.; Reed, M.L.; Serrano, D.; Stewart, W.F. Cutaneous allodynia in the migraine population. Ann. Neurol. 2008, 63, 148–158. [Google Scholar] [CrossRef]
- Mathew, P.G.; Cutrer, F.M.; Garza, I. A touchy subject: An assessment of cutaneous allodynia in a chronic migraine population. J. Pain Res. 2016, 9, 101–104. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, X.; Zhong, D.; Luo, L.; Yang, H.; Zhou, J.; He, M.; Shi, L.; Li, J.; Zheng, H.; et al. Effectiveness and safety of acupuncture for migraine: An overview of systematic reviews. Pain Res. Manag. 2020, 2020, 3825617. [Google Scholar] [CrossRef]
- Boyer, N.; Dallel, R.; Artola, A.; Monconduit, L. General trigeminospinal central sensitization and impaired descending pain inhibitory controls contribute to migraine progression. Pain 2014, 155, 1196–1205. [Google Scholar] [CrossRef]
- Edelmayer, R.M.; Vanderah, T.W.; Majuta, L.; Zhang, E.T.; Fioravanti, B.; De Felice, M.; Chichorro, J.G.; Ossipov, M.H.; King, T.; Lai, J.; et al. Medullary pain facilitating neurons mediate allodynia in headache-related pain. Ann. Neurol. 2009, 65, 184–193. [Google Scholar] [CrossRef]
- Conti, P.; D’Ovidio, C.; Conti, C.; Gallenga, C.E.; Lauritano, D.; Caraffa, A.; Kritas, S.K.; Ronconi, G. Progression in migraine: Role of mast cells and proinflammatory and anti-inflammatory cytokines. Eur. J. Pharmacol. 2019, 844, 87–94. [Google Scholar] [CrossRef]
- Sommer, C.; Leinders, M.; Uceyler, N. Inflammation in the pathophysiology of neuropathic pain. Pain 2018, 159, 595–602. [Google Scholar] [CrossRef]
- Tozaki-Saitoh, H.; Tsuda, M. Microglia-neuron interactions in the models of neuropathic pain. Biochem. Pharmacol. 2019, 169, 113614. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M. Microglia-Mediated Regulation of Neuropathic Pain: Molecular and Cellular Mechanisms. Biol. Pharm. Bull. 2019, 42, 1959–1968. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Tsuda, M. Microglia in neuropathic pain: Cellular and molecular mechanisms and therapeutic potential. Nat. Rev. Neurosci. 2018, 19, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Guo, W.; Zou, S.; Ren, K.; Dubner, R. Supraspinal glial-neuronal interactions contribute to descending pain facilitation. J. Neurosci. 2008, 28, 10482–10495. [Google Scholar] [CrossRef]
- Fried, N.T.; Maxwell, C.R.; Elliott, M.B.; Oshinsky, M.L. Region-specific disruption of the blood-brain barrier following repeated inflammatory dural stimulation in a rat model of chronic trigeminal allodynia. Cephalalgia 2018, 38, 674–689. [Google Scholar] [CrossRef]
- He, W.; Long, T.; Pan, Q.; Zhang, S.; Zhang, Y.; Zhang, D.; Qin, G.; Chen, L.; Zhou, J. Microglial NLRP3 inflammasome activation mediates IL-1β release and contributes to central sensitization in a recurrent nitroglycerin-induced migraine model. J. Neuroinflamm. 2019, 16, 78. [Google Scholar] [CrossRef]
- Raghavendra, V.; Tanga, F.; DeLeo, J.A. Inhibition of microglial activation attenuates the development but not existing hypersensitivity in a rat model of neuropathy. J. Pharmacol. Exp. Ther. 2003, 306, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Wieseler, J.; Ellis, A.; McFadden, A.; Stone, K.; Brown, K.; Cady, S.; Bastos, L.F.; Sprunger, D.; Rezvani, N.; Johnson, K.; et al. Supradural inflammatory soup in awake and freely moving rats induces facial allodynia that is blocked by putative immune modulators. Brain Res. 2017, 1664, 87–94. [Google Scholar] [CrossRef]
- Lai, H.C.; Lin, Y.W.; Hsieh, C.L. Acupuncture-Analgesia-Mediated Alleviation of Central Sensitization. Evid. Based Complement. Altern. Med. 2019, 2019, 6173412. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, X.; Guo, J.; Liu, H.; Zhang, Y.; Liu, C.; Yi, J.; Wang, L.; Zhao, J.; Li, S. Efficacy of acupuncture for migraine prophylaxis: A single-blinded, double-dummy, randomized controlled trial. Pain 2011, 152, 1864–1871. [Google Scholar] [CrossRef]
- Zhang, N.; Houle, T.; Hindiyeh, N.; Aurora, S.K. Systematic Review: Acupuncture vs. Standard Pharmacological Therapy for Migraine Prevention. Headache 2020, 60, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Wang, D.; Fan, G. Acupoint selection for acute migraine treated with acupuncture. Chin. Acupunct. Moxibustion 2018, 38, 1183–1188. [Google Scholar]
- Pei, P.; Liu, L.; Zhao, L.; Cui, Y.; Qu, Z.; Wang, L. Effect of electroacupuncture pretreatment at GB20 on behaviour and the descending pain modulatory system in a rat model of migraine. Acupunct. Med. 2016, 34, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Pei, P.; Liu, L.; Zhao, L.; Qu, Z.; Tang, C.; Wang, L.; Yang, W. Electroacupuncture exerts an anti-migraine effect via modulation of the 5-HT7 receptor in the conscious rat. Acupunct. Med. 2019, 37, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Liu, L.; Pei, P.; Qu, Z.; Zhu, Y.; Wang, L. Electroacupuncture at Fengchi (GB20) inhibits calcitonin gene-related peptide expression in the trigeminovascular system of a rat model of migraine. Neural Regen. Res. 2017, 12, 804–811. [Google Scholar]
- Burstein, R.; Yarnitsky, D.; Goor-Aryeh, I.; Ransil, B.J.; Bajwa, Z.H. An association between migraine and cutaneous allodynia. Ann. Neurol. 2000, 47, 614–624. [Google Scholar] [CrossRef]
- Noseda, R.; Burstein, R. Migraine pathophysiology: Anatomy of the trigeminovascular pathway and associated neurological symptoms, cortical spreading depression, sensitization, and modulation of pain. Pain 2013, 154, S44–S53. [Google Scholar] [CrossRef]
- Burstein, R.; Noseda, R.; Borsook, D. Migraine: Multiple processes, complex pathophysiology. J. Neurosci. 2015, 35, 6619–6629. [Google Scholar] [CrossRef]
- Tsuda, M. Microglia in the CNS and Neuropathic Pain. Adv. Exp. Med. Biol. 2018, 1099, 77–91. [Google Scholar]
- Gwak, Y.S.; Hulsebosch, C.E.; Leem, J.W. Neuronal-Glial Interactions Maintain Chronic Neuropathic Pain after Spinal Cord Injury. Neural Plast. 2017, 2017, 2480689. [Google Scholar] [CrossRef]
- Magni, G.; Boccazzi, M.; Bodini, A.; Abbracchio, M.P.; van den Maagdenberg, A.M.; Ceruti, S. Basal astrocyte and microglia activation in the central nervous system of Familial Hemiplegic Migraine Type I mice. Cephalalgia 2019, 39, 1809–1817. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, C.; Burstein, R. Sensitization of the trigeminovascular pathway: Perspective and implications to migraine pathophysiology. J. Clin. Neurol. 2012, 8, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Rozen, T.; Swidan, S.Z. Elevation of CSF tumor necrosis factor alpha levels in new daily persistent headache and treatment refractory chronic migraine. Headache 2007, 47, 1050–1055. [Google Scholar] [CrossRef] [PubMed]
- Hazey, M.A.; Van Norman, A.J.; Armistead, D.L. Melkersson-Rosenthal Syndrome with migraine-like headaches treated with minocycline: A case report and review of the literature. W. Va. Med. J. 2009, 105, 15–17. [Google Scholar]
- Ledeboer, A.; Sloane, E.M.; Milligan, E.D.; Frank, M.G.; Mahony, J.H.; Maier, S.F.; Watkins, L.R. Minocycline attenuates mechanical allodynia and proinflammatory cytokine expression in rat models of pain facilitation. Pain 2005, 115, 71–83. [Google Scholar] [CrossRef]
- Aczél, T.; Kecskés, A.; Kun, J.; Szenthe, K.; Bánáti, F.; Szathmary, S.; Herczeg, R.; Urbán, P.; Gyenesei, A.; Gaszner, B.; et al. Hemokinin-1 gene expression is upregulated in trigeminal ganglia in an inflammatory orofacial pain model: Potential role in peripheral sensitization. Int. J. Mol. Sci. 2020, 21, 2938. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pei, P.; Cui, S.; Zhang, S.; Hu, S.; Wang, L.; Yang, W. Effect of Electroacupuncture at Fengchi on Facial Allodynia, Microglial Activation, and Microglia–Neuron Interaction in a Rat Model of Migraine. Brain Sci. 2022, 12, 1100. https://doi.org/10.3390/brainsci12081100
Pei P, Cui S, Zhang S, Hu S, Wang L, Yang W. Effect of Electroacupuncture at Fengchi on Facial Allodynia, Microglial Activation, and Microglia–Neuron Interaction in a Rat Model of Migraine. Brain Sciences. 2022; 12(8):1100. https://doi.org/10.3390/brainsci12081100
Chicago/Turabian StylePei, Pei, Shengwei Cui, Shuaishuai Zhang, Sheng Hu, Linpeng Wang, and Wenming Yang. 2022. "Effect of Electroacupuncture at Fengchi on Facial Allodynia, Microglial Activation, and Microglia–Neuron Interaction in a Rat Model of Migraine" Brain Sciences 12, no. 8: 1100. https://doi.org/10.3390/brainsci12081100