
Memantine Disrupts Motor Coordination through Anxiety-like Behavior in CD1 Mice

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Memantine Administration
2.2. Behavioral Testing
2.2.1. Rotarod Test
2.2.2. Contextual and Cued Fear Conditioning
2.2.3. Elevated Plus Maze
2.2.4. Tail Suspension Test
2.3. Electrophysiology
2.3.1. Acute Slice Preparation
2.3.2. Field Excitatory Postsynaptic Potential (fEPSP) Recording
2.4. Statistical Analysis
3. Results
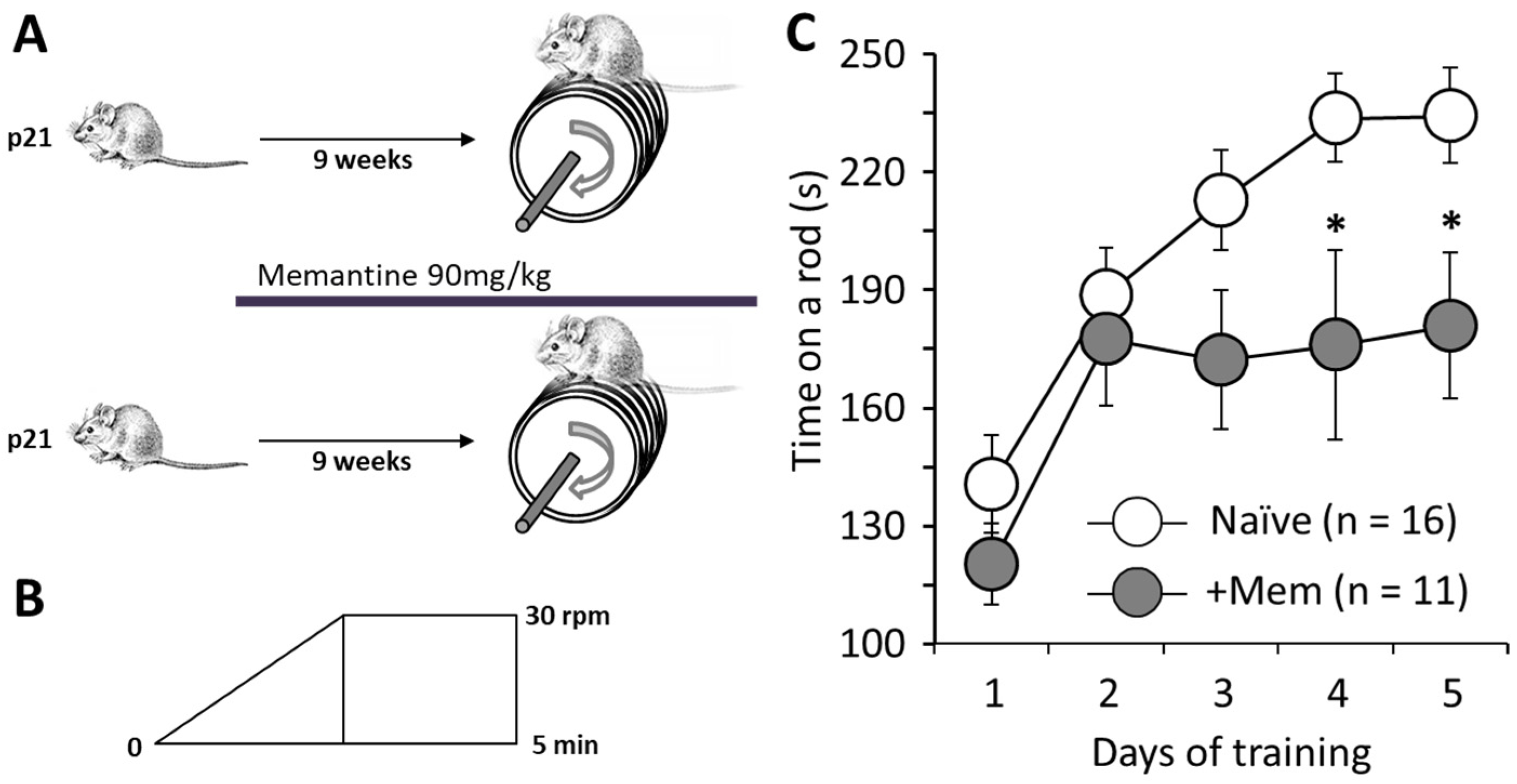
3.1. Chronic Administration of Memantine Affects Rotarod Performance
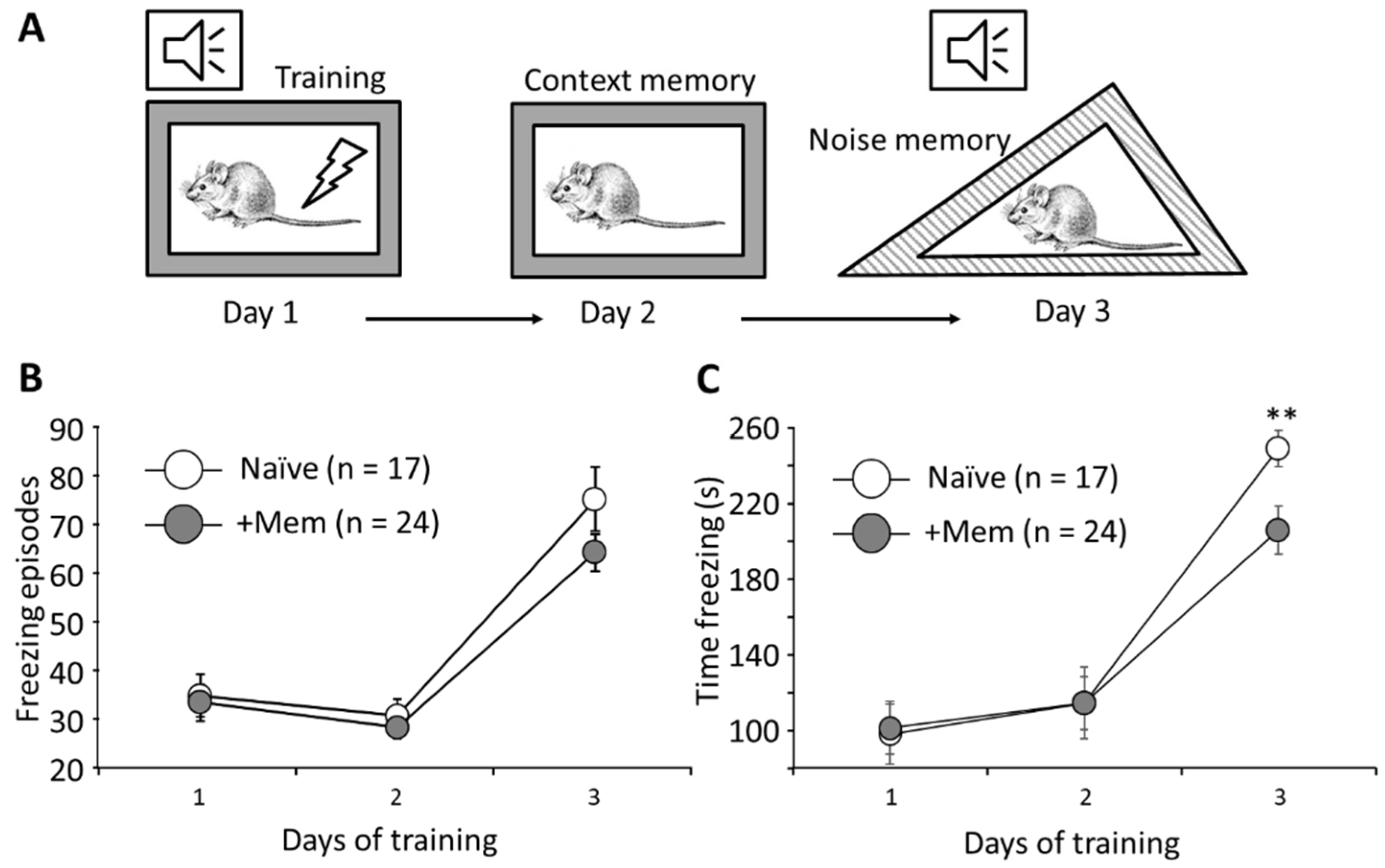
3.2. Long-Term Memantine Administration Affects Contextual Memory Retrieval
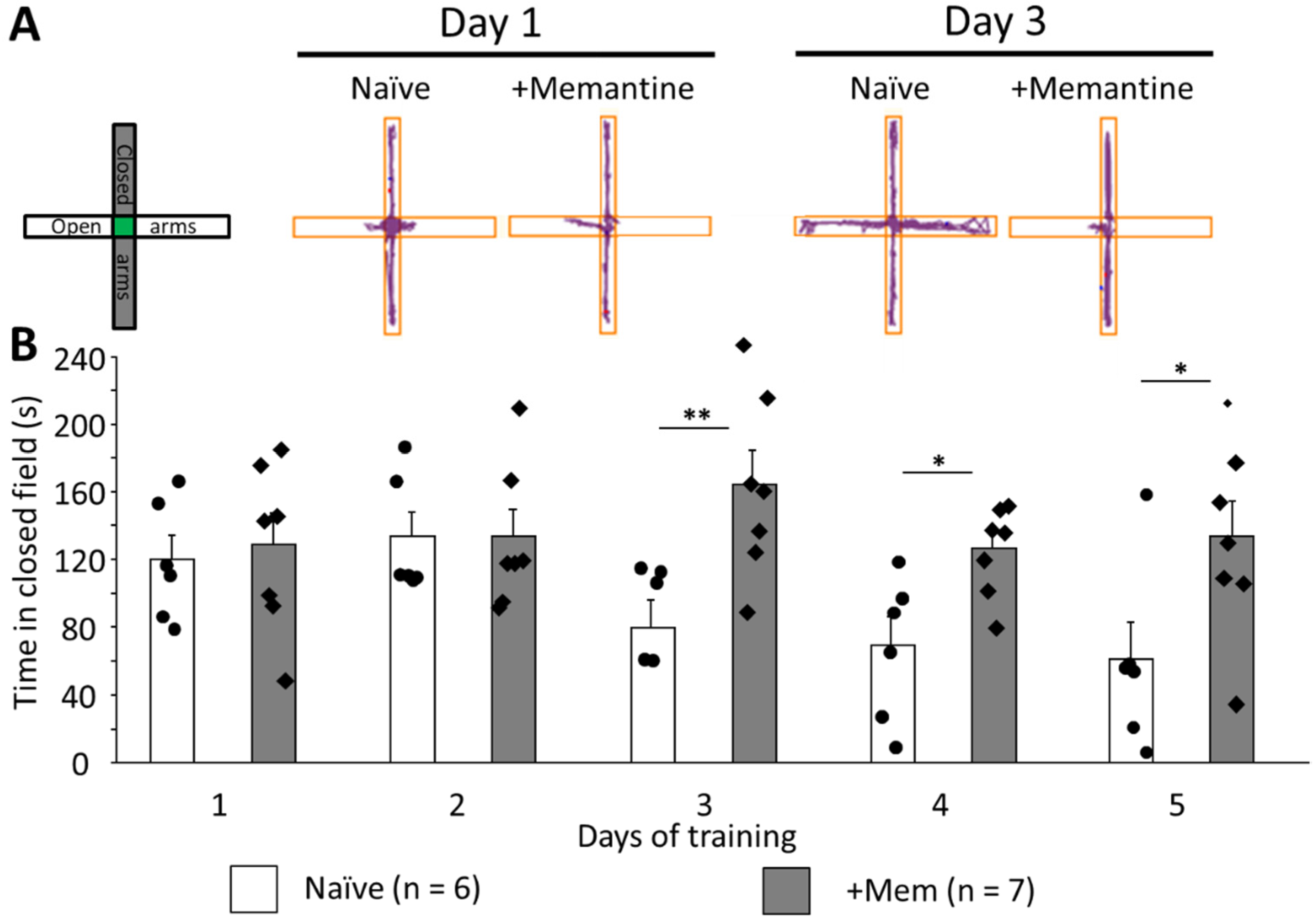
3.3. Long-Term Memantine Administration Prevents Adaptation to Anxiogenic Environment
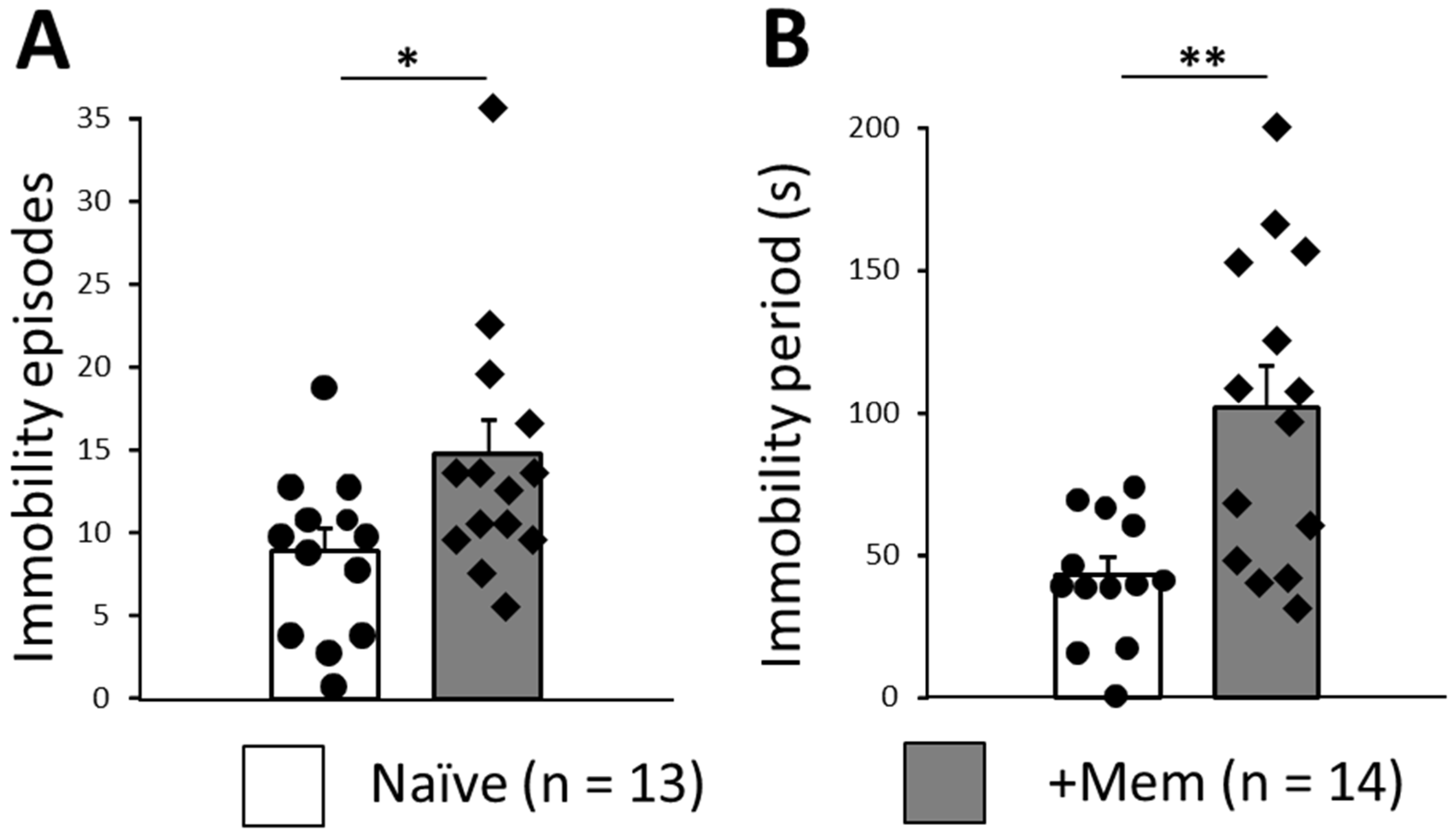
3.4. Long-Term Memantine Administration Induces Signs of Behavioral Despair in TST
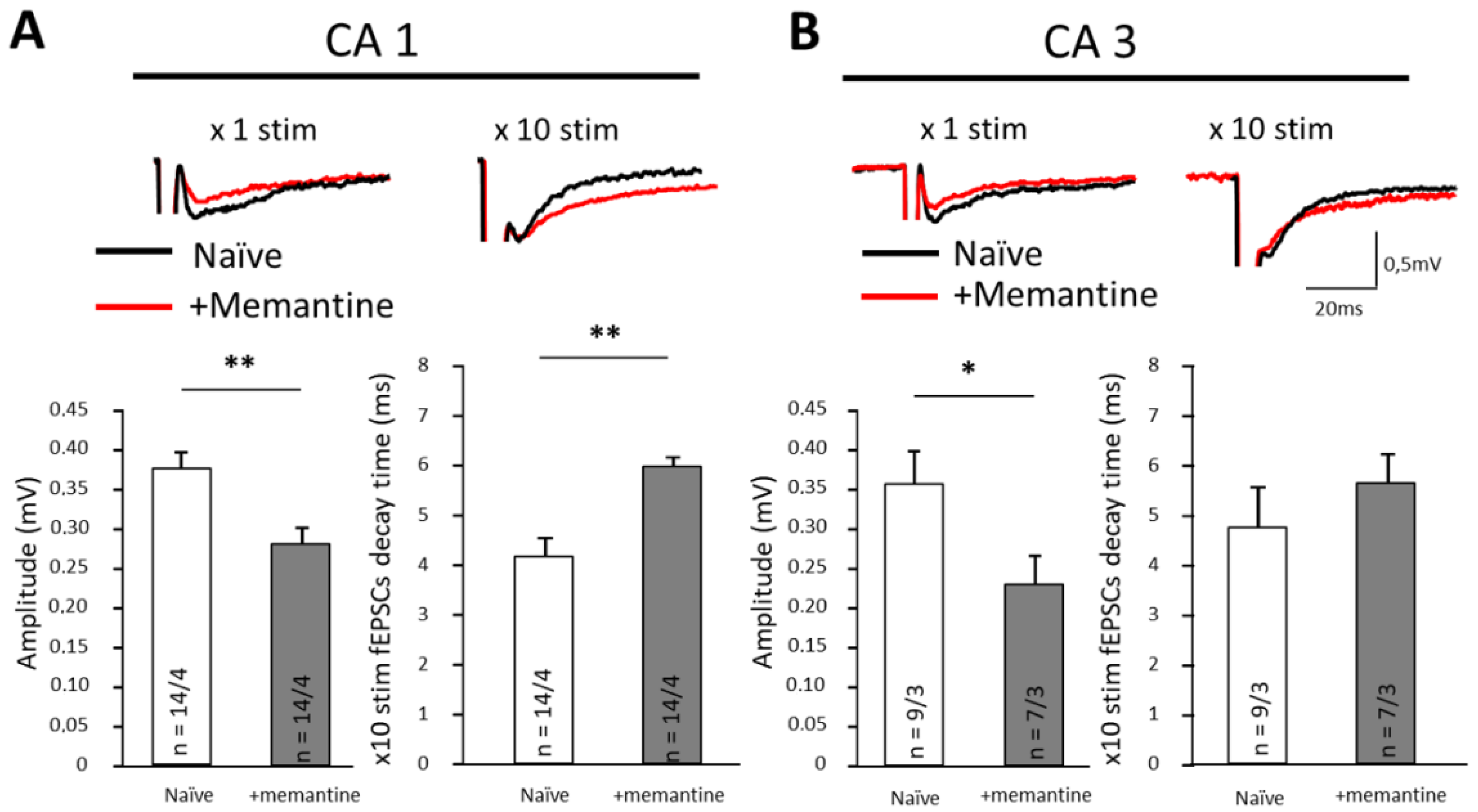
3.5. Memantine Disrupts Synaptic Transmission in Ventral Hippocampus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berliocchi, L.; Bano, D.; Nicotera, P. Ca2+ signals and death programmes in neurons. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 2255–2258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothstein, J.D. Excitatory hypothesis. Neurology 1996, 47 (Suppl. S2), S19–S25. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.M.; Keating, G.M. Memantine: A review of its use in Alzheimer’s disease. Drugs 2006, 66, 1515–1534. [Google Scholar] [CrossRef]
- Rogawski, M.A.; Wenk, G.L. The neuropharmacological basis for the use of memantine in the treatment of Alzheimer’s disease. CNS Drug Rev. 2003, 9, 275–308. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, S.; Kishi, T.; Iwata, N. Memantine monotherapy for Alzheimer’s disease: A systematic review and meta-analysis. PLoS ONE 2015, 10, e0123289. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Zhou, X.; Zhang, Q. Effectiveness and safety of memantine treatment for Alzheimer’s disease. J. Alzheimer’s Dis. 2013, 36, 445–458. [Google Scholar] [CrossRef]
- Zimmer, E.R.; Torrez, V.R.; Kalinine, E.; Augustin, M.C.; Zenki, K.C.; Almeida, R.F.; Hansel, G.; Muller, A.P.; Souza, D.O.; Machado-Vieira, R.; et al. Long-term NMDAR antagonism correlates reduced astrocytic glutamate uptake with anxiety-like phenotype. Front. Cell. Neurosci. 2015, 3, 219. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, S.; Kishi, T.; Nomura, I.; Sakuma, K.; Okuya, M.; Ikuta, T.; Iwata, N. The efficacy and safety of memantine for the treatment of Alzheimer’s disease. Expert Opin. Drug Saf. 2018, 17, 1053–1061. [Google Scholar] [CrossRef]
- Reisberg, B.; Doody, R.; Stöffler, A.; Schmitt, F.; Ferris, S.; Möbius, H.J. Memantine Study Group. Memantine in moderate-to-severe Alzheimer’s disease. N. Engl. J. Med. 2003, 348, 1333–1341. [Google Scholar] [CrossRef]
- Schwartz, T.L.; Siddiqui, U.A.; Raza, S. Memantine as an augmentation therapy for anxiety disorders. Case Rep. Psychiatry 2012, 2012, 749796. [Google Scholar] [CrossRef] [Green Version]
- Shuvaev, A.N.; Belozor, O.S.; Mozhei, O.; Yakovleva, D.A.; Potapenko, I.V.; Shuvaev, A.N.; Smolnikova, M.V.; Salmin, V.V.; Salmina, A.B.; Hirai, H.; et al. Chronic optogenetic stimulation of Bergman glia leads to dysfunction of EAAT1 and Purkinje cell death, mimicking the events caused by expression of pathogenic ataxin-1. Neurobiol. Dis. 2021, 154, 105340. [Google Scholar] [CrossRef] [PubMed]
- Shuvaev, A.N.; Belozor, O.S.; Mozhei, O.I.; Khilazheva, E.D.; Shuvaev, A.N.; Fritsler, Y.V.; Kasparov, S. Protective Effect of Memantine on Bergmann Glia and Purkinje Cells Morphology in Optogenetic Model of Neurodegeneration in Mice. Int. J. Mol. Sci. 2021, 22, 7822. [Google Scholar] [CrossRef]
- Bachmanov, A.A.; Reed, D.R.; Beauchamp, G.K.; Tordoff, M.G. Food intake, water intake, and drinking spout side preference of 28 mouse strains. Behav. Genet. 2002, 32, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Joo, I.S.; Hwang, D.H.; Seok, J.I.; Shin, S.K.; Kim, S.U. Oral administration of memantine prolongs survival in a transgenic mouse model of amyotrophic lateral sclerosis. J. Clin. Neurol. 2007, 3, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Shoji, H.; Takao, K.; Hattori, S.; Miyakawa, T. Contextual and Cued Fear Conditioning Test Using a Video Analyzing System in Mice. J. Vis. Exp. 2014, 85, e50871. [Google Scholar] [CrossRef] [Green Version]
- Morimura, N.; Yasuda, H.; Yamaguchi, K.; Katayama, K.; Hatayama, M.; Tomioka, N.H.; Odagawa, M.; Kamiya, A.; Iwayama, Y.; Maekawa, M.; et al. Autism-like Behaviours and Enhanced Memory Formation and Synaptic Plasticity in Lrfn2/SALM1-Deficient Mice. Nat. Commun. 2017, 8, 15800. [Google Scholar] [CrossRef] [Green Version]
- Walf, A.; Frye, C. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Asher, M.; Rosa, J.G.; Cvetanovic, M. Mood alterations in mouse models of Spinocerebellar Ataxia type 1. Sci Rep. 2021, 11, 713. [Google Scholar] [CrossRef]
- Krishnan, V.; Nestler, E.J. Animal models of depression: Molecular perspectives. Curr. Top Behav. Neurosci. 2011, 7, 121–147. [Google Scholar] [CrossRef] [Green Version]
- Cryan, J.F.; Mombereau, C.; Vassout, A. The tail suspension test as a model for assessing antidepressant activity: Review of pharmacological and genetic studies in mice. Neurosci. Biobehav. Rev. 2005, 29, 571–625. [Google Scholar] [CrossRef]
- Mathis, D.M.; Furman, J.L.; Norris, C.M. Preparation of Acute Hippocampal Slices from Rats and Transgenic Mice for the Study of Synaptic Alterations during Aging and Amyloid Pathology. J. Vis. Exp. 2011, 49, e2330. [Google Scholar] [CrossRef] [PubMed]
- Komleva, Y.K.; Lopatina, O.L.; Gorina, I.V.; Shuvaev, A.N.; Chernykh, A.; Potapenko, I.V.; Salmina, A.B. NLRP3 deficiency-induced hippocampal dysfunction and anxiety-like behavior in mice. Brain Res. 2021, 1752, 147220. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.; Holmes, A. The ascent of mouse: Advances in modelling human depression and anxiety. Nat. Rev. Drug Discov. 2005, 4, 775–790. [Google Scholar] [CrossRef]
- Hanson, J.E.; Meilandt, W.J.; Gogineni, A.; Reynen, P.; Herrington, J.; Weimer, R.M. Chronic GluN2B antagonism disrupts behavior in wild-type mice without protecting against synapse loss or memory impairment in Alzheimer’s disease mouse models. J. Neurosci. 2014, 34, 8277–8288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHugh, S.B.; Deacon, R.M.; Rawlins, J.N.; Bannerman, D.M. Amygdala and ventral hippocampus contribute differentially to mechanisms of fear and anxiety. Behav. Neurosci. 2004, 118, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, Y.; Shao, S.; Cui, S.; Wan, Y.; Yi, M. Ventral Hippocampus Modulates Anxiety-Like Behavior in Male But Not Female C57BL/6J Mice. Neuroscience 2019, 418, 50–58. [Google Scholar] [CrossRef]
- Duan, K.; Gu, Q.; Petralia, R.S.; Wang, Y.X.; Panja, D.; Liu, X.; Lehmann, M.L.; Zhu, H.; Zhu, J.; Li, Z. Mitophagy in the basolateral amygdala mediates increased anxiety induced by aversive social experience. Neuron 2021, 109, 3793–3809.e8. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kim, S.H.; Noh, Y.H.; Choi, B.M.; Noh, G.J.; Park, W.D.; Kim, E.J.; Cho, I.H.; Bae, C.S. Pharmacokinetics of Memantine after a Single and Multiple Dose of Oral and Patch Administration in Rats. Basic Clin. Pharmacol. Toxicol. 2016, 118, 122–127, Erratum in Basic Clin. Pharmacol. Toxicol. 2016, 118, 320. [Google Scholar] [CrossRef] [Green Version]
- Saab, B.J.; Roder, J.C. Acute pharmacokinetics of memantine in the mouse. Pharmacology 2011, 88, 284–287. [Google Scholar] [CrossRef]
- Spanagel, R.; Eilbacher, B.; Wilke, R. Memantine-induced dopamine release in the prefrontal cortex and striatum of the rat—A pharmacokinetic microdialysis study. Eur. J. Pharmacol. 1994, 262, 21–26. [Google Scholar] [CrossRef]
- Kotermanski, S.E.; Wood, J.T.; Johnson, J.W. Memantine binding to a superficial site on NMDA receptors contributes to partial trapping. J. Physiol. 2009, 587, 4589–4604. [Google Scholar] [CrossRef]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef]
- Kos, T.; Popik, P.A. Comparison of the predictive therapeutic and undesired side-effects of the NMDA receptor antagonist, memantine, in mice. Behav. Pharmacol. 2005, 16, 155–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CA1 | Amplitude (mV) | Rise Time (ms) | Decay Time (×1 stim.) | Decay Time (×10 stim.) |
|---|---|---|---|---|
| Control (n = 14/4) | 0.38 ± 0.02 | 2.8 ± 0.4 | 7.9 ± 0.5 | 4.2 ± 0.3 |
| +Memantine (n = 14/4) | 0.29 ± 0.02 ** | 2.2 ± 0.2 | 7.8 ± 0.7 | 5.9 ± 0.5 ** |
| CA3 | Amplitude (mV) | Rise Time (ms) | Decay Time (×1 stim.) | Decay Time (×10 stim.) |
|---|---|---|---|---|
| Control (n = 9/3) | 0.36 ± 0.04 | 2.1 ± 0.1 | 5.2 ± 0.6 | 4.9 ± 0.9 |
| +Memantine (n = 7/3) | 0.22 ± 0.03 * | 1.7 ± 0.2 | 4.9 ± 1.1 | 5.4 ± 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shuvaev, A.N.; Belozor, O.S.; Mozhei, O.I.; Mileiko, A.G.; Mosina, L.D.; Laletina, I.V.; Mikhailov, I.G.; Fritsler, Y.V.; Shuvaev, A.N.; Teschemacher, A.G.; et al. Memantine Disrupts Motor Coordination through Anxiety-like Behavior in CD1 Mice. Brain Sci. 2022, 12, 495. https://doi.org/10.3390/brainsci12040495
Shuvaev AN, Belozor OS, Mozhei OI, Mileiko AG, Mosina LD, Laletina IV, Mikhailov IG, Fritsler YV, Shuvaev AN, Teschemacher AG, et al. Memantine Disrupts Motor Coordination through Anxiety-like Behavior in CD1 Mice. Brain Sciences. 2022; 12(4):495. https://doi.org/10.3390/brainsci12040495
Chicago/Turabian StyleShuvaev, Anton N., Olga S. Belozor, Oleg I. Mozhei, Aleksandra G. Mileiko, Ludmila D. Mosina, Irina V. Laletina, Ilia G. Mikhailov, Yana V. Fritsler, Andrey N. Shuvaev, Anja G. Teschemacher, and et al. 2022. "Memantine Disrupts Motor Coordination through Anxiety-like Behavior in CD1 Mice" Brain Sciences 12, no. 4: 495. https://doi.org/10.3390/brainsci12040495