
The Detection of Face-like Stimuli at the Edge of the Infant Visual Field


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
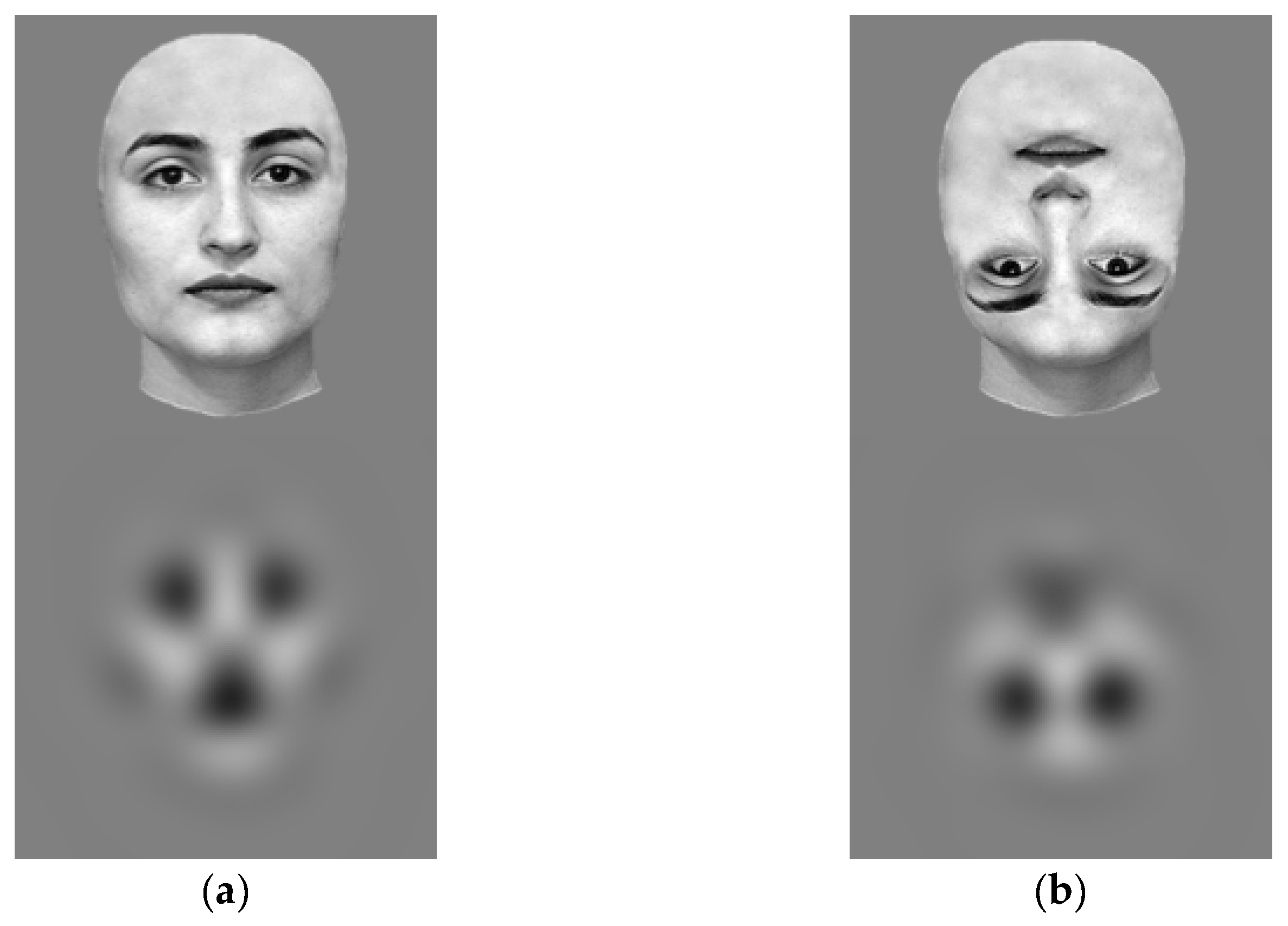
2.2. Stimuli
2.3. Apparatus and Procedure
2.4. Data Processing and Coding
2.5. Analysis and Statistics
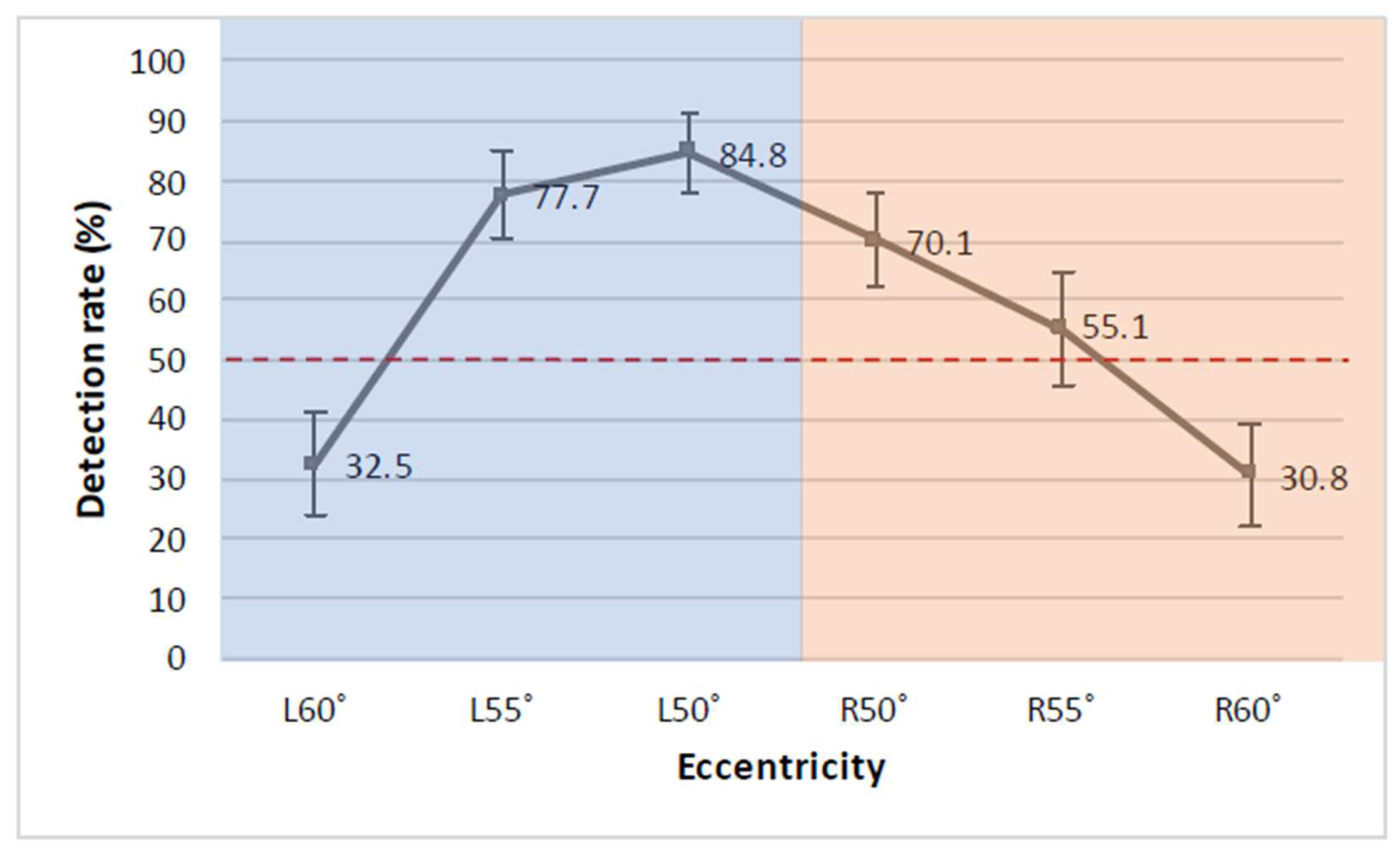
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, M.; Morton, J. Biology and Cognitive Development: The Case of Face Recognition; Blackwell: Oxford, UK, 1991. [Google Scholar]
- Valenza, E.; Simion, F.; Cassia, V.M.; Umiltà, C. Face preference at birth. J. Exp. Psychol. Hum. 1996, 22, 892. [Google Scholar] [CrossRef]
- Reid, V.M.; Dunn, K.; Young, R.J.; Amu, J.; Donovan, T.; Reissland, N. The human fetus preferentially engages with face-like visual stimuli. Curr. Biol. 2017, 27, 1825–1828.e1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turati, C.; Valenza, E.; Leo, I.; Simion, F. Three-month-olds’ visual preference for faces and its underlying visual processing mechanisms. J. Exp. Child Psychol. 2005, 90, 255–273. [Google Scholar] [CrossRef] [PubMed]
- Chien, S.H.-L. No more top-heavy bias: Infants and adults prefer upright faces but not top-heavy geometric or face-like patterns. J. Vis. 2011, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cashon, C.H.; Cohen, L.B. Beyond U-shaped development in infants’ processing of faces: An information-processing account. J. Cogn. Dev. 2004, 5, 59–80. [Google Scholar] [CrossRef]
- Cohen, L.B.; Cashon, C.H. Do 7-month-old infants process independent features or facial configurations? Infant Child Dev. 2001, 10, 83–92. [Google Scholar] [CrossRef]
- Pascalis, O.; De Haan, M.; Nelson, C.A. Is face processing species-specific during the first year of life? Sci. J. 2002, 296, 1321–1323. [Google Scholar] [CrossRef] [Green Version]
- Quinn, P.C.; Yahr, J.; Kuhn, A.; Slater, A.M.; Pascalis, O. Representation of the gender of human faces by infants: A preference for female. Perception 2002, 31, 1109–1121. [Google Scholar] [CrossRef] [Green Version]
- Morton, J.; Johnson, M.H. CONSPEC and CONLERN: A two-process theory of infant face recognition. Psychol. Rev. 1991, 98, 164. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.H.; Senju, A.; Tomalski, P. The two-process theory of face processing: Modifications based on two decades of data from infants and adults. Neurosci. Biobehav. Rev. 2015, 50, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Simion, F.; Macchi Cassia, V.; Turati, C.; Valenza, E. The origins of face perception: Specific versus non-specific mechanisms. Infant Child Dev. 2001, 10, 59–65. [Google Scholar] [CrossRef]
- Slater, A.; Quinn, P.C.; Kelly, D.J.; Lee, K.; Longmore, C.A.; McDonald, P.R.; Pascalis, O. The shaping of the face space in early infancy: Becoming a native face processor. Child Dev. Perspect. 2010, 4, 205–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, N.; Paikan, A.; Gredebäck, G.; Rea, F.; Metta, G. Staring us in the face? An embodied theory of innate face preference. Dev. Sci. 2014, 17, 809–825. [Google Scholar] [CrossRef] [PubMed]
- Curcio, C.A.; Sloan, K.R.; Kalina, R.E.; Hendrickson, A.E. Human photoreceptor topography. J. Comp. Neurol. 1990, 292, 497–523. [Google Scholar] [CrossRef]
- Fletcher-Watson, S.; Findlay, J.M.; Leekam, S.R.; Benson, V. Rapid detection of person information in a naturalistic scene. Perception 2008, 37, 571–583. [Google Scholar] [CrossRef]
- Kelly, D.J.; Duarte, S.; Meary, D.; Bindemann, M.; Pascalis, O. Infants rapidly detect human faces in complex naturalistic visual scenes. Dev. Sci. 2019, 22, e12829. [Google Scholar] [CrossRef] [Green Version]
- Freedman, E.G. Coordination of the eyes and head during visual orienting. Exp. Brain Res. 2008, 190, 369–387. [Google Scholar] [CrossRef] [Green Version]
- Stahl, J.S. Amplitude of human head movements associated with horizontal saccades. Exp. Brain Res. 1999, 126, 41–54. [Google Scholar] [CrossRef]
- Goren, C.C.; Sarty, M.; Wu, P.Y. Visual following and pattern discrimination of face-like stimuli by newborn infants. Pediatrics 1975, 56, 544–549. [Google Scholar] [CrossRef]
- Johnson, M.H.; Dziurawiec, S.; Ellis, H.; Morton, J. Newborns’ preferential tracking of face-like stimuli and its subsequent decline. Cognition 1991, 40, 1–19. [Google Scholar] [CrossRef]
- Johnson, M.H. Subcortical face processing. Nat. Rev. Neurosci. 2005, 6, 766–774. [Google Scholar] [CrossRef]
- Dobson, V.; Brown, A.M.; Harvey, E.M.; Narter, D.B. Visual field extent in children 3.5–30 months of age tested with a double-arc LED perimeter. Vis. Res. 1998, 38, 2743–2760. [Google Scholar] [CrossRef] [Green Version]
- Harris, P.; MacFarlane, A. The growth of the effective visual field from birth to seven weeks. J. Exp. Child Psychol. 1974, 18, 340–348. [Google Scholar] [CrossRef]
- Maurer, D.; Lewis, T.L. The development of peripheral vision and its physiological underpinnings. In Newborn Attention: Biological Constraints and the Influence of Experience; Weiss, M.J.S., Zelazo, P.R., Eds.; Ablex Publishing: Norwood, NJ, USA, 1991; pp. 218–255. [Google Scholar]
- Mayer, D.L.; Fulton, A.B.; Cummings, M.F. Visual fields of infants assessed with a new perimetric technique. Investig. Ophth. Vis. Sci. 1988, 29, 452–459. [Google Scholar]
- Capparini, C.; To, M.P.S.; Reid, V.M. Identifying the limits of peripheral visual processing in 9-month-old infants. Dev. Psychobiol. 2022, in press. [Google Scholar] [CrossRef]
- Dundas, E.; Gastgeb, H.; Strauss, M.S. Left visual field biases when infants process faces: A comparison of infants at high- and low-risk for autism spectrum disorder. J. Autism Dev. Disord. 2012, 42, 2659–2668. [Google Scholar] [CrossRef]
- Yovel, G.; Tambini, A.; Brandman, T. The asymmetry of the fusiform face area is a stable individual characteristic that underlies the left-visual-field superiority for faces. Neuropsychologia 2008, 46, 3061–3068. [Google Scholar] [CrossRef]
- Mäkelä, P.; Näsänen, R.; Rovamo, J.; Melmoth, D. Identification of facial images in peripheral vision. Vis. Res. 2001, 41, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Martelli, M.; Majaj, N.J.; Pelli, D.G. Are faces processed like words? A diagnostic test for recognition by parts. J. Vis. 2005, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Kanwisher, N. Faces and places: Of central (and peripheral) interest. Nat. Neurosci. 2001, 4, 455–456. [Google Scholar] [CrossRef]
- Levy, I.; Hasson, U.; Avidan, G.; Hendler, T.; Malach, R. Center–periphery organization of human object areas. Nat. Neurosci. 2001, 4, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Rousselet, G.A.; Husk, J.S.; Bennett, P.J.; Sekuler, A.B. Spatial scaling factors explain eccentricity effects on face ERPs. J. Vis. 2005, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Jebara, N.; Pins, D.; Despretz, P.; Boucart, M. Face or building superiority in peripheral vision reversed by task requirements. Adv. Cogn. Psychol. 2009, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Hershler, O.; Golan, T.; Bentin, S.; Hochstein, S. The wide window of face detection. J. Vis. 2010, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Boucart, M.; Lenoble, Q.; Quettelart, J.; Szaffarczyk, S.; Despretz, P.; Thorpe, S.J. Finding faces, animals, and vehicles in far peripheral vision. J. Vis. 2016, 16, 10. [Google Scholar] [CrossRef]
- Macchi Cassia, V.; Turati, C.; Simion, F. Can a nonspecific bias toward top-heavy patterns explain newborns’ face preference? Psychol. Sci. 2004, 15, 379–383. [Google Scholar] [CrossRef]
- To, M.; Regan, B.; Wood, D.; Mollon, J. Vision out of the corner of the eye. Vis. Res. 2011, 51, 203–214. [Google Scholar] [CrossRef]
- Brainard, D.H.; Vision, S. The psychophysics toolbox. Spat. Vis. 1997, 10, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Kleiner, M.; Brainard, D.; Pelli, D.; Ingling, A.; Murray, R.; Broussard, C. What’s new in Psychtoolbox-3? Perception 2007, 36, 1–16. [Google Scholar]
- ELAN, Version 5.9 [Computer Software] Nijmegen: Max Planck Institute for Psycholinguistics, The Language Archive. 2020. Available online: https://archive.mpi.nl/tla/elan (accessed on 27 February 2022).
- Brugman, H.; Russel, A. Annotating Multi-media/Multi-modal Resources with ELAN. In Proceedings of the LREC, Fourth International Conference on Language Resources and Evaluation, Lisbon, Portugal, 26–28 May 2004; pp. 2065–2068. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 27 February 2022).
- Quené, H.; Van den Bergh, H. On multi-level modeling of data from repeated measures designs: A tutorial. Speech Commun. 2004, 43, 103–121. [Google Scholar] [CrossRef]
- Lewis, T.L.; Maurer, D. The development of the temporal and nasal visual fields during infancy. Vis. Res. 1992, 32, 903–911. [Google Scholar] [CrossRef]
- Johnson, M.H. Face processing as a brain adaptation at multiple timescales. Q. J. Exp. Psychol. 2011, 64, 1873–1888. [Google Scholar] [CrossRef] [PubMed]
- Tomalski, P.; Csibra, G.; Johnson, M.H. Rapid orienting toward face-like stimuli with gaze-relevant contrast information. Perception 2009, 38, 569–578. [Google Scholar] [CrossRef]
- Tomalski, P.; Johnson, M.H.; Csibra, G. Temporal-nasal asymmetry of rapid orienting to face-like stimuli. Neuroreport 2009, 20, 1309–1312. [Google Scholar] [CrossRef] [PubMed]
- Gliga, T.; Elsabbagh, M.; Andravizou, A.; Johnson, M. Faces attract infants’ attention in complex displays. Infancy 2009, 14, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Valenza, E.; Otsuka, Y.; Bulf, H.; Ichikawa, H.; Kanazawa, S.; Yamaguchi, M.K. Face orientation and motion differently affect the deployment of visual attention in newborns and 4-month-old infants. PLoS ONE 2015, 10, e0136965. [Google Scholar] [CrossRef] [Green Version]
- Brown, V.; Huey, D.; Findlay, J.M. Face detection in peripheral vision: Do faces pop out? Perception 1997, 26, 1555–1570. [Google Scholar] [CrossRef]
- Calvo, M.G.; Nummenmaa, L.; Avero, P. Recognition advantage of happy faces in extrafoveal vision: Featural and affective processing. Vis. Cogn. 2010, 18, 1274–1297. [Google Scholar] [CrossRef]
- Guo, K.; Meints, K.; Hall, C.; Hall, S.; Mills, D. Left gaze bias in humans, rhesus monkeys and domestic dogs. Anim. Cogn. 2009, 12, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Carey, S.; Diamond, R. Maturational determination of the developmental course of face encoding. In Biological Studies of Mental Processes; Caplan, D., Ed.; The MIT Press: Cambridge, MA, USA, 1980; pp. 60–93. [Google Scholar]
- Kanwisher, N.; Yovel, G. The fusiform face area: A cortical region specialized for the perception of faces. Philos. Trans. R. Soc. B 2006, 361, 2109–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, S.C.; Banich, M.T.; Koch-Weser, M.P. Face recognition: A general or specific right hemisphere capacity? Brain Cogn. 1988, 8, 303–325. [Google Scholar] [CrossRef]
- Forrester, G.S.; Davis, R.; Mareschal, D.; Malatesta, G.; Todd, B.K. The left cradling bias: An evolutionary facilitator of social cognition? Cortex 2019, 118, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Giljov, A.; Karenina, K.; Malashichev, Y. Facing each other: Mammal mothers and infants prefer the position favouring right hemisphere processing. Biol. Lett. 2018, 14, 20170707. [Google Scholar] [CrossRef]
- Malatesta, G.; Marzoli, D.; Tommasi, L. Keep a left profile, baby! The left-cradling bias is associated with a preference for left-facing profiles of human babies. Symmetry 2020, 12, 911. [Google Scholar] [CrossRef]
- Davis, R.; Donati, G.; Finnegan, K.; Boardman, J.P.; Fletcher-Watson, S.; Forrester, G.S. Social gaze in preterm infants may act as an early indicator of atypical lateralization. Child Dev. 2022. Advance online publication. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fixed Effects | |||||
|---|---|---|---|---|---|
| Estimate | SE | 95% CI | Z | p | |
| Intercept | 1.58 | 0.37 | 0.85, 2.31 | 4.24 | <0.001 *** |
| Eccentricity (55°) | −0.59 | 0.39 | −1.35, 0.18 | −1.50 | 0.13 |
| Eccentricity (60°) | −2.01 | 0.41 | −2.81, −1.22 | −4.95 | <0.001 *** |
| Side (Right) | −0.66 | 0.32 | −1.29, −0.03 | −2.04 | 0.04 * |
| Random Effects | |||||
| Variance | SD | 95% CI | |||
| Intercept | 0.20 | 0.45 | 0, 1.04 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capparini, C.; To, M.P.S.; Reid, V.M. The Detection of Face-like Stimuli at the Edge of the Infant Visual Field. Brain Sci. 2022, 12, 493. https://doi.org/10.3390/brainsci12040493
Capparini C, To MPS, Reid VM. The Detection of Face-like Stimuli at the Edge of the Infant Visual Field. Brain Sciences. 2022; 12(4):493. https://doi.org/10.3390/brainsci12040493
Chicago/Turabian StyleCapparini, Chiara, Michelle P. S. To, and Vincent M. Reid. 2022. "The Detection of Face-like Stimuli at the Edge of the Infant Visual Field" Brain Sciences 12, no. 4: 493. https://doi.org/10.3390/brainsci12040493