
It Takes Two: Interpersonal Neural Synchrony Is Increased after Musical Interaction


Abstract
:1. Introduction
2. Materials and Methods
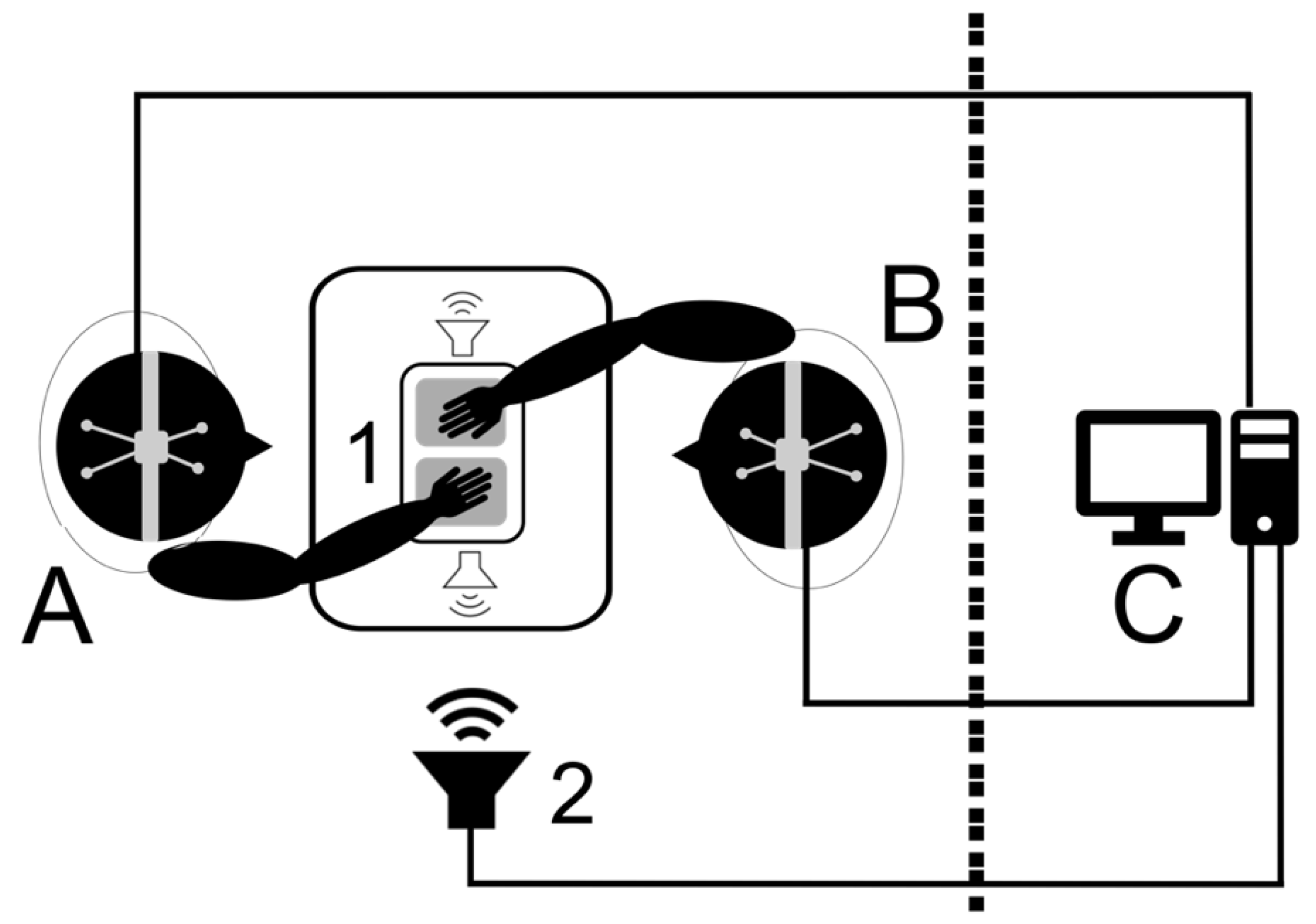
2.1. Participants and Setup
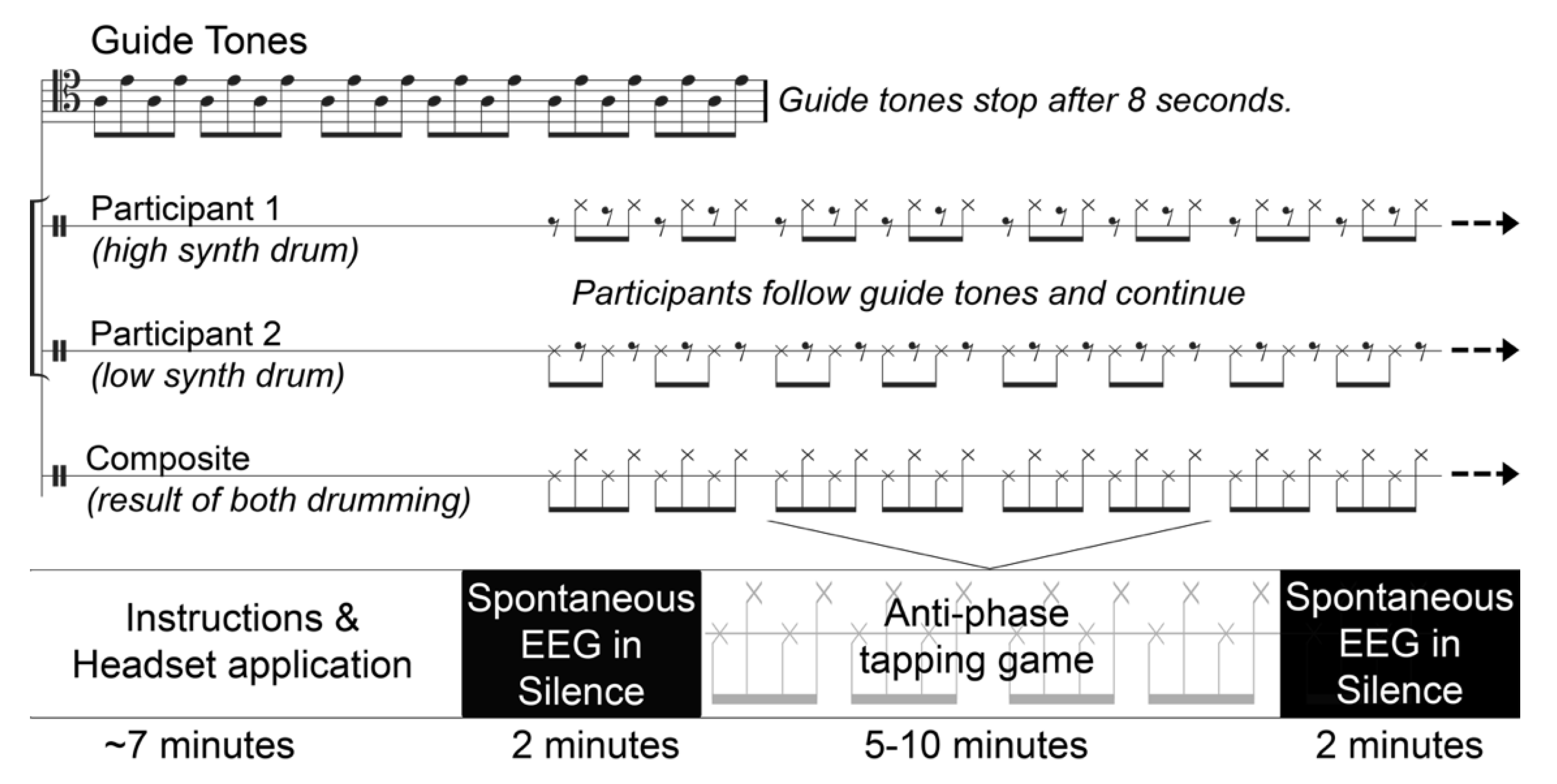
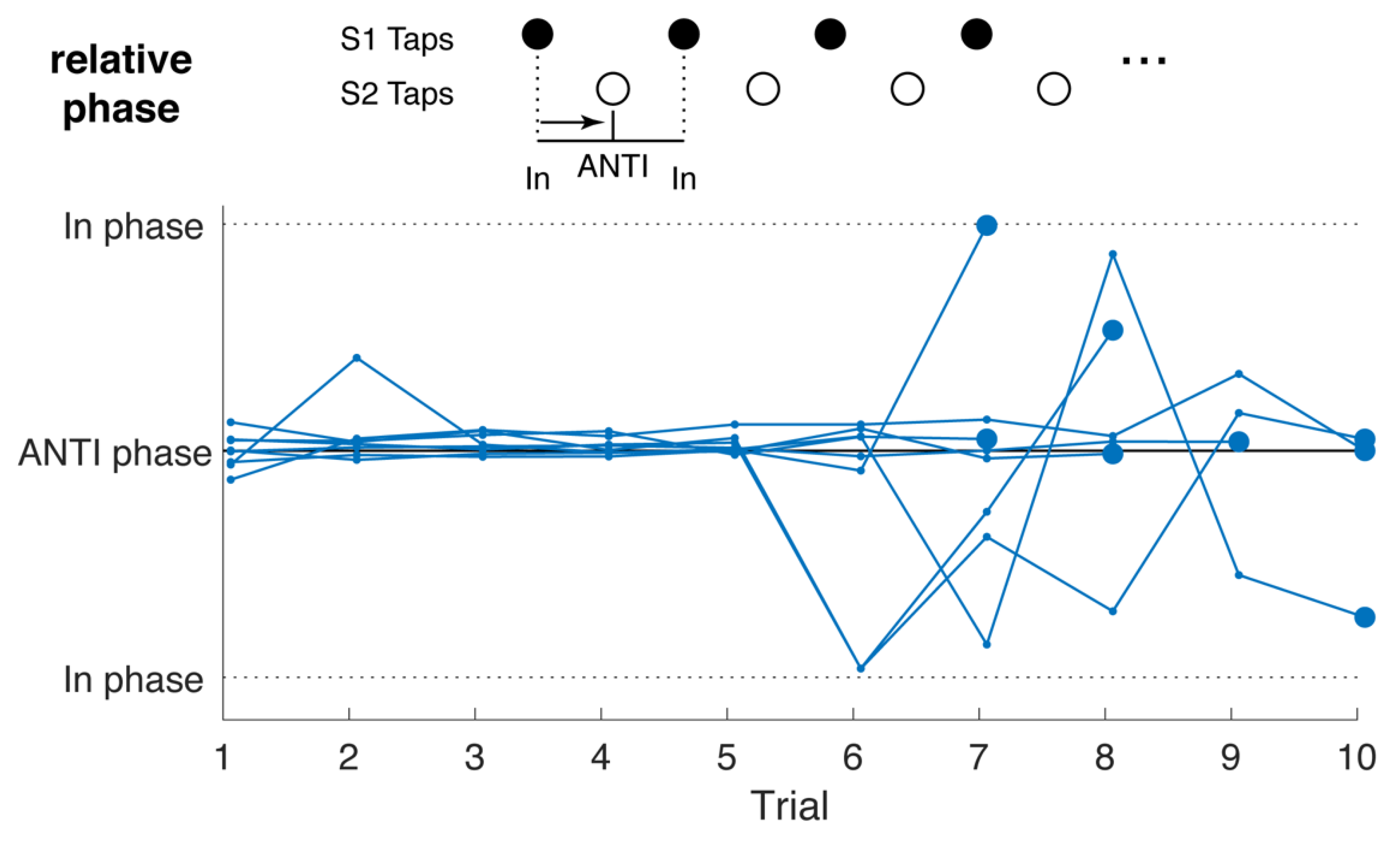
2.2. Task
2.3. Procedure
3. Data Analysis
3.1. EEG Processing
3.2. Statistical Analysis
4. Results
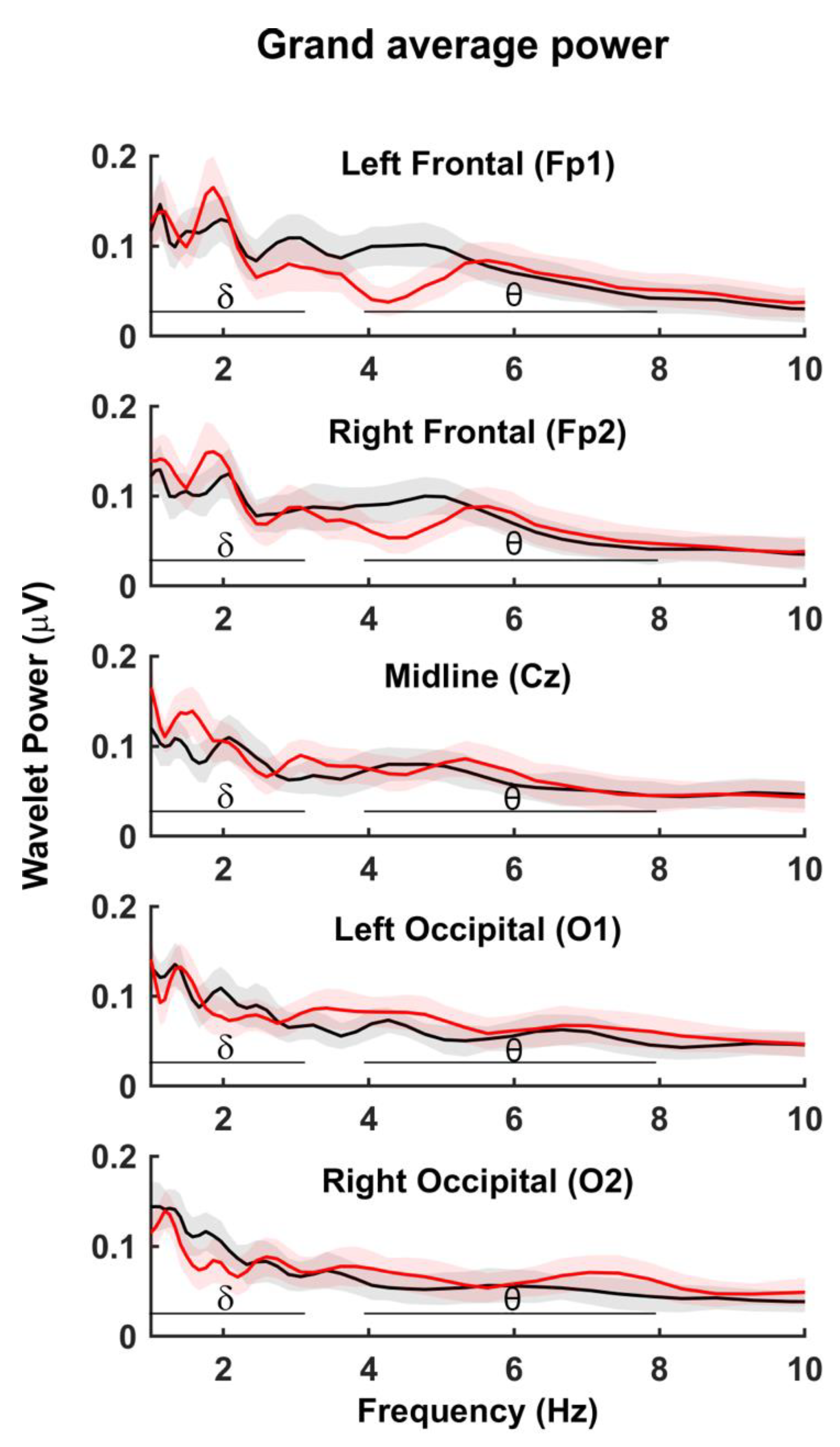
4.1. Wavelet Power
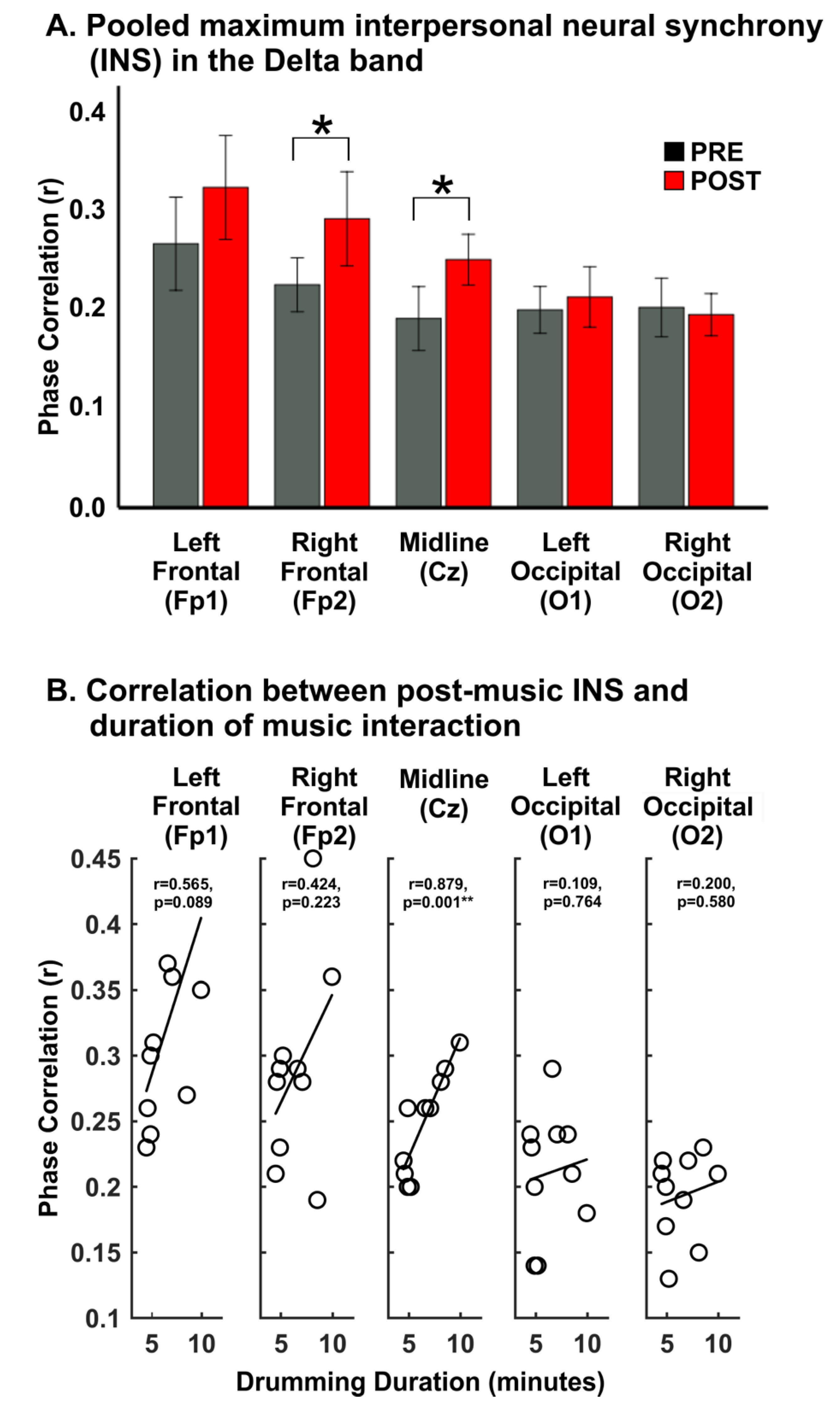
4.2. Interpersonal Neural Synchrony—Frequency Specific Phase Coupling
4.3. Relationships between Interpersonal Neural Synchrony and Duration of Musical Interaction
4.4. Tapping Characterization
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Channels | Delta Power (SE) | Theta Power (SE) |
|---|---|---|---|
| PRE (before rhythm activity) | Fp1 | 14.3 (1.9) | 7.1 (1.0) |
| Fp2 | 13.3 (1.9) | 6.9 (1.0) | |
| Cz | 5.7 (0.7) | 4.5 (0.5) | |
| O1 | 7.9 (2.2) | 4.9 (0.8) | |
| O2 | 10.4 (3.4) | 5.3 (0.8) | |
| POST (after rhythm activity) | Fp1 | 17.6 (2.5) | 7.0 (0.9) |
| Fp2 | 17.4 (2.4) | 6.9 (0.9) | |
| Cz | 6.9 (0.9) | 4.5 (0.5) | |
| O1 | 8.5 (3.5) | 4.6 (1.1) | |
| O2 | 9.0 (2.4) | 4.6 (0.8) |
Appendix B
| Maximum Inter-Brain Correlation, Delta Band | |||||||
|---|---|---|---|---|---|---|---|
| Condition Dyad | R Deltamax Fp1 | R Deltamax Fp2 | R Deltamax Cz | R Deltamax O1 | R Deltamax O2 | ||
| PRE | 1 | 0.19 | 0.17 | 0.19 | 0.13 | 0.15 | |
| 2 | 0.24 | 0.25 | 0.16 | 0.23 | 0.18 | ||
| 3 | 0.22 | 0.21 | 0.15 | 0.20 | 0.27 | ||
| 4 | 0.20 | 0.18 | 0.27 | 0.25 | 0.16 | ||
| 5 | 0.44 | 0.27 | 0.10 | 0.16 | 0.20 | ||
| 6 | 0.23 | 0.15 | 0.17 | 0.18 | 0.17 | ||
| 7 | 0.30 | 0.28 | 0.19 | 0.17 | 0.23 | ||
| 8 | 0.28 | 0.24 | 0.26 | 0.23 | 0.26 | ||
| 9 | 0.22 | 0.23 | 0.20 | 0.19 | 0.23 | ||
| 10 | 0.30 | 0.24 | 0.21 | 0.23 | 0.13 | ||
| Mean | 0.2634 | 0.2225 | 0.1888 | 0.1974 | 0.1998 | ||
| Std. Error of Mean | 0.02327 | 0.01348 | 0.01596 | 0.01169 | 0.01460 | ||
| POST | 1 | 0.37 | 0.29 | 0.26 | 0.29 | 0.19 | |
| 2 | 0.51 | 0.45 | 0.28 | 0.24 | 0.15 | ||
| 3 | 0.26 | 0.28 | 0.21 | 0.23 | 0.22 | ||
| 4 | 0.31 | 0.30 | 0.17 | 0.14 | 0.13 | ||
| 5 | 0.23 | 0.21 | 0.22 | 0.24 | 0.21 | ||
| 6 | 0.36 | 0.28 | 0.26 | 0.24 | 0.22 | ||
| 7 | 0.27 | 0.19 | 0.29 | 0.21 | 0.23 | ||
| 8 | 0.35 | 0.36 | 0.31 | 0.18 | 0.21 | ||
| 9 | 0.30 | 0.29 | 0.26 | 0.20 | 0.17 | ||
| 10 | 0.24 | 0.23 | 0.20 | 0.14 | 0.20 | ||
| Mean | 0.3197 | 0.2883 | 0.2446 | 0.2103 | 0.1927 | ||
| Std. Error of Mean | 0.02595 | 0.02352 | 0.01426 | 0.01510 | 0.01050 | ||
| Maximum Inter-Brain Correlation, Theta Band | |||||||
|---|---|---|---|---|---|---|---|
| Condition Dyad | R Thetamax Fp1 | R Thetamax Fp2 | R Thetamax Cz | R Thetamax O1 | R Thetamax O2 | ||
| PRE | 1 | 0.16 | 0.14 | 0.15 | 0.11 | 0.08 | |
| 2 | 0.16 | 0.13 | 0.11 | 0.08 | 0.08 | ||
| 3 | 0.19 | 0.17 | 0.14 | 0.09 | 0.08 | ||
| 4 | 0.20 | 0.20 | 0.16 | 0.12 | 0.12 | ||
| 5 | 0.14 | 0.10 | 0.11 | 0.09 | 0.05 | ||
| 6 | 0.10 | 0.12 | 0.12 | 0.06 | 0.07 | ||
| 7 | 0.14 | 0.08 | 0.13 | 0.10 | 0.08 | ||
| 8 | 0.10 | 0.11 | 0.09 | 0.15 | 0.15 | ||
| 9 | 0.15 | 0.17 | 0.14 | 0.10 | 0.10 | ||
| 10 | 0.13 | 0.12 | 0.09 | 0.08 | 0.14 | ||
| Mean | 0.1467 | 0.1349 | 0.1256 | 0.0985 | 0.0955 | ||
| Std. Error of Mean | 0.01044 | 0.01150 | 0.00759 | 0.00775 | 0.01020 | ||
| POST | 1 | 0.16 | 0.13 | 0.13 | 0.10 | 0.07 | |
| 2 | 0.15 | 0.19 | 0.12 | 0.07 | 0.09 | ||
| 3 | 0.15 | 0.12 | 0.16 | 0.18 | 0.10 | ||
| 4 | 0.06 | 0.08 | 0.09 | 0.11 | 0.12 | ||
| 5 | 0.14 | 0.16 | 0.19 | 0.12 | 0.14 | ||
| 6 | 0.10 | 0.09 | 0.08 | 0.09 | 0.18 | ||
| 7 | 0.13 | 0.08 | 0.10 | 0.12 | 0.10 | ||
| 8 | 0.06 | 0.07 | 0.13 | 0.15 | 0.08 | ||
| 9 | 0.17 | 0.18 | 0.14 | 0.15 | 0.17 | ||
| 10 | 0.13 | 0.11 | 0.11 | 0.09 | 0.13 | ||
| Mean | 0.1256 | 0.1208 | 0.1245 | 0.1187 | 0.1171 | ||
| Std. Error of Mean | 0.01205 | 0.01362 | 0.01038 | 0.01040 | 0.01211 | ||
| Dyad | Duration (Seconds) |
|---|---|
| 1 | 594 |
| 2 | 534 |
| 3 | 486 |
| 4 | 378 |
| 5 | 366 |
| 6 | 318 |
| 7 | 312 |
| 8 | 300 |
| 9 | 294 |
| 10 | 276 |
References
- Delaherche, E.; Boucenna, S.; Karp, K.; Michelet, S.; Achard, C.; Chetouani, M. Social Coordination Assessment: Distinguishing between Shape and Timing. In Proceedings of the IAPR Workshop on Multimodal Pattern Recognition of Social Signals in Human-Computer Interaction; Springer: Berlin/Heidelberg, Germany, 2012; pp. 9–18. [Google Scholar]
- Kinsbourne, M.; Helt, M. Entrainment, Mimicry, and Interpersonal Synchrony. In The Neuropsychology of Autism; Oxford University Press: Oxford, UK, 2011; pp. 339–365. [Google Scholar]
- Chetouani, M.; Delaherche, E.; Dumas, G.; Cohen, D. 15 Interpersonal Synchrony: From Social Perception to Social Interaction. In Social Signal Processing; Cambridge University Press: Cambridge, UK, 2017; p. 202. [Google Scholar]
- Mogan, R.; Fischer, R.; Bulbulia, J.A. To Be in Synchrony or Not? A Meta-Analysis of Synchrony’s Effects on Behavior, Perception, Cognition and Affect. J. Exp. Soc. Psychol. 2017, 72, 13–20. [Google Scholar] [CrossRef]
- Beňuš, Š. Social Aspects of Entrainment in Spoken Interaction. Cogn. Comput. 2014, 6, 802–813. [Google Scholar] [CrossRef]
- Lumsden, J.; Miles, L.K.; Macrae, C.N. Sync or Sink? Interpersonal Synchrony Impacts Self-Esteem. Front. Psychol. 2014, 5, 1064. [Google Scholar] [CrossRef]
- Khalil, A.K.; Minces, V.; McLoughlin, G.; Chiba, A. Group Rhythmic Synchrony and Attention in Children. Front. Psychol. 2013, 4, 564. [Google Scholar] [CrossRef] [Green Version]
- Cirelli, L.K.; Einarson, K.M.; Trainor, L.J. Interpersonal Synchrony Increases Prosocial Behavior in Infants. Dev. Sci. 2014, 17, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Rennung, M.; Göritz, A.S. Prosocial Consequences of Interpersonal Synchrony. Z. Psychol. 2016, 224, 168–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tunçgenç, B.; Cohen, E. Interpersonal Movement Synchrony Facilitates Pro-Social Behavior in Children’s Peer-Play. Dev. Sci. 2018, 21, e12505. [Google Scholar] [CrossRef] [PubMed]
- Cross, L.; Turgeon, M.; Atherton, G. How Moving Together Binds Us Together: The Social Consequences of Interpersonal Entrainment and Group Processes. Open Psychol. 2019, 1, 273–302. [Google Scholar] [CrossRef]
- Fessler, D.M.; Holbrook, C. Synchronized Behavior Increases Assessments of the Formidability and Cohesion of Coalitions. Evol. Hum. Behav. 2016, 37, 502–509. [Google Scholar] [CrossRef] [Green Version]
- Hove, M.J.; Risen, J.L. It’s All in the Timing: Interpersonal Synchrony Increases Affiliation. Soc. Cogn. 2009, 27, 949–960. [Google Scholar] [CrossRef] [Green Version]
- Paxton, A.; Dale, R. Argument Disrupts Interpersonal Synchrony; SAGE Publications Sage UK: London, UK, 2013. [Google Scholar]
- Kupper, Z.; Ramseyer, F.; Hoffmann, H.; Tschacher, W. Nonverbal Synchrony in Social Interactions of Patients with Schizophrenia Indicates Socio-Communicative Deficits. PLoS ONE 2015, 10, e0145882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, M.; Srinivasan, S.M.; Bhat, A.N. Comparing Motor Performance, Praxis, Coordination, and Interpersonal Synchrony between Children with and without Autism Spectrum Disorder (ASD). Res. Dev. Disabil. 2018, 72, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Toplak, M.E.; Dockstader, C.; Tannock, R. Temporal Information Processing in ADHD: Findings to Date and New Methods. J. Neurosci. Methods 2006, 151, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.; Livingstone, S.R.; Bosnyak, D.J.; Trainor, L.J. Body Sway Reflects Leadership in Joint Music Performance. Proc. Natl. Acad. Sci. USA 2017, 114, E4134–E4141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colley, I.; Varlet, M.; MacRitchie, J.; Keller, P.E. The Influence of a Conductor and Co-Performer on Auditory-Motor Synchronisation, Temporal Prediction, and Ancillary Entrainment in a Musical Drumming Task. Hum. Mov. Sci. 2020, 72, 102653. [Google Scholar] [CrossRef] [PubMed]
- Eerola, T.; Jakubowski, K.; Moran, N.; Keller, P.E.; Clayton, M. Shared Periodic Performer Movements Coordinate Interactions in Duo Improvisations. R. Soc. Open Sci. 2018, 5, 171520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellamil, M.; Berson, J.; Wong, J.; Buckley, L.; Margulies, D.S. One in the Dance: Musical Correlates of Group Synchrony in a Real-World Club Environment. PLoS ONE 2016, 11, e0164783. [Google Scholar] [CrossRef]
- Himberg, T.; Laroche, J.; Bigé, R.; Buchkowski, M.; Bachrach, A. Coordinated Interpersonal Behaviour in Collective Dance Improvisation: The Aesthetics of Kinaesthetic Togetherness. Behav. Sci. 2018, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, P. Ensemble Performance: Interpersonal Alignment of Musical Expression; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Reddish, P.; Bulbulia, J.; Fischer, R. Does Synchrony Promote Generalized Prosociality? Relig. Brain Behav. 2014, 4, 3–19. [Google Scholar] [CrossRef]
- Lucas, G.; Clayton, M.; Leante, L. Inter-Group Entrainment in Afro-Brazilian Congado Ritual. Empir. Musicol. Rev. 2011, 6, 75–102. [Google Scholar] [CrossRef] [Green Version]
- Turino, T. Music as Social Life: The Politics of Participation; University of Chicago Press: Chicago, IL, USA, 2008. [Google Scholar]
- Witek, M.A. Filling in: Syncopation, Pleasure and Distributed Embodiment in Groove. Music. Anal. 2017, 36, 138–160. [Google Scholar] [CrossRef]
- Higgins, K.M. The Music between Us; University of Chicago Press: Chicago, IL, USA, 2012. [Google Scholar]
- Mithen, S. The Music Instinct. Ann. N. Y. Acad. Sci. 2009, 1169, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Trevarthen, C. Communicative Musicality: The Human Impulse to Create and Share Music. In Musical Imaginations: Multidisciplinary Perspectives on Creativity, Performance, and Perception; Oxford University Press: Oxford, UK, 2012; pp. 259–284. [Google Scholar]
- Herholz, S.C.; Zatorre, R.J. Musical Training as a Framework for Brain Plasticity: Behavior, Function, and Structure. Neuron 2012, 76, 486–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merrett, D.L.; Peretz, I.; Wilson, S.J. Moderating Variables of Music Training-Induced Neuroplasticity: A Review and Discussion. Front. Psychol. 2013, 4, 606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraus, N.; Slater, J.; Thompson, E.C.; Hornickel, J.; Strait, D.L.; Nicol, T.; White-Schwoch, T. Music Enrichment Programs Improve the Neural Encoding of Speech in At-Risk Children. J. Neurosci. 2014, 34, 11913–11918. [Google Scholar] [CrossRef] [PubMed]
- Reybrouck, M.; Brattico, E. Neuroplasticity beyond Sounds: Neural Adaptations Following Long-Term Musical Aesthetic Experiences. Brain Sci. 2015, 5, 69–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babiloni, F.; Astolfi, L. Social Neuroscience and Hyperscanning Techniques: Past, Present and Future. Neurosci. Biobehav. Rev. 2014, 44, 76–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czeszumski, A.; Eustergerling, S.; Lang, A.; Menrath, D.; Gerstenberger, M.; Schuberth, S.; Schreiber, F.; Rendon, Z.Z.; König, P. Hyperscanning: A Valid Method to Study Neural Inter-Brain Underpinnings of Social Interaction. Front. Hum. Neurosci. 2020, 14, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, T.; Tanabe, H.C.; Sadato, N. Hyperscanning Neuroimaging Technique to Reveal the “Two-in-One” System in Social Interactions. Neurosci. Res. 2015, 90, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Balconi, M.; Vanutelli, M.E. Cooperation and Competition with Hyperscanning Methods: Review and Future Application to Emotion Domain. Front. Comput. Neurosci. 2017, 11, 86. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Cerritos, C.; Khan, F. Neural Mechanisms Underlying Interpersonal Coordination: A Review of Hyperscanning Research. Soc. Personal. Psychol. Compass 2018, 12, e12421. [Google Scholar] [CrossRef]
- Liu, D.; Liu, S.; Liu, X.; Zhang, C.; Li, A.; Jin, C.; Chen, Y.; Wang, H.; Zhang, X. Interactive Brain Activity: Review and Progress on EEG-Based Hyperscanning in Social Interactions. Front. Psychol. 2018, 9, 1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.-Y.; Luan, P.; Zhang, J.; Xiang, Y.-T.; Niu, H.; Yuan, Z. Concurrent Mapping of Brain Activation from Multiple Subjects during Social Interaction by Hyperscanning: A Mini-Review. Quant. Imaging Med. Surg. 2018, 8, 819. [Google Scholar] [CrossRef] [PubMed]
- Dikker, S.; Wan, L.; Davidesco, I.; Kaggen, L.; Oostrik, M.; McClintock, J.; Rowland, J.; Michalareas, G.; Van Bavel, J.J.; Ding, M.; et al. Brain-to-Brain Synchrony Tracks Real-World Dynamic Group Interactions in the Classroom. Curr. Biol. 2017, 27, 1375–1380. [Google Scholar] [CrossRef]
- Hasegawa, C.; Ikeda, T.; Yoshimura, Y.; Hiraishi, H.; Takahashi, T.; Furutani, N.; Hayashi, N.; Minabe, Y.; Hirata, M.; Asada, M.; et al. Mu Rhythm Suppression Reflects Mother-Child Face-to-Face Interactions: A Pilot Study with Simultaneous MEG Recording. Sci. Rep. 2016, 6, 34977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Dai, B.; Peng, D.; Zhu, C.; Liu, L.; Lu, C. Neural Synchronization during Face-to-Face Communication. J. Neurosci. 2012, 32, 16064–16069. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Mok, C.; Witt, E.E.; Pradhan, A.H.; Chen, J.E.; Reiss, A.L. NIRS-Based Hyperscanning Reveals Inter-Brain Neural Synchronization during Cooperative Jenga Game with Face-to-Face Communication. Front. Hum. Neurosci. 2016, 10, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balconi, M.; Vanutelli, M.E. EEG Hyperscanning and Behavioral Synchronization during a Joint Actions. Neuropsychol. Trends. 2018, 23–47. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Bryant, D.M.; Reiss, A.L. NIRS-Based Hyperscanning Reveals Increased Interpersonal Coherence in Superior Frontal Cortex during Cooperation. Neuroimage 2012, 59, 2430–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Sun, B.; Chen, C.; Li, W.; Wang, Y.; Zhang, W.; Xiao, W.; Shao, Y. Self–Other Overlap and Interpersonal Neural Synchronization Serially Mediate the Effect of Behavioral Synchronization on Prosociality. Soc. Cogn. Affect. Neurosci. 2020, 15, 203–214. [Google Scholar] [CrossRef]
- Fishburn, F.A.; Murty, V.P.; Hlutkowsky, C.O.; MacGillivray, C.E.; Bemis, L.M.; Murphy, M.E.; Huppert, T.J.; Perlman, S.B. Putting Our Heads Together: Interpersonal Neural Synchronization as a Biological Mechanism for Shared Intentionality. Soc. Cogn. Affect. Neurosci. 2018, 13, 841–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Han, Z.R.; Wang, H.; Hu, Y.; Wang, Q.; Feng, S.; Yi, L. The Relation of Parental Emotion Regulation to Child Autism Spectrum Disorder Core Symptoms: The Moderating Role of Child Cardiac Vagal Activity. Front. Psychol. 2018, 9, 2480. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, H.; Luo, H.; Zhang, X.; Zhang, R.; Li, X. Interpersonal Neural Synchronization During Cooperative Behavior of Basketball Players: A FNIRS-Based Hyperscanning Study. Front. Hum. Neurosci. 2020, 14, 169. [Google Scholar] [CrossRef] [PubMed]
- Toppi, J.; Borghini, G.; Petti, M.; He, E.J.; De Giusti, V.; He, B.; Astolfi, L.; Babiloni, F. Investigating Cooperative Behavior in Ecological Settings: An EEG Hyperscanning Study. PLoS ONE 2016, 11, e0154236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Meng, T.; Hou, Y.; Pan, Y.; Hu, Y. Interpersonal Brain Synchronization Associated with Working Alliance during Psychological Counseling. Psychiatry Res. Neuroimaging 2018, 282, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Schleihauf, H.; Kayhan, E.; Matthes, D.; Vrtička, P.; Hoehl, S. The Effects of Interaction Quality on Neural Synchrony during Mother-Child Problem Solving. Cortex 2020, 124, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Reindl, V.; Gerloff, C.; Scharke, W.; Konrad, K. Brain-to-Brain Synchrony in Parent-Child Dyads and the Relationship with Emotion Regulation Revealed by FNIRS-Based Hyperscanning. NeuroImage 2018, 178, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Kinreich, S.; Djalovski, A.; Kraus, L.; Louzoun, Y.; Feldman, R. Brain-to-Brain Synchrony during Naturalistic Social Interactions. Sci. Rep. 2017, 7, 1–12. [Google Scholar]
- Long, Y.; Zheng, L.; Zhao, H.; Zhou, S.; Zhai, Y.; Lu, C. Interpersonal Neural Synchronization during Interpersonal Touch Underlies Affiliative Pair Bonding between Romantic Couples. Cereb. Cortex 2021, 31, 1647–1659. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Guo, C.; Han, S. Oxytocin Enhances Inter-Brain Synchrony during Social Coordination in Male Adults. Soc. Cogn. Affect. Neurosci. 2016, 11, 1882–1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acquadro, M.A.; Congedo, M.; De Riddeer, D. Music Performance as an Experimental Approach to Hyperscanning Studies. Front. Hum. Neurosci. 2016, 10, 242. [Google Scholar] [CrossRef] [PubMed]
- Osaka, N.; Minamoto, T.; Yaoi, K.; Azuma, M.; Shimada, Y.M.; Osaka, M. How Two Brains Make One Synchronized Mind in the Inferior Frontal Cortex: FNIRS-Based Hyperscanning during Cooperative Singing. Front. Psychol. 2015, 6, 1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Song, B.; Hu, Y.; Pan, Y.; Hu, Y. The Averaged Inter-Brain Coherence between the Audience and a Violinist Predicts the Popularity of Violin Performance. Neuroimage 2020, 211, 116655. [Google Scholar] [CrossRef]
- Pan, Y.; Novembre, G.; Song, B.; Zhu, Y.; Hu, Y. Dual Brain Stimulation Enhances Interpersonal Learning through Spontaneous Movement Synchrony. Soc. Cogn. Affect. Neurosci. 2021, 16, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Babiloni, C.; Vecchio, F.; Infarinato, F.; Buffo, P.; Marzano, N.; Spada, D.; Rossi, S.; Bruni, I.; Rossini, P.M.; Perani, D. Simultaneous Recording of Electroencephalographic Data in Musicians Playing in Ensemble. Cortex 2011, 47, 1082–1090. [Google Scholar] [CrossRef] [PubMed]
- Babiloni, C.; Buffo, P.; Vecchio, F.; Marzano, N.; Del Percio, C.; Spada, D.; Rossi, S.; Bruni, I.; Rossini, P.M.; Perani, D. Brains “in Concert”: Frontal Oscillatory Alpha Rhythms and Empathy in Professional Musicians. Neuroimage 2012, 60, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Washburn, A.; Román, I.; Huberth, M.; Gang, N.; Dauer, T.; Reid, W.; Nanou, C.; Wright, M.; Fujioka, T. Musical Role Asymmetries in Piano Duet Performance Influence Alpha-Band Neural Oscillation and Behavioral Synchronization. Front. Neurosci. 2019, 13, 1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindenberger, U.; Li, S.-C.; Gruber, W.; Müller, V. Brains Swinging in Concert: Cortical Phase Synchronization while Playing Guitar. BMC Neurosci. 2009, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sänger, J.; Müller, V.; Lindenberger, U. Intra-and Interbrain Synchronization and Network Properties When Playing Guitar in Duets. Front. Hum. Neurosci. 2012, 6, 312. [Google Scholar] [CrossRef] [Green Version]
- Müller, V.; Sänger, J.; Lindenberger, U. Intra-and Inter-Brain Synchronization during Musical Improvisation on the Guitar. PLoS ONE 2013, 8, e73852. [Google Scholar] [CrossRef] [Green Version]
- Pérez, A.; Carreiras, M.; Duñabeitia, J.A. Brain-to-Brain Entrainment: EEG Interbrain Synchronization While Speaking and Listening. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymanski, C.; Pesquita, A.; Brennan, A.A.; Perdikis, D.; Enns, J.T.; Brick, T.R.; Müller, V.; Lindenberger, U. Teams on the Same Wavelength Perform Better: Inter-Brain Phase Synchronization Constitutes a Neural Substrate for Social Facilitation. Neuroimage 2017, 152, 425–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakatos, P.; Shah, A.S.; Knuth, K.H.; Ulbert, I.; Karmos, G.; Schroeder, C.E. An Oscillatory Hierarchy Controlling Neuronal Excitability and Stimulus Processing in the Auditory Cortex. J. Neurophysiol. 2005, 94, 1904–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Atteveldt, N.; Musacchia, G.; Zion-Golumbic, E.; Sehatpour, P.; Javitt, D.C.; Schroeder, C. Complementary FMRI and EEG Evidence for More Efficient Neural Processing of Rhythmic vs. Unpredictably Timed Sounds. Front. Psychol. 2015, 6, 1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrams, D.A.; Nicol, T.; Zecker, S.; Kraus, N. Right-Hemisphere Auditory Cortex Is Dominant for Coding Syllable Patterns in Speech. J. Neurosci. 2008, 28, 3958–3965. [Google Scholar] [PubMed]
- Giraud, A.-L.; Poeppel, D. Cortical Oscillations and Speech Processing: Emerging Computational Principles and Operations. Nat. Neurosci. 2012, 15, 511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Poeppel, D. Cortical Oscillations in Auditory Perception and Speech: Evidence for Two Temporal Windows in Human Auditory Cortex. Front. Psychol. 2012, 3, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golumbic, E.M.Z.; Poeppel, D.; Schroeder, C.E. Temporal Context in Speech Processing and Attentional Stream Selection: A Behavioral and Neural Perspective. Brain Lang. 2012, 122, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Lerousseau, J.P.; Trebuchon, A.; Morillon, B.; Schön, D. Frequency Selectivity of Persistent Cortical Oscillatory Responses to Auditory Rhythmic Stimulation. Journal of Neuroscience 2021, 41, 7991–8006. [Google Scholar]
- Rimmele, J.M.; Morillon, B.; Poeppel, D.; Arnal, L.H. Proactive Sensing of Periodic and Aperiodic Auditory Patterns. Trends Cogn. Sci. 2018, 22, 870–882. [Google Scholar] [CrossRef]
- Lloyd, D. The Musical Structure of Time in the Brain: Repetition, Rhythm, and Harmony in FMRI During Rest and Passive Movie Viewing. Front. Comput. Neurosci. 2020, 13, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardy, B.G.; Calabrese, C.; De Lellis, P.; Bourgeaud, S.; Colomer, C.; Pla, S.; di Bernardo, M. Moving in Unison after Perceptual Interruption. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Djalovski, A.; Dumas, G.; Kinreich, S.; Feldman, R. Human Attachments Shape Interbrain Synchrony toward Efficient Performance of Social Goals. Neuroimage 2021, 226, 117600. [Google Scholar] [CrossRef]
- Chi, Y.M.; Wang, Y.; Wang, Y.-T.; Jung, T.-P.; Kerth, T.; Cao, Y. A Practical Mobile Dry EEG System for Human Computer Interfaces. In Proceedings of the International Conference on Augmented Cognition; Springer: Berlin/Heidelberg, Germany, 2013; pp. 649–655. [Google Scholar]
- Oullier, O.; De Guzman, G.C.; Jantzen, K.J.; Lagarde, J.; Scott Kelso, J.A. Social Coordination Dynamics: Measuring Human Bonding. Soc. Neurosci. 2008, 3, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Repp, B.H.; Su, Y.-H. Sensorimotor Synchronization: A Review of Recent Research (2006–2012). Psychon. Bull. Rev. 2013, 20, 403–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaut, M.H.; Demartin, M.; Sanes, J.N. Brain Networks for Integrative Rhythm Formation. PLoS ONE 2008, 3, e2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strang, G. Wavelet Transforms versus Fourier Transforms. Bull. Am. Math. Soc. 1993, 28, 288–305. [Google Scholar] [CrossRef] [Green Version]
- Holschneider, M.; Kronland-Martinet, R.; Morlet, J.; Tchamitchian, P. A Real-Time Algorithm for Signal Analysis with the Help of the Wavelet Transform. In Wavelets; Springer: Berlin/Heidelberg, Germany, 1990; pp. 286–297. [Google Scholar]
- Schmidt, R.C.; Carello, C.; Turvey, M.T. Phase Transitions and Critical Fluctuations in the Visual Coordination of Rhythmic Movements between People. J. Exp. Psychol. Hum. Percept. Perform. 1990, 16, 227. [Google Scholar] [CrossRef] [PubMed]
- London, J. Hearing in Time: Psychological Aspects of Musical Meter; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Gugnowska, K.; Novembre, G.; Kohler, N.; Villringer, A.; Keller, P.E.; Sammler, D. Endogenous Sources of Interbrain Synchrony in Duetting Pianists. Cereb. Cortex 2022, bhab469. [Google Scholar] [CrossRef]
- Burgess, A.P. On the Interpretation of Synchronization in EEG Hyperscanning Studies: A Cautionary Note. Front. Hum. Neurosci. 2013, 7, 881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, K.; Hao, N. When Do We Fall in Neural Synchrony with Others? Soc. Cogn. Affect. Neurosci. 2019, 14, 253–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novembre, G.; Iannetti, G.D. Hyperscanning Alone Cannot Prove Causality. Multibrain Stimulation Can. Trends Cogn. Sci. 2021, 25, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.F.d.C. Hyperscanning: Beyond the Hype. Neuron 2021, 109, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Koban, L.; Ramamoorthy, A.; Konvalinka, I. Why Do We Fall into Sync with Others? Interpersonal Synchronization and the Brain’s Optimization Principle. Soc. Neurosci. 2019, 14, 1–9. [Google Scholar] [CrossRef]
- Mastroianni, A.M.; Gilbert, D.T.; Cooney, G.; Wilson, T.D. Do Conversations End When People Want Them To? Proc. Natl. Acad. Sci. USA 2021, 118, e2011809118. [Google Scholar] [CrossRef] [PubMed]
- Large, E.W.; Jones, M.R. The Dynamics of Attending: How People Track Time-Varying Events. Psychol. Rev. 1999, 106, 119. [Google Scholar] [CrossRef]
- De Graaf, T.A.; Gross, J.; Paterson, G.; Rusch, T.; Sack, A.T.; Thut, G. Alpha-Band Rhythms in Visual Task Performance: Phase-Locking by Rhythmic Sensory Stimulation. PLoS ONE 2013, 8, e60035. [Google Scholar] [CrossRef] [PubMed]
- Mathewson, K.E.; Prudhomme, C.; Fabiani, M.; Beck, D.M.; Lleras, A.; Gratton, G. Making Waves in the Stream of Consciousness: Entraining Oscillations in EEG Alpha and Fluctuations in Visual Awareness with Rhythmic Visual Stimulation. J. Cogn. Neurosci. 2012, 24, 2321–2333. [Google Scholar] [CrossRef]
- Spaak, E.; de Lange, F.P.; Jensen, O. Local Entrainment of Alpha Oscillations by Visual Stimuli Causes Cyclic Modulation of Perception. J. Neurosci. 2014, 34, 3536–3544. [Google Scholar] [CrossRef]
- Stupacher, J.; Witte, M.; Hove, M.J.; Wood, G. Neural Entrainment in Drum Rhythms with Silent Breaks: Evidence from Steady-State Evoked and Event-Related Potentials. J. Cogn. Neurosci. 2016, 28, 1865–1877. [Google Scholar] [CrossRef]
- Ross, J.M.; Iversen, J.R.; Balasubramaniam, R. Motor Simulation Theories of Musical Beat Perception. Neurocase 2016, 22, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Iversen, J.R.; Balasubramaniam, R. Synchronization and Temporal Processing. Curr. Opin. Behav. Sci. 2016, 8, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Iversen, J.; Repp, B.; Patel, A. Top-down Control of Rhythm Perception Modulates Early Auditory Responses. Ann. N. Y. Acad. Sci. 2009, 1169, 58–73. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalil, A.; Musacchia, G.; Iversen, J.R. It Takes Two: Interpersonal Neural Synchrony Is Increased after Musical Interaction. Brain Sci. 2022, 12, 409. https://doi.org/10.3390/brainsci12030409
Khalil A, Musacchia G, Iversen JR. It Takes Two: Interpersonal Neural Synchrony Is Increased after Musical Interaction. Brain Sciences. 2022; 12(3):409. https://doi.org/10.3390/brainsci12030409
Chicago/Turabian StyleKhalil, Alexander, Gabriella Musacchia, and John Rehner Iversen. 2022. "It Takes Two: Interpersonal Neural Synchrony Is Increased after Musical Interaction" Brain Sciences 12, no. 3: 409. https://doi.org/10.3390/brainsci12030409