
Threat Detection in Nearby Space Mobilizes Human Ventral Premotor Cortex, Intraparietal Sulcus, and Amygdala

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
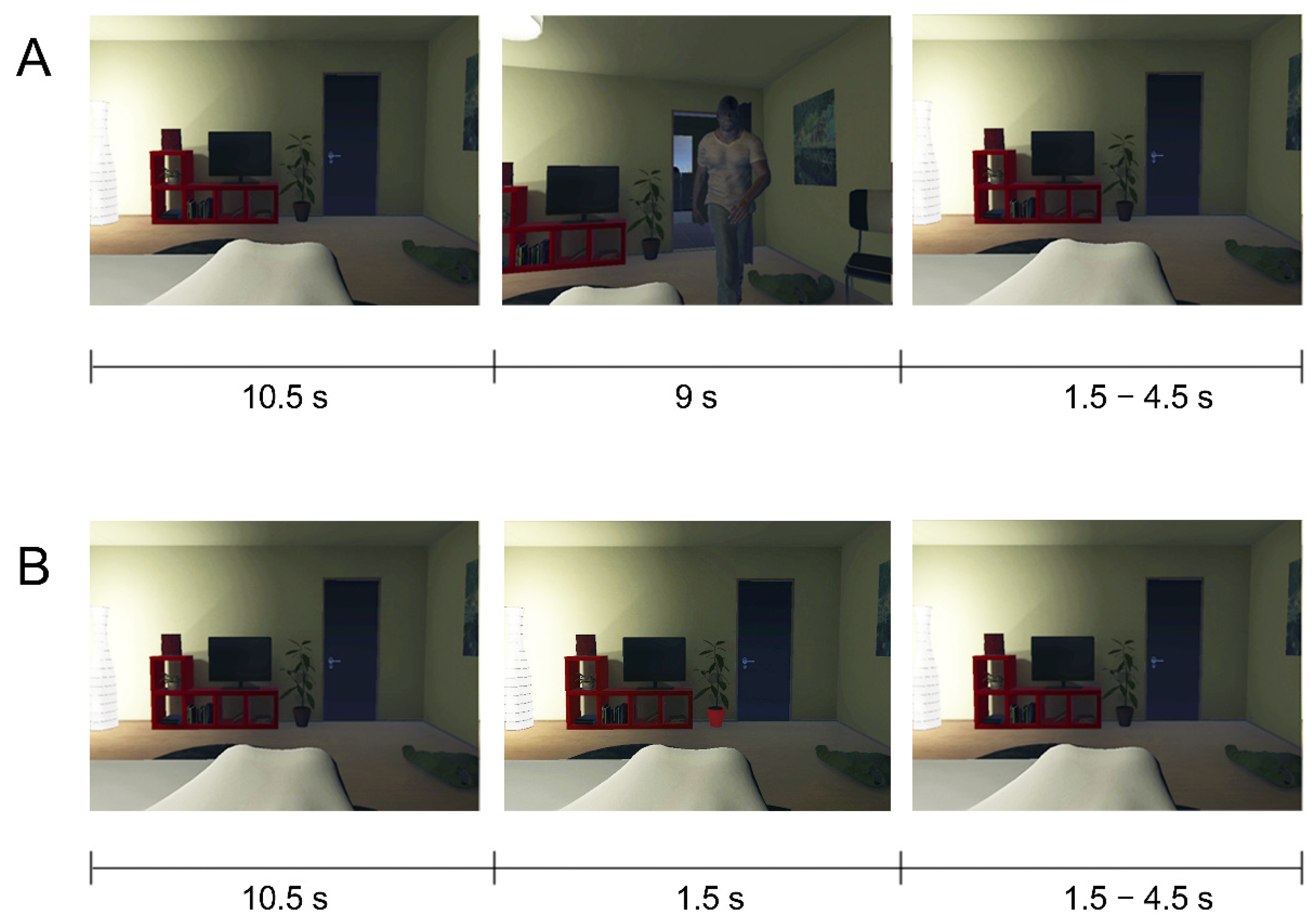
2.2. Stimuli and Materials
2.3. Procedure and Task
2.4. Design
2.5. Data Acquisition
2.6. Data Analyses
2.6.1. Functional MRI Pre-Processing
2.6.2. Behavioral Statistical Analyses
2.6.3. Functional MRI Statistical Analyses
3. Results
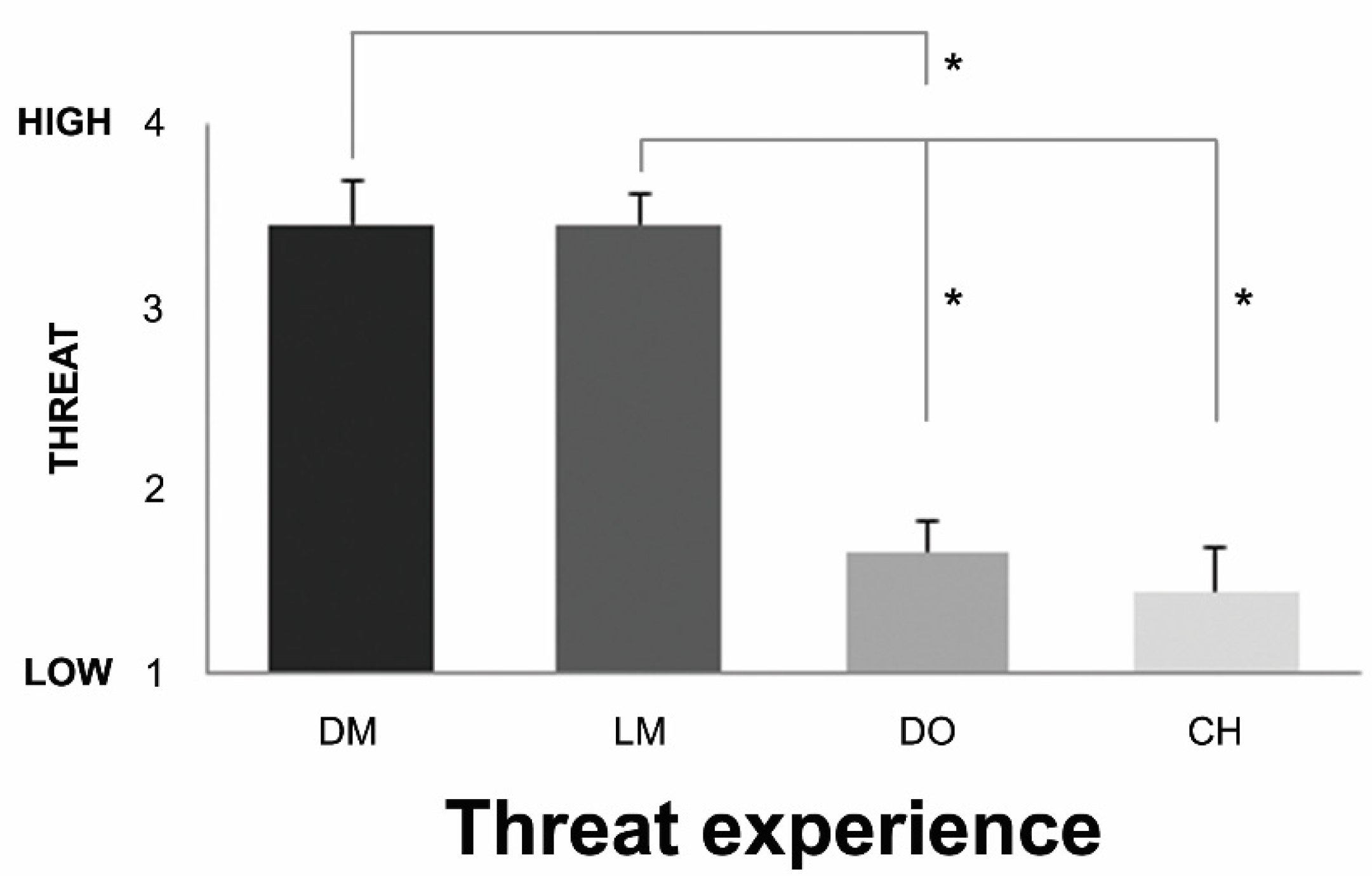
3.1. Behavioral Results
3.2. Visual Looming in Nearby Space
3.3. Threat Perception in Nearby Space
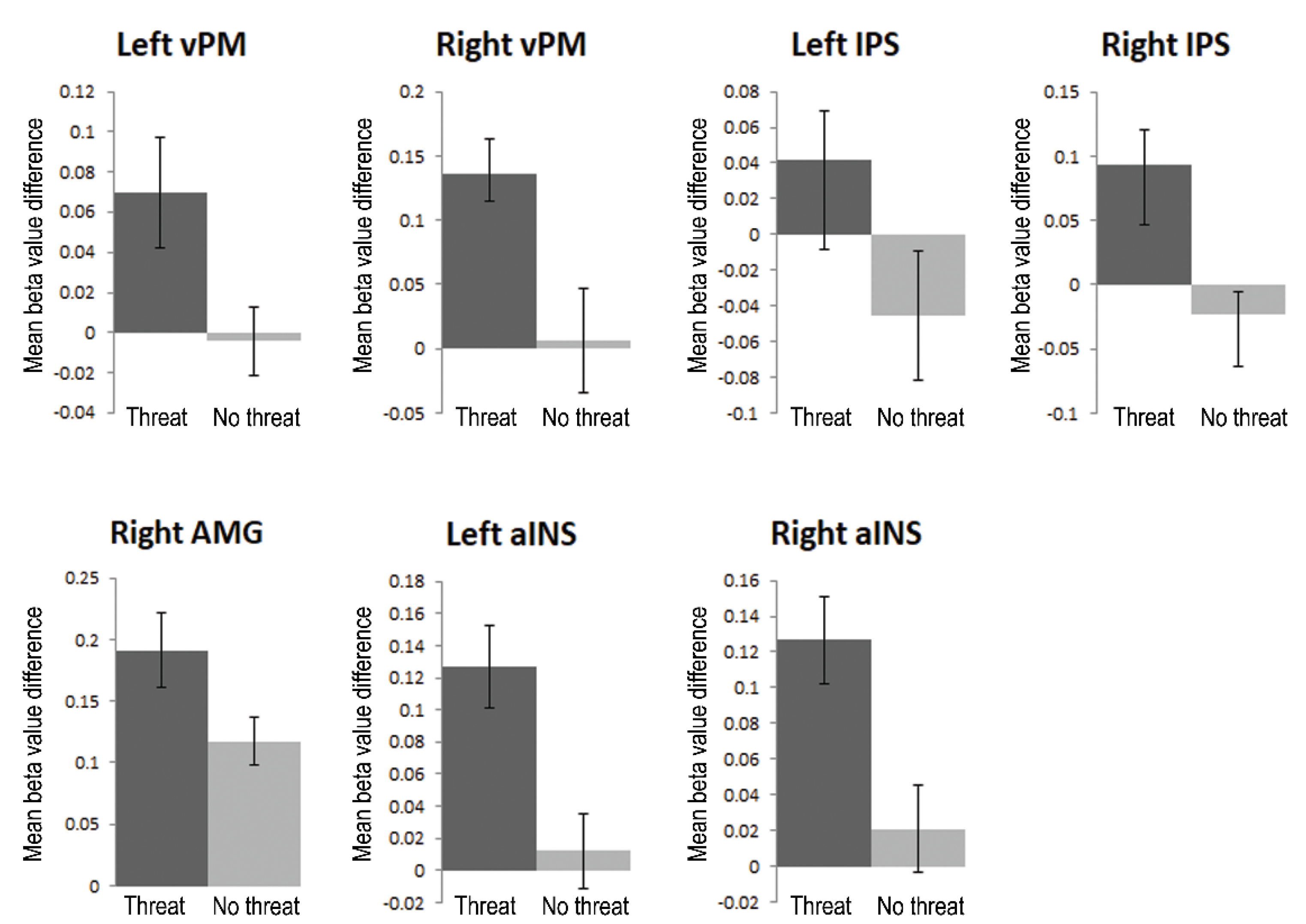
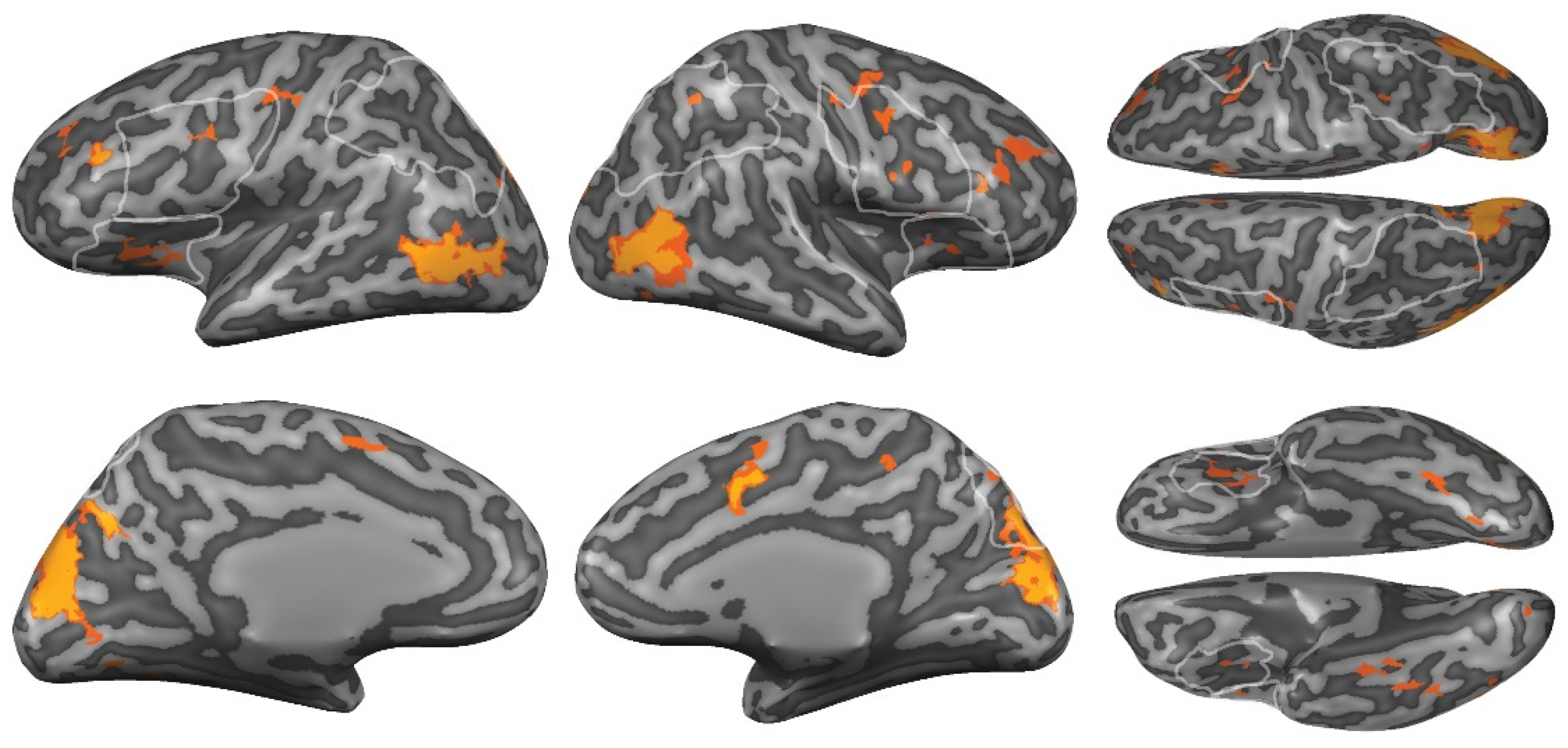
3.3.1. Region-of-Interest Analyses
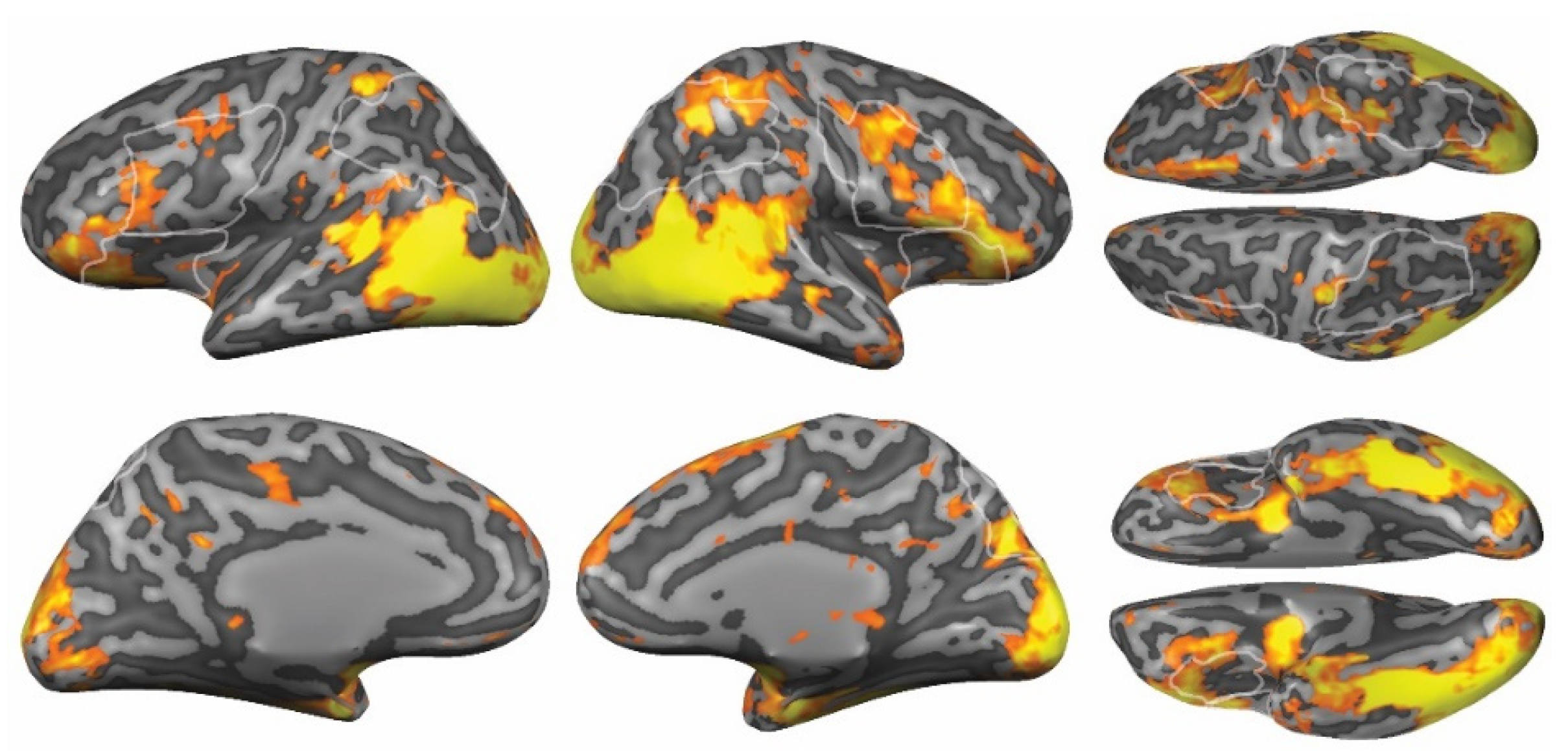
3.3.2. Whole-Brain Analyses
4. Discussion
4.1. Defensive Behavior in Peripersonal Space
4.2. Behavioral Relevance
4.3. Naturalistic Approaches
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Below You See Stills from the Events You Observed in the Scanner. Please RANK These Events on Perceived Threat, from Low (1) to High (4) Threat. | ||||
|---|---|---|---|---|
| 1 (Low Threat) | 2 | 3 | 4 (High Threat) | |
| Man A | x | |||
| Child | x | |||
| Dog | x | |||
| Man B | x | |||
| Questions | Mean Response and Standard Error |
|---|---|
| 1. To what extend did you feel as if you were in the room and lived the situation as if it were real? | 4.00 ± 0.74 |
| 2. Although you knew you were not there, to what extent did you have the illusion as if you were in the room? | 3.89 ± 0.70 |
| 3. To what extent did you think things like “I know this isn’t real”, but then surprisingly finding yourself reacting as if it was real? | 3.67 ± 0.74 |
| 4. To what extent was your sense of being in the room stronger than your sense of being in the scanner? | 3.44 ± 0.58 |
| 5. To what extent were your emotional responses during the events in the room similar to a real situation? | 3.22 ± 0.72 |
| 6. To what extent were the thoughts that you had during the events in the room similar to a real situation? | 3.33 ± 0.79 |
| 7. To what extent were the physical responses that you had during the events in the room similar to a real situation? | 3.44 ± 0.84 |
Appendix B
| Avatar Presence | ||||
|---|---|---|---|---|
| Region | Peak Voxel Coordinates | Cluster Size | ||
| x | y | z | ||
| Left inferior frontal gyrus | −47 | 21 | 40 | 6545 |
| Left inferior frontal gyrus | −45 | 41 | 4 | 3442 |
| Right inferior frontal gyrus | 55 | 27 | 24 | 6033 |
| Medial superior frontal gyrus | 9 | 9 | 64 | 9645 |
| Anterior medial superior frontal gyrus | −3 | 43 | 40 | 14,724 |
| Left premotor cortex | −41 | 9 | 52 | 3141 |
| Right premotor cortex | 37 | −5 | 40 | 6244 |
| Left insula | −41 | 21 | −4 | 10,745 |
| Right insula | 51 | 23 | 6 | 11,803 |
| Left superior parietal lobe | −55 | −57 | 30 | 2255 |
| Left intraparietal sulcus | −35 | −43 | 52 | 3072 |
| Right intraparietal sulcus | 29 | −53 | 48 | 8015 |
| Right precuneus | 17 | −79 | 34 | 4539 |
| Right supramarginal gyrus | 47 | −37 | 16 | 6454 |
| Left lateral sulcus | −55 | −41 | 16 | 1281 |
| Left mid cingulate sulcus | −15 | −21 | 38 | 1186 |
| Medial posterior cingulate sulcus | −1 | −47 | 24 | 2275 |
| Left parahippocampal gyrus | −35 | −7 | −20 | 2160 |
| Right parahippocampal gyrus | 37 | −3 | −24 | 2002 |
| Left occipito-temporal gyrus | −37 | −45 | −10 | 7223 |
| Right occipito-temporal gyrus | 37 | −41 | −14 | 7618 |
| Left middle occipital gyrus | −45 | −69 | −4 | 18,119 |
| Right middle occipital gyrus | 41 | −67 | 0 | 18,492 |
| Left middle occipital gyrus | −15 | −99 | 6 | 18,119 |
| Right middle occipital gyrus | 13 | −97 | 4 | 18,492 |
| Left inferior occipital gyrus | −43 | −81 | −2 | 11,367 |
| Right inferior occipital gyrus | 41 | −75 | −6 | 13,711 |
| Left posterior cingulate sulcus | −53 | −41 | 20 | 1979 |
| Left amygdala | −25 | −3 | −12 | 2745 |
| Right amygdala | 17 | −5 | −10 | 2590 |
| Left pulvinar | −19 | −25 | −2 | 1638 |
| Right pulvinar | 15 | −25 | 2 | 1876 |
| Threat Perception | ||||
|---|---|---|---|---|
| Region | Peak Voxel Coordinates | Cluster Size | ||
| x | y | z | ||
| Left middle frontal gyrus | −39 | 29 | 32 | 180 |
| Right anterior cingulate cortex | 9 | 3 | 40 | 167 |
| Left middle occipital gyrus | −49 | −77 | 6 | 2302 |
| Right middle occipital gyrus | 39 | −63 | 8 | 2941 |
| Left cuneus | −7 | −81 | 24 | 5965 |
| Right cuneus | 1 | −79 | 14 | 6149 |
References
- Rizzolatti, G.; Scandolara, C.; Matelli, M.; Gentilucci, M. Afferent properties of periarcuate neurons in macaque monkeys. II. Visual responses. Behav. Brain Res. 1981, 2, 147–163. [Google Scholar] [CrossRef]
- Bufacchi, R.J.; Iannetti, G.D. An Action Field Theory of Peripersonal Space. Trends Cogn. Sci. 2018, 22, 1076–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gelder, B. Towarsds the neurobiology of emotional body language. Nat. Rev. Neurosci. 2006, 7, 242–249. [Google Scholar] [CrossRef]
- Haxby, J.V.; Hoffman, E.A.; Gobbini, M.I. Human neural systems for face recognition and social communication. Biol. Psychiatry 2002, 51, 59–67. [Google Scholar] [CrossRef]
- Bignall, K.E.; Imbert, M. Polysensory and cortico-cortical projections to frontal lobe of squirrel and rhesus monkeys. Electroencephalogr. Clin. Neurophysiol. 1969, 26, 206–215. [Google Scholar] [CrossRef]
- Colby, C.L.; Duhamel, J.R.; Goldberg, M.E. Ventral intraparietal area of the macaque: Anatomic location and visual response properties. J. Neurophysiol. 1993, 69, 902–914. [Google Scholar] [CrossRef]
- Duhamel, J.R.; Colby, C.L.; Goldberg, M.E. Ventral intraparietal area of the macaque: Congruent visual and somatic response properties. J. Neurophysiol. 1998, 79, 126–136. [Google Scholar] [CrossRef]
- Graziano, M.S.; Yap, G.S.; Gross, C.G. Coding of visual space by premotor neurons. Science 1994, 266, 1054–1057. [Google Scholar] [CrossRef]
- Schlack, A.; Sterbing-D’Angelo, S.J.; Hartung, K.; Hoffmann, K.P.; Bremmer, F. Multisensory space representations in the macaque ventral intraparietal area. J. Neurosci. 2005, 25, 4616–4625. [Google Scholar] [CrossRef] [Green Version]
- Cooke, D.F.; Graziano, M.S. Defensive movements evoked by air puff in monkeys. J. Neurophysiol. 2003, 90, 3317–3329. [Google Scholar] [CrossRef]
- Cooke, D.F.; Graziano, M.S. Sensorimotor integration in the precentral gyrus: Polysensory neurons and defensive movements. J. Neurophysiol. 2004, 91, 1648–1660. [Google Scholar] [CrossRef] [PubMed]
- Cooke, D.F.; Taylor, C.S.; Moore, T.; Graziano, M.S. Complex movements evoked by microstimulation of the ventral intraparietal area. Proc. Natl. Acad. Sci. USA 2003, 100, 6163–6168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brozzoli, C.; Gentile, G.; Ehrsson, H.H. That’s near my hand! Parietal and premotor coding of hand-centered space contributes to localization and self-attribution of the hand. J. Neurosci. 2012, 32, 14573–14582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentile, G.; Petkova, V.I.; Ehrsson, H.H. Integration of visual and tactile signals from the hand in the human brain: An FMRI study. J. Neurophysiol. 2011, 105, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.S.; Chen, C.F.; Tran, A.T.; Holstein, K.L.; Sereno, M.I. Mapping multisensory parietal face and body areas in humans. Proc. Natl. Acad. Sci. USA 2012, 109, 18114–18119. [Google Scholar] [CrossRef] [Green Version]
- Makin, T.R.; Holmes, N.P.; Zohary, E. Is that near my hand? Multisensory representation of peripersonal space in human intraparietal sulcus. J. Neurosci. 2007, 27, 731–740. [Google Scholar] [CrossRef]
- Sereno, M.I.; Huang, R.S. A human parietal face area contains aligned head-centered visual and tactile maps. Nat. Neurosci. 2006, 9, 1337–1343. [Google Scholar] [CrossRef]
- Holt, D.J.; Cassidy, B.S.; Yue, X.; Rauch, S.L.; Boeke, E.A.; Nasr, S.; Tootell, R.B.; Coombs, G., III. Neural correlates of personal space intrusion. J. Neurosci. 2014, 34, 4123–4134. [Google Scholar] [CrossRef] [Green Version]
- Bach, D.R.; Schachinger, H.; Neuhoff, J.G.; Esposito, F.; Di Salle, F.; Lehmann, C.; Herdener, M.; Scheffler, K.; Seifritz, E. Rising sound intensity: An intrinsic warning cue activating the amygdala. Cereb. Cortex 2008, 18, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.S.; Chen, C.F.; Sereno, M.I. Spatiotemporal integration of looming visual and tactile stimuli near the face. Hum. Brain Mapp. 2018, 39, 2156–2176. [Google Scholar] [CrossRef] [Green Version]
- Bremmer, F.; Schlack, A.; Shah, N.J.; Zafiris, O.; Kubischik, M.; Hoffmann, K.; Zilles, K.; Fink, G.R. Polymodal motion processing in posterior parietal and premotor cortex: A human fMRI study strongly implies equivalencies between humans and monkeys. Neuron 2001, 29, 287–296. [Google Scholar] [CrossRef]
- Riskind, J.H.; Moore, R.; Bowley, L. The looming of spiders: The fearful perceptual distortion of movement and menace. Behav. Res. Ther. 1995, 33, 171–178. [Google Scholar] [CrossRef]
- Brendel, E.; DeLucia, P.R.; Hecht, H.; Stacy, R.L.; Larsen, J.T. Threatening pictures induce shortened time-to-contact estimates. Atten. Percept. Psychophys. 2012, 74, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Vagnoni, E.; Lourenco, S.F.; Longo, M.R. Threat modulates perception of looming visual stimuli. Curr. Biol. 2012, 22, R826–R827. [Google Scholar] [CrossRef] [Green Version]
- Ahs, F.; Dunsmoor, J.E.; Zielinski, D.; LaBar, K.S. Spatial proximity amplifies valence in emotional memory and defensive approach-avoidance. Neuropsychologia 2015, 70, 476–485. [Google Scholar] [CrossRef] [Green Version]
- Cartaud, A.; Ruggiero, G.; Ott, L.; Iachini, T.; Coello, Y. Physiological Response to Facial Expressions in Peripersonal Space Determines Interpersonal Distance in a Social Interaction Context. Front. Psychol. 2018, 9, 657. [Google Scholar] [CrossRef]
- Fossataro, C.; Sambo, C.F.; Garbarini, F.; Iannetti, G.D. Interpersonal interactions and empathy modulate perception of threat and defensive responses. Sci. Rep. 2016, 6, 19353. [Google Scholar] [CrossRef] [Green Version]
- Pellencin, E.; Paladino, M.P.; Herbelin, B.; Serino, A. Social perception of others shapes one’s own multisensory peripersonal space. Cortex 2018, 104, 163–179. [Google Scholar] [CrossRef]
- Ruggiero, G.; Frassinetti, F.; Coello, Y.; Rapuano, M.; di Cola, A.S.; Iachini, T. The effect of facial expressions on peripersonal and interpersonal spaces. Psychol. Res. 2017, 81, 1232–1240. [Google Scholar] [CrossRef]
- Teneggi, C.; Canzoneri, E.; di Pellegrino, G.; Serino, A. Social modulation of peripersonal space boundaries. Curr. Biol. 2013, 23, 406–411. [Google Scholar] [CrossRef] [Green Version]
- De Gelder, B.; Katsyri, J.; de Borst, A.W. Virtual reality and the new psychophysics. Br. J. Psychol. 2018, 109, 421–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Hamilton, A.F.C. Why and how to use virtual reality to study human social interaction: The challenges of exploring a new research landscape. Br. J. Psychol. 2018, 109, 395–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slater, M. Immersion and the illusion of presence in virtual reality. Br. J. Psychol. 2018, 109, 431–433. [Google Scholar] [CrossRef] [PubMed]
- Hindi Attar, C.; Muller, M.M.; Andersen, S.K.; Buchel, C.; Rose, M. Emotional processing in a salient motion context: Integration of motion and emotion in both V5/hMT+ and the amygdala. J. Neurosci. 2010, 30, 5204–5210. [Google Scholar] [CrossRef]
- Lloyd, D.; Morrison, I.; Roberts, N. Role for human posterior parietal cortex in visual processing of aversive objects in peripersonal space. J. Neurophysiol. 2006, 95, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Ousdal, O.T.; Jensen, J.; Server, A.; Hariri, A.R.; Nakstad, P.H.; Andreassen, O.A. The human amygdala is involved in general behavioral relevance detection: Evidence from an event-related functional magnetic resonance imaging Go-NoGo task. Neuroscience 2008, 156, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Billington, J.; Wilkie, R.M.; Field, D.T.; Wann, J.P. Neural processing of imminent collision in humans. Proc. R. Soc. B Biol. Sci. 2011, 278, 1476–1481. [Google Scholar] [CrossRef] [Green Version]
- Coccaro, E.F.; McCloskey, M.S.; Fitzgerald, D.A.; Phan, K.L. Amygdala and orbitofrontal reactivity to social threat in individuals with impulsive aggression. Biol. Psychiatry 2007, 62, 168–178. [Google Scholar] [CrossRef]
- Da Cunha-Bang, S.; Fisher, P.M.; Hjordt, L.V.; Perfalk, E.; Persson Skibsted, A.; Bock, C.; Ohlhues Baandrup, A.; Deen, M.; Thomsen, C.; Sestoft, D.M.; et al. Violent offenders respond to provocations with high amygdala and striatal reactivity. Soc. Cogn. Affect. Neurosci. 2017, 12, 802–810. [Google Scholar] [CrossRef] [Green Version]
- Dambacher, F.; Sack, A.T.; Lobbestael, J.; Arntz, A.; Brugman, S.; Schuhmann, T. Out of control: Evidence for anterior insula involvement in motor impulsivity and reactive aggression. Soc. Cogn. Affect. Neurosci. 2015, 10, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Statistics South Africa. Victims of Crime Survey 2015/16; Statistics South Africa: Pretoria, South Africa, 2017. Available online: https://www.statssa.gov.za/publications/P0341/P03412015.pdf (accessed on 25 February 2022).
- Goebel, R.; Esposito, F.; Formisano, E. Analysis of functional image analysis contest (FIAC) data with brainvoyager QX: From single-subject to cortically aligned group general linear model analysis and self-organizing group independent component analysis. Hum. Brain Mapp. 2006, 27, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Talairach, J.; Tournoux, P. Co-Planar Stereotaxic Atlas of the Human Brain: 3-Dimensional Proportional System: An Approach to Medical Cerebral Imaging; Thieme: New York, NY, USA, 1988. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Society. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- De Borst, A.W.; Sanchez-Vives, M.V.; Slater, M.; de Gelder, B. First-Person Virtual Embodiment Modulates the Cortical Network that Encodes the Bodily Self and Its Surrounding Space during the Experience of Domestic Violence. eNeuro 2020, 7, ENEURO.0263-19.2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graziano, M.S.; Cooke, D.F. Parieto-frontal interactions, personal space, and defensive behavior. Neuropsychologia 2006, 44, 2621–2635. [Google Scholar] [CrossRef] [PubMed]
- Coello, Y.; Bourgeois, J.; Iachini, T. Embodied perception of reachable space: How do we manage threatening objects? Cogn. Processing 2012, 13 (Suppl. S1), S131–S135. [Google Scholar] [CrossRef] [PubMed]
- Ferri, F.; Costantini, M.; Huang, Z.; Perrucci, M.G.; Ferretti, A.; Romani, G.L.; Northoff, G. Intertrial Variability in the Premotor Cortex Accounts for Individual Differences in Peripersonal Space. J. Neurosci. 2015, 35, 16328–16339. [Google Scholar] [CrossRef] [Green Version]
- Guterstam, A.; Bjornsdotter, M.; Gentile, G.; Ehrsson, H.H. Posterior cingulate cortex integrates the senses of self-location and body ownership. Curr. Biol. 2015, 25, 1416–1425. [Google Scholar] [CrossRef] [Green Version]
- Rossetti, A.; Romano, D.; Bolognini, N.; Maravita, A. Dynamic expansion of alert responses to incoming painful stimuli following tool use. Neuropsychologia 2015, 70, 486–494. [Google Scholar] [CrossRef]
- De Haan, A.M.; Smit, M.; van der Stigchel, S.; Dijkerman, H.C. Approaching threat modulates visuotactile interactions in peripersonal space. Exp. Brain Res. 2016, 234, 1875–1884. [Google Scholar] [CrossRef] [Green Version]
- Poliakoff, E.; Miles, E.; Li, X.; Blanchette, I. The effect of visual threat on spatial attention to touch. Cognition 2007, 102, 405–414. [Google Scholar] [CrossRef]
- Vagnoni, E.; Lourenco, S.F.; Longo, M.R. Threat modulates neural responses to looming visual stimuli. Eur. J. Neurosci. 2015, 42, 2190–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyll, S.; Bonath, B.; Schoenfeld, M.A.; Heinze, H.J.; Ohl, F.W.; Noesselt, T. Neural basis of multisensory looming signals. Neuroimage 2013, 65, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Bach, D.R.; Hurlemann, R.; Dolan, R.J. Impaired threat prioritisation after selective bilateral amygdala lesions. Cortex 2015, 63, 206–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mobbs, D.; Yu, R.; Rowe, J.B.; Eich, H.; FeldmanHall, O.; Dalgleish, T. Neural activity associated with monitoring the oscillating threat value of a tarantula. Proc. Natl. Acad. Sci. USA 2010, 107, 20582–20586. [Google Scholar] [CrossRef] [Green Version]
- Feinstein, J.S.; Adolphs, R.; Damasio, A.; Tranel, D. The human amygdala and the induction and experience of fear. Curr. Biol. 2011, 21, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Hadjikhani, N.; de Gelder, B. Seeing fearful body expressions activates the fusiform cortex and amygdala. Curr. Biol. 2003, 13, 2201–2205. [Google Scholar] [CrossRef]
- LeDoux, J. The emotional brain, fear, and the amygdala. Cell. Mol. Neurobiol. 2003, 23, 727–738. [Google Scholar] [CrossRef]
- De Borst, A.W.; de Gelder, B. Clear signals or mixed messages: Inter-individual emotion congruency modulates brain activity underlying affective body perception. Soc. Cogn. Affect. Neurosci. 2016, 11, 1299–1309. [Google Scholar] [CrossRef] [Green Version]
- Rudrauf, D.; David, O.; Lachaux, J.P.; Kovach, C.K.; Martinerie, J.; Renault, B.; Damasio, A. Rapid interactions between the ventral visual stream and emotion-related structures rely on a two-pathway architecture. J. Neurosci. 2008, 28, 2793–2803. [Google Scholar] [CrossRef] [Green Version]
- Mobbs, D.; Petrovic, P.; Marchant, J.L.; Hassabis, D.; Weiskopf, N.; Seymour, B.; Dolan, R.J.; Frith, C.D. When fear is near: Threat imminence elicits prefrontal-periaqueductal gray shifts in humans. Science 2007, 317, 1079–1083. [Google Scholar] [CrossRef] [Green Version]
- Terburg, D.; Scheggia, D.; Triana Del Rio, R.; Klumpers, F.; Ciobanu, A.C.; Morgan, B.; Montoya, E.R.; Bos, P.A.; Giobellina, G.; van den Burg, E.H.; et al. The Basolateral Amygdala Is Essential for Rapid Escape: A Human and Rodent Study. Cell 2018, 175, 723–735.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killgore, W.D.; Yurgelun-Todd, D.A. The right-hemisphere and valence hypotheses: Could they both be right (and sometimes left)? Soc. Cogn. Affect. Neurosci. 2007, 2, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Silberman, E.K.; Weingartner, H. Hemispheric lateralization of functions related to emotion. Brain Cogn. 1986, 5, 322–353. [Google Scholar] [CrossRef] [Green Version]
- Palomero-Gallagher, N.; Amunts, K. A short review on emotion processing: A lateralized network of neuronal networks. Brain Struct. Funct. 2022, 227, 673–684. [Google Scholar] [CrossRef]
- Craig, A.D. A new view of pain as a homeostatic emotion. Trends Neurosci. 2003, 26, 303–307. [Google Scholar] [CrossRef]
- Gonzalez-Franco, M.; Peck, T.C.; Rodriguez-Fornells, A.; Slater, M. A threat to a virtual hand elicits motor cortex activation. Exp. Brain Res. 2014, 232, 875–887. [Google Scholar] [CrossRef] [Green Version]
- Pichon, S.; de Gelder, B.; Grezes, J. Threat prompts defensive brain responses independently of attentional control. Cereb. Cortex 2012, 22, 274–285. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Vives, M.V.; Slater, M. From presence to consciousness through virtual reality. Nat. Rev. Neurosci. 2005, 6, 332–339. [Google Scholar] [CrossRef]
- Mello, M.; Dupont, L.; Engelen, T.; Acciarino, A.; de Borst, A.W.; de Gelder, B. Human freezing responses to virtual characters in immersive virtual reality are impacted by body expression, group affiliation and threat proximity. PsyArXiv 2021. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Borst, A.W.; de Gelder, B. Threat Detection in Nearby Space Mobilizes Human Ventral Premotor Cortex, Intraparietal Sulcus, and Amygdala. Brain Sci. 2022, 12, 391. https://doi.org/10.3390/brainsci12030391
de Borst AW, de Gelder B. Threat Detection in Nearby Space Mobilizes Human Ventral Premotor Cortex, Intraparietal Sulcus, and Amygdala. Brain Sciences. 2022; 12(3):391. https://doi.org/10.3390/brainsci12030391
Chicago/Turabian Stylede Borst, Aline W., and Beatrice de Gelder. 2022. "Threat Detection in Nearby Space Mobilizes Human Ventral Premotor Cortex, Intraparietal Sulcus, and Amygdala" Brain Sciences 12, no. 3: 391. https://doi.org/10.3390/brainsci12030391