
Depression-Associated Negr1 Gene-Deficiency Induces Alterations in the Monoaminergic Neurotransmission Enhancing Time-Dependent Sensitization to Amphetamine in Male Mice

 , ,
, ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
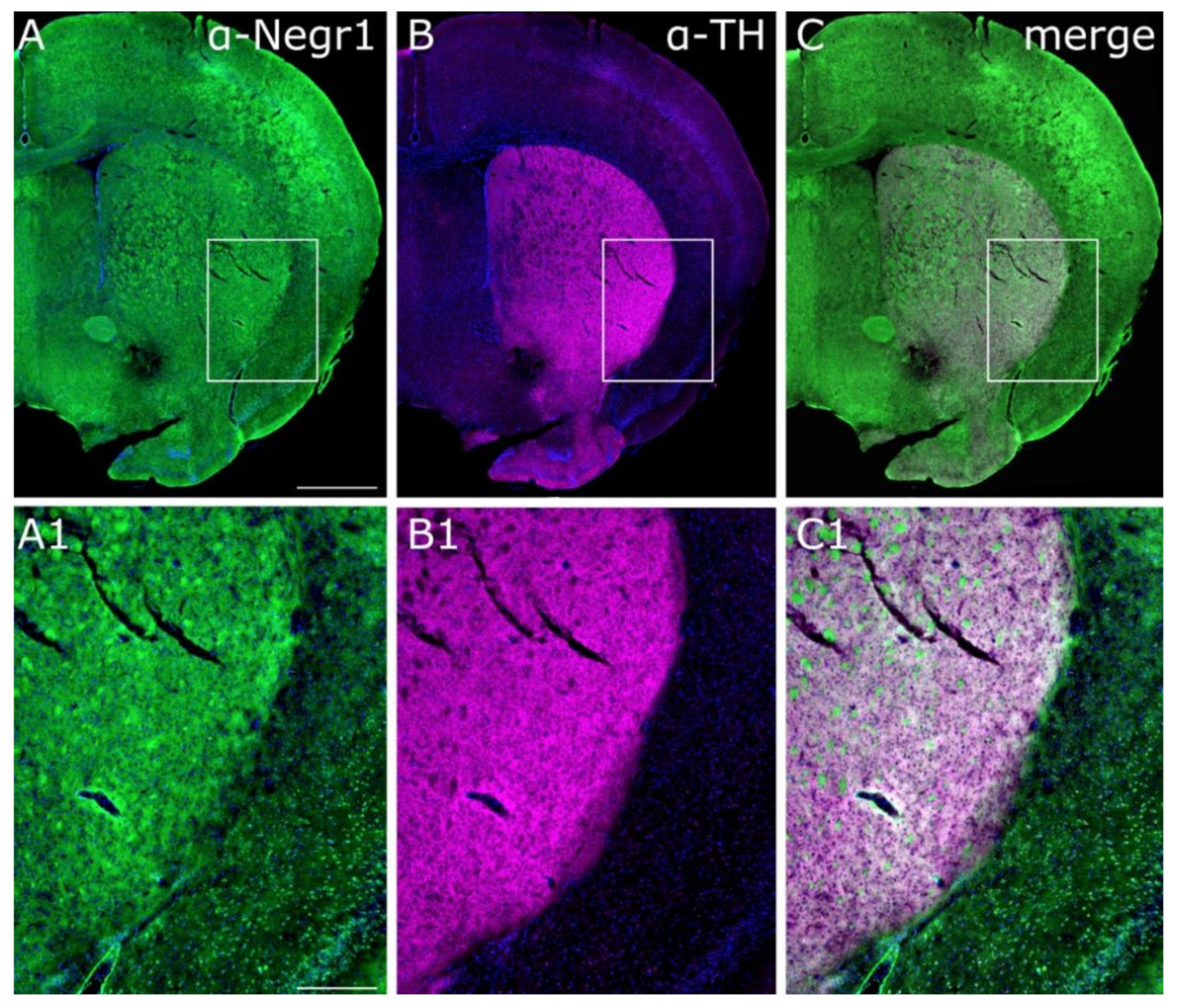{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Acute and Chronic Amphetamine Treatment
2.3. Escitalopram Treatment
2.4. Elevated plus Maze
2.5. Open Field Test
2.6. Tail Suspension Test
2.7. Measurement of Monoamines
2.8. Immunohistochemistry
2.9. RT-qPCR Analysis in Mouse Brain Areas
2.10. Statistical Analysis
3. Results
3.1. Negr1−/− Mice Display Higher Sensitivity to Amphetamine Compared to WT Mice
3.2. Chronic Administration of Amphetamine Increases the Level of Tyrosine Hydroxylase (TH) in VTA
3.3. Amphetamine Increases the Level of Dopamine in Dorsal Striatum (DSTR)
3.4. Chronic Amphetamine Administration Alters the Level of Monoamines in the Hippocampus and Chronic Escitalopram Treatment Causes Weight Difference in the Hippocampus
3.5. Chronic Administration of Escitalopram Alters the Level of Monoamines and Their Metabolites in Raphe
3.6. Chronic Administration of Escitalopram Causes No Alterations in the Behavior of Negr1−/− Mice
4. Discussion
4.1. NEGR1 Expression in Monoaminergic Brain Circuits
4.2. Increased Behavioral Sensitization to Amphetamine and Upregulation of Dat Transcript in Negr1−/− Mice
4.3. Altered Molecular Reactivity to the Amphetamine in the Brains of Negr1−/− Mice
4.4. Negr1−/− Mice Display Reduced Sensitivity to Experimental Manipulations and Show Less Activity during Chronic Injections/Testing
4.5. Escitalopram-Induced Reduction of 5-HT and DA Turnover Is Enhanced in Negr1−/− Mice
4.6. Negr1 Deficiency-Induced Alterations in the Monoaminergic Neurotransmission Could Explain Links of NEGR1 with Both Depression and Obesity Phenotypes
5. Conclusions
6. Limitations of the Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hyde, C.L.; Nagle, M.W.; Tian, C.; Chen, X.; Paciga, S.A.; Wendland, J.R.; Tung, J.Y.; Hinds, D.A.; Perlis, R.H.; Winslow, A.R. Identification of 15 genetic loci associated with risk of major depression in individuals of European descent. Nat. Genet. 2016, 48, 1031–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wray, N.R.; Ripke, S.; Mattheisen, M.; Trzaskowski, M.; Byrne, E.M.; Abdel, A.; Adams, M.J.; Agerbo, E.; Air, T.M.; Andlauer, T.M.F.; et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat. Genet. 2018, 50, 668–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, D.M.; Adams, M.J.; Shirali, M.; Clarke, T.K.; Marioni, R.E.; Davies, G.; Coleman, J.R.I.; Alloza, C.; Shen, X.; Barbu, M.C.; et al. Genome-wide association study of depression phenotypes in UK Biobank identifies variants in excitatory synaptic pathways. Nat. Commun. 2018, 9, 1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, D.M.; Adams, M.J.; Clarke, T.K.; Hafferty, J.D.; Gibson, J.; Shirali, M.; Coleman, J.R.I.; Hagenaars, S.P.; Ward, J.; Eigmore, E.M.; et al. Genome-wide meta-analysis of depression identifies 102 independent variants and highlights the importance of the prefrontal brain regions. Nat. Neurosci. 2019, 22, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levey, D.F.; Stein, M.B.; Wendt, F.R.; Pathak, G.A.; Zhou, H.; Aslan, M.; Quaden, R.; Harrington, K.M.; Nuñez, Y.Z.; Overstreet, C.; et al. Bi-ancestral depression GWAS in the Million Veteran Program and meta-analysis in >1.2 million individuals highlight new therapeutic directions. Nat. Neurosci. 2021, 24, 954–963. [Google Scholar] [CrossRef]
- Thorleifsson, G.; Walters, G.; Gudbjartsson, D.; Steinthorsdottir, V.; Sulem, P.; Helgadottir, A.; Strykarsdottir, U.; Gretarsdottir, S.; Thorlacius, S.; Jonsdottir, I.; et al. Genome-wide association yields new sequence variants at seven loci that associate with measures of obesity. Nat. Genet. 2009, 41, 18–24. [Google Scholar] [CrossRef]
- Speliotes, E.; Willer, C.; Berndt, S.; Monda, K.L.; Thorleifsson, G.; Jackson, A.U.; Allen, H.L.; Lindgren, C.M.; Luan, J.; Mägi, R.; et al. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat. Genet. 2010, 42, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, E.; Huang, N.; Bochukova, E.G.; Keogh, J.M.; Lindsay, S.; Grag, S.; Henning, E.; Blackburn, H.; Loos, R.J.F.; Wareham, N.J.; et al. Genome-wide SNP and CNV analysis identifies common and low-frequency variants associated with severe early-onset obesity. Nat. Genet. 2013, 45, 513–517. [Google Scholar] [CrossRef]
- Maccarrone, G.; Ditzen, C.; Yassouridis, A.; Rewerts, C.; Uhr, M.; Uhlen, M.; Holsboer, F.; Turck, C.W. Psychiatric patient stratification using biosignatures based on cerebrospinal fluid protein expression clusters. J. Psychiatr. Res. 2013, 47, 1572–1580. [Google Scholar] [CrossRef]
- Dall’Aglio, L.; Lewis, C.M.; Pain, O. Delineating the Genetic Component of Gene Expression in Major Depression. Biol. Psychiatry 2021, 89, 627–636. [Google Scholar] [CrossRef]
- Deng, Y.-T.; Ou, Y.-N.; Wu, B.-S.; Yang, Y.X.; Jiang, Y.; Huang, Y.-Y.; Liu, Y.; Tan, L.; Dong, Q.; Suckling, J.; et al. Identifying causal genes for depression via integration of the proteome and transcriptome from brain and blood. Mol. Psychiatry 2022, 27, 2849–2857. [Google Scholar] [CrossRef]
- Cross-Disorder Group of the Psychiatric Genomics Consortium. Genomic Relationships, Novel Loci, and Pleiotropic Mechanisms across Eight Psychiatric Disorders. Cell 2019, 179, 1469–1482.e11. [Google Scholar] [CrossRef] [Green Version]
- Cox, D.A.; Gottschalk, M.G.; Wesseling, H.; Ernst, A.; Cooper, J.D.; Bahn, S. Proteomic system evaluation of the molecular validity of preclinical psychosis models compared to schizophrenia brain pathology. Schizophr. Res. 2016, 177, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Karis, K.; Eskla, K.-L.; Kaare, M.; Täht, K.; Tuusov, J.; Visnapuu, T.; Innos, J.; Jayaram, M.; Timmusk, T.; Weickert, C.S.; et al. Altered Expression Profile of IgLON Family of Neural Cell Adhesion Molecules in the Dorsolateral Prefrontal Cortex of Schizophrenia Patients. Front. Mol. Neurosci. 2018, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- Sniekers, S.; Stringer, S.; Watanabe, K.; Jansen, P.R.; Coleman, J.R.I.; Krapohl, E.; Taskesen, E.; Hammerschlag, A.R.; Okbay, A.; Zabaneh, D.; et al. Genome-wide association meta-analysis of 78,308 individuals identifies new loci and genes influencing human intelligence. Nat. Genet. 2017, 49, 1107–1112. [Google Scholar] [CrossRef] [Green Version]
- Veerappa, A.M.; Saldanha, M.; Padakannaya, P.; Ramachandra, N.B. Family-based genome-wide copy number scan identifies five new genes of dyslexia involved in dendritic spinal plasticity. J. Hum. Genet. 2013, 58, 539–547. [Google Scholar] [CrossRef] [Green Version]
- Marshall, C.R.; Noor, A.; Vincent, J.B.; Lionel, A.C.; Feuk, L.; Skaug, J.; Shago, M.; Moessner, R.; Pinto, D.; Ren, Y.; et al. Structural variation of chromosomes in autism spectrum disorder. Am. J. Hum. Genet. 2008, 82, 477–488. [Google Scholar] [CrossRef] [Green Version]
- Michaelson, J.J.; Shi, Y.; Gujral, M.; Zheng, H.; Malhotra, D.; Jin, X.; Jian, M.; Liu, G.; Greer, D.; Bhandari, A.; et al. Whole-genome sequencing in autism identifies hot spots for de novo germline mutation. Cell 2012, 151, 1431–1442. [Google Scholar] [CrossRef] [Green Version]
- Genovese, A.; Cox, D.M.; Butler, M.G. Partial Deletion of Chromosome 1p31.1 Including only the Neuronal Growth Regulator 1 Gene in Two Siblings. J. Pediatr Genet. 2015, 4, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Vanaveski, T.; Singh, K.; Narvik, J.; Eskla, K.-L.; Visnapuu, T.; Heinla, I.; Jayaram, M.; Innos, J.; Lilleväli, K.; Philips, M.-A.; et al. Promoter-Specific Expression and Genomic Structure of IgLON Family Genes in Mouse. Front. Neurosci. 2017, 11, 38. [Google Scholar] [CrossRef]
- Ranaivoson, F.M.; Turk, L.S.; Ozgul, S.; Kakehi, S.; Daake, D.; Lopez, N.; Trobiani, L.; Jaco, A.D.; Denissova, N.; Demeler, B.; et al. A Proteomic Screen of Neuronal Cell-Surface Molecules Reveals IgLONs as Structurally Conserved Interaction. Modul. Synapse. Struct. 2019, 27, 893–906.e9. [Google Scholar] [CrossRef]
- Ramani, S.R.; Tom, I.; Lewin-Koh, N.; Wranik, B.; De Palastis, L.; Zhang, J.; Eaton, D.; Gonzalez, L.C. A secreted protein microarray platform for extracellular protein interaction discovery. Anal. Biochem. 2012, 420, 127–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourhaghighi, R.; Ash, P.E.A.; Phanse, S.; Goebels, F.; Hu, L.Z.M.; Chen, S.; Zhang, Y.; Wierbowski, S.D.; Boudeau, S.; Moutaoufjk, M.T.; et al. BraInMap Elucidates the Macromolecular Connectivity Landscape of Mammalian Brain. Cell Syst. 2020, 10, 333–350. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Lilleväli, K.; Gilbert, S.F.; Bregin, A.; Narvik, J.; Jayaram, M.; Rahi, M.; Innos, J.; Kaasik, A.; Vasar, E.; et al. The combined impact of IgLON family proteins Lsamp and Neurotrimin on developing neurons and behavioral profiles in mouse. Brain. Res. Bull. 2018, 140, 5–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T.; Yamada, M.; Maekawa, S.; Nakashima, T.; Miyata, S. IgLON cell adhesion molecule Kilon is a crucial modulator for synapse number in hippocampal neurons. Brain Res. 2008, 1224, 1–11. [Google Scholar] [CrossRef]
- Pischedda, F.; Szczurowska, J.; Cirnaru, M.D.; Gisert, F.; Vezzoli, E.; Ueffing, M.; Sala, C.; Francolini, M.; Hauck, S.M.; Cancedda, L.; et al. Cell Surface Biotinylation Assay to Reveal Membrane-associated Neuronal Cues: Negr1 Regulates Dendritic Arborization. Mol Cell Proteomics 2014, 13, 733–748. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Jayaram, M.; Kaare, M.; Leidmaa, E.; Jagomäe, T.; Heinla, I.; Hickey, M.A.; Kaasik, A.; Schäfer, M.K.; Innos, J.; et al. Neural cell adhesion molecule Negr1 deficiency in mouse results in structural brain endophenotypes and behavioral deviations related to psychiatric disorders. Sci. Rep. 2019, 9, 5457. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, M.; Bräuer, A.U.; Savaskan, N.E.; Rathjen, F.G.; Brümmendorf, T. Neurotractin/kilon promotes neurite outgrowth and is expressed on reactive astrocytes after entorhinal cortex lesion. Mol. Cell. Neurosci. 2005, 29, 580–590. [Google Scholar] [CrossRef]
- Singh, K.; Loreth, D.; Pöttker, B.; Hefti, K.; Innos, J.; Schwald, K.; Hengstler, H.; Menzel, L.; Sommer, C.J.; Radyushkin, K.; et al. Neuronal Growth and Behavioral Alterations in Mice Deficient for the Psychiatric Disease-Associated Negr1 Gene. Front. Mol. Neurosci. 2018, 11, 30. [Google Scholar] [CrossRef] [Green Version]
- Noh, K.; Lee, H.; Choi, T.-Y.; Joo, Y.; Kim, S.-J.; Kim, H.; Kim, J.Y.; Jahng, J.W.; Lee, S.; Choi, S.-Y.; et al. Negr1 controls adult hippocampal neurogenesis and affective behaviors. Mol. Psychiatry 2019, 24, 1189–1205. [Google Scholar] [CrossRef]
- Szczurkowska, J.; Pischedda, F.; Pinto, B.; Managò, F.; Haas, C.A.; Summa, M.; Bertorelli, R.; Papaleo, F.; Schäfer, M.K.; Piccoli, G.; et al. NEGR1 and FGFR2 cooperatively regulate cortical development and core behaviours related to autism disorders in mice. Brain 2018, 141, 2772–2794. [Google Scholar] [CrossRef]
- Kaare, M.; Mikheim, K.; Lilleväli, K.; Kilk, K.; Jagomäe, T.; Leidmaa, E.; Piirsalu, M.; Porosk, R.; Singh, K.; Reimets, R.; et al. High-Fat Diet Induces Pre-Diabetes and Distinct Sex-Specific Metabolic Alterations in Negr1-Deficient Mice. Biomedicine 2021, 9, 1148. [Google Scholar] [CrossRef]
- Belujon, P.; Grace, A.A. Dopamine System Dysregulation in Major Depressive Disorder. Int. J. Neuropsychophrmacolo. 2017, 20, 1036–1046. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, E.E.; Brignani, S.; Adolfs, Y.; Lemstra, S.; Demmers, J.; Vidaki, M.; Donahoo, A.-L.S.; Lilleväli, K.; Vasar, E.; Richards, L.J.; et al. Subdomain-Mediated Axon-Axon Signaling and Chemoattraction Cooperate to Regulate Afferent Innervation of the Lateral Habenula. Neuron 2014, 83, 372–387. [Google Scholar] [CrossRef] [Green Version]
- Inta, D.; Meyer-Lindenberg, A.; Gass, P. Alterations in postnatal neurogenesis and dopamine dysregulation in schizophrenia: A hypothesis. Schizophr Bull. 2010, 37, 674–680. [Google Scholar] [CrossRef] [Green Version]
- Okaty, B.W.; Freret, M.E.; Rood, B.D.; Brust, R.D.; Hennessy, M.L.; Bairos, D.; Kim, J.K.; Cook, M.N.; Dymecki, S.M. Multi-Scale Molecular Deconstruction of the Serotonin Neuron System. Neuron 2015, 88, 774–791. [Google Scholar] [CrossRef] [Green Version]
- Tamási, V.; Petschner, P.; Adori, C.; Kirilly, E.; Ando, R.D.; Tothfalusi, L.; Juhasz, G.; Bagdy, G. Transcriptional Evidence for the Role of Chronic Venlafaxine Treatment in Neurotrophic Signaling and Neuroplasticity Including also Glutatmatergic- and Insulin-Mediated Neuronal Processes. PLoS ONE 2014, 9, e113662. [Google Scholar] [CrossRef] [Green Version]
- Carboni, L.; Pischedda, F.; Piccoli, G.; Lauria, M.; Musazzi, L.; Popoli, M.; Mathé, A.A.; Domenici, E. Depression-Associated Gene Negr1-Fgfr2 Pathway Is Altered by Antidepressant Treatment. Cell 2020, 9, 1818. [Google Scholar] [CrossRef]
- Mustard, C.; Whitfield, P.; Megson, I.; Wei, J. P-1104–The Effect of Clozapine on the Expression of Obesity Genes. Eur. Psychiatry 2012, 27, 1. [Google Scholar] [CrossRef]
- Innos, J.; Leidmaa, E.; Philips, M.-A.; Sütt, S.; Alttoa, A.; Harro, J.; Kõks, S.; Vasar, E. Lsamp−/− mice display lower sensitivity to amphetamine and have elevated 5-HT turnover. Biochem. Biophys. Res. Commun. 2013, 430, 413–418. [Google Scholar] [CrossRef]
- Bregin, A.; Kaare, M.; Jagomäe, T.; Karis, K.; Singh, K.; Laugus, K.; Innos, J.; Leidmaa, E.; Heinla, I.; Visnapuu, T.; et al. Expression and impact of Lsamp neural adhesion molecule in the serotonergic neurotransmission system. Pharm. Biochem. Behav. 2020, 198, 173017. [Google Scholar] [CrossRef]
- Innos, J.; Philips, M.A.; Leidmaa, E.; Heinla, I.; Raud, S.; Reemann, P.; Plaas, M.; Nurk, K.; Kurrikoff, K.; Matto, V.; et al. Lower anxiety and a decrease in agonistic behaviour in Lsamp-deficient mice. Behav. Brain Res. 2011, 217, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Innos, J.; Philips, M.-A.; Raud, S.; Lilleväli, K.; Kõks, S.; Vasar, E. Deletion of the Lsamp gene lowers sensitivity to stressful environmental manipulations in mice. Behav. Brain Res. 2012, 228, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Mazitov, T.; Brefgin, A.; Philips, M.A.; Innos, J.; Vasar, E. Deficit in emotional learning in neurotrimin knockout mice. Behav. Brain Res. 2017, 28, 311–318. [Google Scholar] [CrossRef]
- Lee, A.W.S.; Hengstler, H.; Schwald, K.; Diaz, M.B.; Loreth, D.; Kirsch, M.; Kretz, O.; Haas, C.A.; de Angelis, M.H.; Herzig, S.; et al. Functional Inactivation of the Genome-Wide Association Study Obesity Gene Neuronal Growth Regulator 1 in Mice Causes a Body Mass Phenotype. PLoS ONE 2012, 7, e41537. [Google Scholar]
- Sokolov, B.P.; Schindler, C.W.; Cadet, J.L. Chronic methamphetamine increases fighting in mice. Pharmacol. Biochem. Behav. 2004, 77, 319–326. [Google Scholar] [CrossRef]
- Visnapuu, T.; Plaas, M.; Reimets, R.; Raud, S.; Terasmaa, A.; Kõks, S.; Sütt, S.; Luuk, H.; Hundahl, C.A.; Eskla, K.-L.; et al. Evidence of impaired function of dopaminergic system in Wfs1-deficient mice. Behav. Brain Res. 2013, 244, 90–99. [Google Scholar] [CrossRef]
- Vanaveski, T.; Narvik, J.; Innos, J.; Philips, M.-A.; Ottas, A.; Plaas, M.; Haring, L.; Zilmer, M.; Vasar, E. Repeated Administration of D-Amphetamine Induces Distinct Alterations in Behavior and Metabolite Level in 129Sv and Bl6 Mouse Strains. Front. Neurosci. 2018, 12, 399. [Google Scholar] [CrossRef] [Green Version]
- Franklin, K.B.J.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- Bakker, R.; Tiesinga, P.; Kötter, R. The Scalable Brain Atlas: Instant Web-Based Access to Public Brain Atlases and Related Content. Neuroinform 2015, 13, 353–366. [Google Scholar] [CrossRef] [Green Version]
- Varul, J.; Eskla, K.-L.; Piirsalu, M.; Innos, J.; Philips, M.-A.; Visnapuu, T.; Plaas, M.; Vasar, E. Dopamine System, NMDA Receptor and EGF Family Expressions in Brain Structures of Bl6 and 129Sv Strains Displaying Different Behavioral Adaptation. Brain Sci. 2021, 11, 725. [Google Scholar] [CrossRef]
- Scholl, J.L.; Feng, N.; Watt, M.J.; Renner, K.J.; Forster, G.L. Individual differences in amphetamine sensitization, behavior and central monoamines. Physiol Behav. 2009, 96, 493–504. [Google Scholar] [CrossRef]
- Geiger, B.M.; Haburcak, M.; Avena, N.M.; Moyer, M.C.; Hoebel, B.G.; Pothos, E.N. Deficits of mesolimbic dopamine neurotransmission in rat dietary obesity. Neuroscience 2009, 159, 1193–1199. [Google Scholar] [CrossRef] [Green Version]
- Sjödin, A.; Gasteyger, C.; Nielsen, A.L.; Raben, A.; Mikkelsen, J.D.; Jensen, J.K.S.; Meier, D.; Astrup, A. The effect of the triple monoamine reuptake inhibitor tesofensine on energy metabolism and appetite in overweight and moderately obese men. Int. J. Obes. 2010, 34, 1634–1643. [Google Scholar] [CrossRef] [Green Version]
- Jagomäe, T.; Singh, K.; Philips, M.A.; Jayaram, M.; Seppa, K.; Tekko, T.; Gilbert, S.F.; Vasar, E.; Lilleväli, K. Alternative Promoter Use Governs the Expression of IgLON Cell Adhesion Molecules in Histogenetic Fields of the Embryonic Mouse Brain. Int. J. Mol. Sci. 2021, 22, 6955. [Google Scholar] [CrossRef]
- Venkannagari, H.; Kasper, J.M.; Misra, A.; Rush, S.A.; Fan, S.; Lee, H.; Sun, H.; Seshadrinathan, S.; Machius, M.; Hommel, J.D.; et al. Highly Conserved Molecular Features in IgLONs Contrast Their Distinct Structural and Biological Outcomes. J. Mol. Biol. 2020, 432, 5287–5303. [Google Scholar] [CrossRef]
- Paulson, P.E.; Robinson, T.E. Amphetamine-induced time-dependent sensitization of dopamine neurotransmission in the dorsal and ventral striatum: A microdialysis study in behaving rats. Synapse 1995, 19, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Avelar, A.J.; Juliano, S.A.; Garris, P.A. Amphetamine augments vesicular dopamine release in the dorsal and ventral striatum through different mechanisms. J. Neurochem. 2013, 125, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Siciliano, C.A.; Calipari, E.S.; Jones, S.R. Amphetamine potency varies with dopamine uptake rate across striatal subregions. J. Neurochem. 2014, 131, 348–355. [Google Scholar] [CrossRef] [Green Version]
- Salahpour, A.; Ramsey, A.J.; Medvedev, I.O.; Kile, B.; Sotnikova, T.D.; Holmstrand, E.; Ghisi, V.; Nicholls, P.J.; Wong, L.; Murphy, K.; et al. Increased amphetamine-induced hyperactivity and reward in mice overexpressing the dopamine transporter. Proc. Natl. Acad. Sci. USA 2008, 105, 4405–4410. [Google Scholar] [CrossRef] [Green Version]
- Cagniard, B.; Sotnikova, T.D.; Gainetdinov, R.R.; Zhuang, X. The Dopamine Transporter Expression Level Differentially Affects Responses to Cocaine and Amphetamine. J. Neurogenet. 2014, 28, 112–121. [Google Scholar] [CrossRef]
- Shilling, P.D.; Kelsoe, J.R.; Segal, D.S. Dopamine transporter mRNA is up-regulated in the substantia nigra and the ventral tegmental area of amphetamine-sensitized rats. Neurosci. Lett. 1997, 236, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Dietz, D.M.; Tapocik, J.; Gaval-Cruz, M.; Kabbaj, M. Dopamine transporter, but not tyrosine hydroxylase, may be implicated in determining individual differences in behavioral sensitization to amphetamine. Physiol Behav. 2005, 86, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Underhill, S.M.; Hullihen, P.D.; Chen, J.; Fenollar-Ferre, C.; Rizzo, M.A.; Ingram, S.L.; Amara, S.G. Amphetamines signal through intracellular TAAR1 receptors coupled to Gα13 and GαS in discrete subcellular domains. Mol. Psychiatry. 2021, 26, 1208–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, A.M.; Domonkos, A.; Fernandez-Ruiz, A.; Freund, T.F.; Varga, V. Hippocampal Network Dynamics during Rearing Episodes. Cell Rep. 2018, 23, 1706–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czéh, B.; Michaelis, T.; Watanabe, T.; Frahm, J.; de Biurrun, G.; van Kampen, M.; Bartolomucci, A.; Fuchs, E. Stress-induced changes in cerebral metabolites, hippocampal volume, and cell proliferation are prevented by antidepressant treatment with tianeptine. Proc. Natl. Acad. Sci. USA 2001, 98, 12796–12801. [Google Scholar] [CrossRef] [Green Version]
- Sapolsky, R.M. Depression, antidepressants, and the shrinking hippocampus. Proc. Natl. Acad. Sci. USA 2001, 98, 12320–12322. [Google Scholar] [CrossRef] [Green Version]
- Kempermann, G.; Song, H.; Gage, F.H. Neurogenesis in the Adult Hippocampus. Cold Spring Harb. Perspect. Biol. 2015, 7, a018812. [Google Scholar] [CrossRef] [Green Version]
- Navailles, S.; De Deurwaerdère, P. Presynaptic control of serotonin on striatal dopamine function. Psychopharmacology 2011, 213, 213–242. [Google Scholar] [CrossRef]
- Fazekas, C.L.; Bellardie, M.; Török, B.; Sipos, E.; Tóth, B.; Baranyi, M.; Sperlágh, B.; Dobos-Kovács, M.; Chaillou, E.; Zelena, D. Pharmacogenetic excitation of the median raphe region affects social and depressive-like behavior and core body temperature in male mice. Life Sci. 2021, 286, 120037. [Google Scholar] [CrossRef]
- Admon, R.; Pizzagalli, D.A. Dysfunctional Reward Processing in Depression. Curr. Opin. Psychol. 2015, 4, 114–118. [Google Scholar] [CrossRef] [Green Version]
- Ng, T.H.; Alloy, L.B.; Smith, D.V. Meta-analysis of reward processing in major depressive disorder reveals distinct abnormalities within the reward circuit. Transl. Psychiatry 2019, 9, 293. [Google Scholar] [CrossRef]
- Kenny, P.J. Reward mechanisms in obesity: New insights and future directions. Neuron 2011, 69, 664–679. [Google Scholar] [CrossRef] [Green Version]
- Yadid, G.; Friedman, A. Dynamics of the dopaminergic system as a key component to the understanding of depression. Prog. Brain Res. 2008, 172, 265–286. [Google Scholar] [CrossRef]
- Van Galen, K.A.; ter Horst, K.W.; Serlie, M.J. Serotonin, food intake, obesity. Etiol. Pathophysiol. 2021, 22, e13210. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaare, M.; Jayaram, M.; Jagomäe, T.; Singh, K.; Kilk, K.; Mikheim, K.; Leevik, M.; Leidmaa, E.; Varul, J.; Nõmm, H.; et al. Depression-Associated Negr1 Gene-Deficiency Induces Alterations in the Monoaminergic Neurotransmission Enhancing Time-Dependent Sensitization to Amphetamine in Male Mice. Brain Sci. 2022, 12, 1696. https://doi.org/10.3390/brainsci12121696
Kaare M, Jayaram M, Jagomäe T, Singh K, Kilk K, Mikheim K, Leevik M, Leidmaa E, Varul J, Nõmm H, et al. Depression-Associated Negr1 Gene-Deficiency Induces Alterations in the Monoaminergic Neurotransmission Enhancing Time-Dependent Sensitization to Amphetamine in Male Mice. Brain Sciences. 2022; 12(12):1696. https://doi.org/10.3390/brainsci12121696
Chicago/Turabian StyleKaare, Maria, Mohan Jayaram, Toomas Jagomäe, Katyayani Singh, Kalle Kilk, Kaie Mikheim, Marko Leevik, Este Leidmaa, Jane Varul, Helis Nõmm, and et al. 2022. "Depression-Associated Negr1 Gene-Deficiency Induces Alterations in the Monoaminergic Neurotransmission Enhancing Time-Dependent Sensitization to Amphetamine in Male Mice" Brain Sciences 12, no. 12: 1696. https://doi.org/10.3390/brainsci12121696