
Intermodulation from Unisensory to Multisensory Perception: A Review

Abstract
:1. Introduction
2. Definitions of IMs
2.1. Time and Frequency Domains
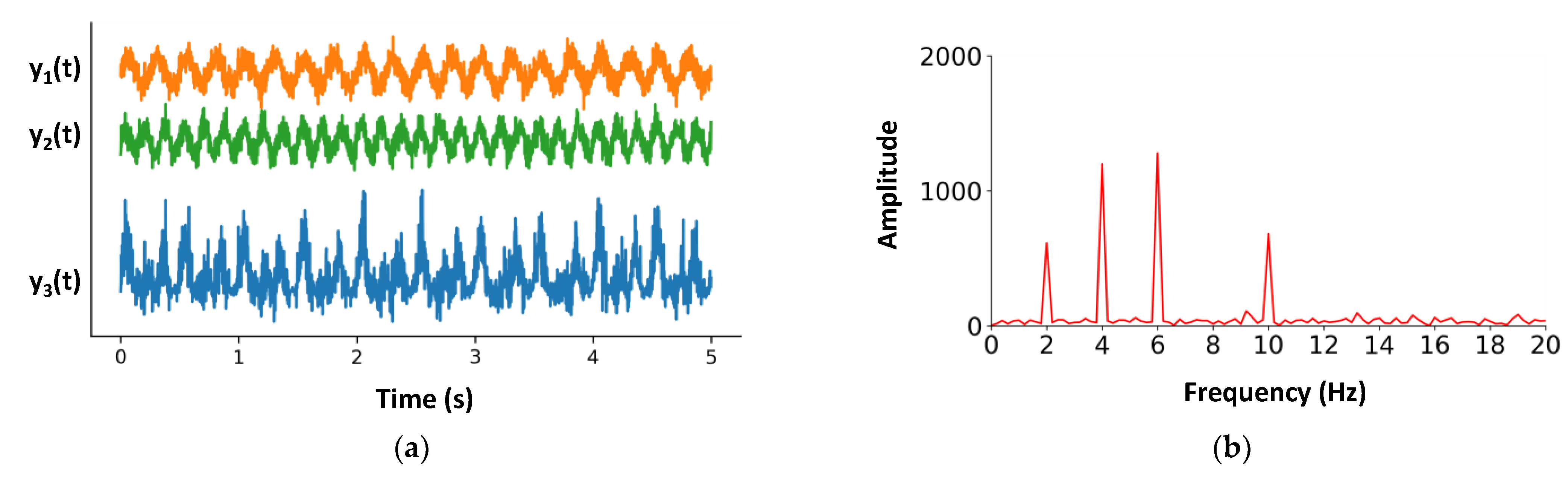
2.2. Example of IMs
2.3. The Order of IMs and Its Role in Cognition
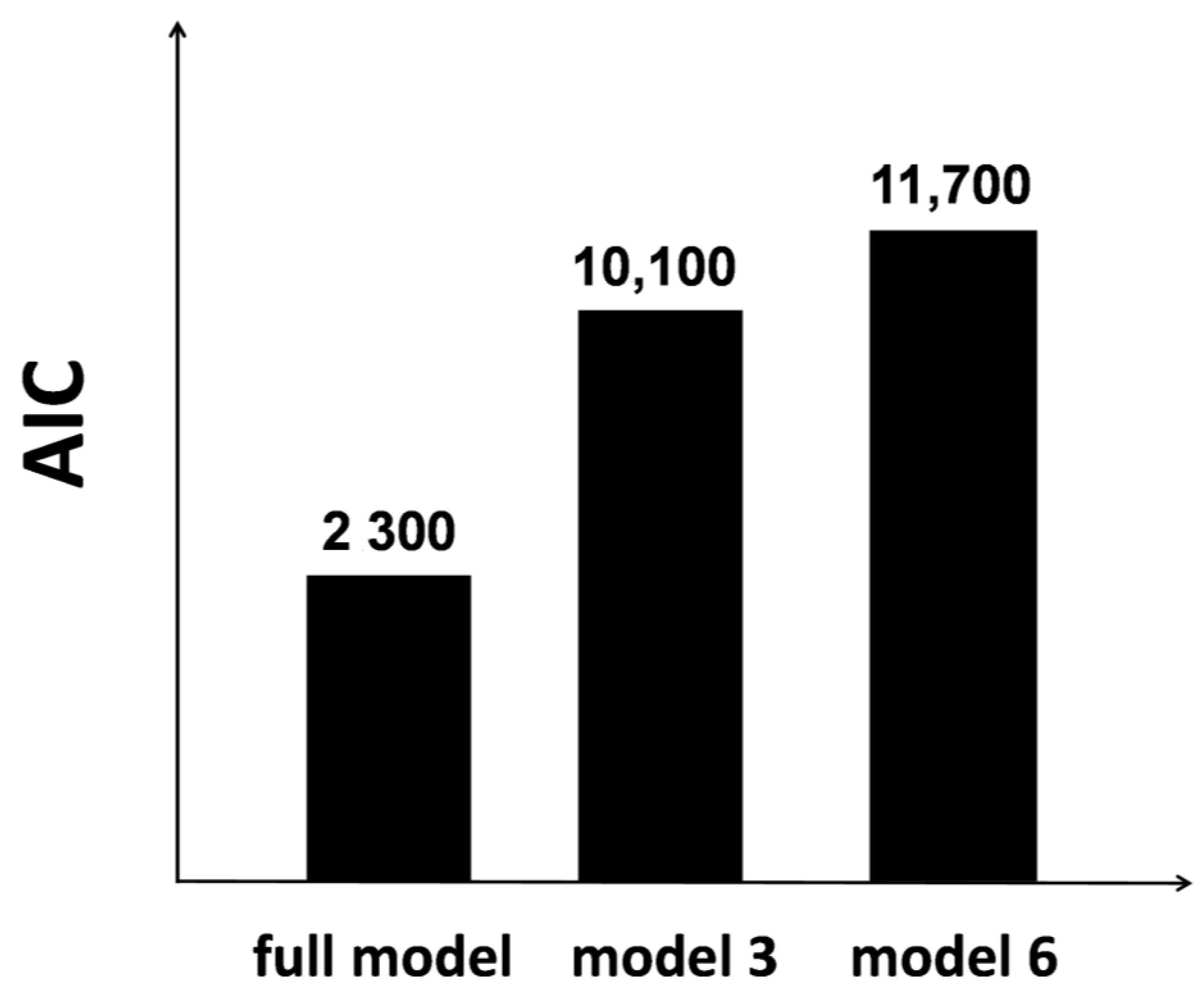
3. Modeling for Neural Interaction Processing
4. IMs for Multisensory Perception
4.1. IMs Studies of Multisensory Perception
4.2. IMs in the Spatial and Temporal Rules of Multisensory Perception
4.3. The Role of IMs in the Relationship between Multisensory Perception and Attention
4.4. The Role of IMs in the Diagnosis of Pathology by Brain–Computer Interface (BCI)
5. Limitations and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cardullo, F.; Sweet, B.; Hosman, R.; Coon, C. The Human Visual System and its Role in Motion Perception. In Proceedings of the AIAA Modeling and Simulation Technologies Conference, Portland, OR, USA, 8–11 August 2011; p. 6422. [Google Scholar]
- Delis, I.; Ince, R.A.; Sajda, P.; Wang, Q. Neural encoding of active multi-sensing enhances perceptual decision-making via a synergistic cross-modal interaction. J. Neurosci. 2022, 42, 2344–2355. [Google Scholar] [CrossRef] [PubMed]
- Flaten, E.; Marshall, S.A.; Dittrich, A.; Trainor, L.J. Evidence for top-down metre perception in infancy as shown by primed neural responses to an ambiguous rhythm. Eur. J. Neurosci. 2022, 55, 2003–2023. [Google Scholar] [CrossRef] [PubMed]
- Pang, C.Y.; Mueller, M.M. Competitive interactions in somatosensory cortex for concurrent vibrotactile stimulation between and within hands. Biol. Psychol. 2015, 110, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Nozaradan, S.; Peretz, I.; Mouraux, A. Steady-state evoked potentials as an index of multisensory temporal binding. NeuroImage 2012, 60, 21–28. [Google Scholar] [CrossRef]
- Porcu, E.; Keitel, C.; Müller, M.M. Concurrent visual and tactile steady-state evoked potentials index allocation of inter-modal attention: A frequency-tagging study. Neurosci. Lett. 2013, 556, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Boremanse, A.; Norcia, A.M.; Rossion, B. An objective signature for visual binding of face parts in the human brain. J. Vis. 2013, 13, 6. [Google Scholar] [CrossRef]
- Norcia, A.M.; Appelbaum, L.; Ales, J.M.; Cottereau, B.R.; Rossion, B. The steady-state visual evoked potential in vision research: A review. J. Vis. 2015, 15, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lithfous, S.; Rossion, B. Electrophysiological individual face adaptation effects with fast periodic visual stimulation resist long interruptions in adaptation. Biol. Psychol. 2018, 133, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Joan, K.T.; Rossion, B. An objective electrophysiological marker of face individualisation impairment in acquired prosopagnosia with fast periodic visual stimulation. Neuropsychologia 2016, 83, 100–113. [Google Scholar]
- Vergeer, M.; Kogo, N.; Nikolaev, A.R.; Alp, N.; Loozen, V.; Schraepen, B.; Wagemans, J. EEG frequency tagging reveals higher order intermodulation components as neural markers of learned holistic shape representations. Vis. Res. 2018, 152, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Mao, Y.; Ku, Y.; Chen, J. Holistic integration in the processing of Chinese characters as revealed by electroencephalography frequency tagging. Perception 2020, 49, 658–671. [Google Scholar] [CrossRef] [PubMed]
- Alp, N.; Kogo, N.; Van Belle, G.; Wagemans, J.; Rossion, B. Frequency tagging yields an objective neural signature of Gestalt formation. Brain Cogn. 2016, 104, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Gundlach, C.; Müller, M.M. Perception of illusory contours forms intermodulation responses of steady state visual evoked potentials as a neural signature of spatial integration. Biol. Psychol. 2013, 94, 55–60. [Google Scholar] [CrossRef]
- Aissani, C.; Cottereau, B.; Dumas, G.; Paradis, A.-L.; Lorenceau, J. Magnetoencephalographic signatures of visual form and motion binding. Brain Res. 2011, 1408, 27–40. [Google Scholar] [CrossRef]
- Ratliff, F.; Zemon, V. Some new methods for the analysis of lateral interactions that influence the visual evoked potential. Ann. New York Acad. Sci. 1982, 338, 113–124. [Google Scholar] [CrossRef]
- Zemon, V.; Ratliff, F. Visual evoked potentials: Evidence for lateral interactions. Proc. Natl. Acad. Sci. USA 1982, 79, 5723–5726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Rozell, C.J. Visual nonclassical receptive field effects emerge from sparse coding in a dynamical system. PLoS Comput. Biol. 2013, 9, e1003191. [Google Scholar] [CrossRef] [PubMed]
- Kouh, M.; Poggio, T. A canonical neural circuit for cortical nonlinear operations. Neural Comput. 2008, 20, 1427–1451. [Google Scholar] [CrossRef] [PubMed]
- Gordon, N.; Hohwy, J.; Davidson, M.J.; van Boxtel, J.J.; Tsuchiya, N. From intermodulation components to visual perception and cognition-a review. NeuroImage 2019, 199, 480–494. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, Y.; Li, R.; Chen, S.C.-Y.; Vickery, R.M.; Morley, J.W.; Tsuchiya, N. Steady state evoked potential (SSEP) responses in the primary and secondary somatosensory cortices of anesthetized cats: Nonlinearity characterized by harmonic and intermodulation frequencies. PLoS ONE 2021, 16, e0240147. [Google Scholar] [CrossRef] [PubMed]
- Heeger, D.J. Half-squaring in responses of cat striate cells. Vis. Neurosci. 1992, 9, 427–443. [Google Scholar] [CrossRef]
- Gabbiani, F.; Krapp, H.G.; Hatsopoulos, N.; Mo, C.-H.; Koch, C.; Laurent, G. Multiplication and stimulus invariance in a looming-sensitive neuron. J. Physiol. 2004, 98, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Peña, J.L.; Konishi, M. Robustness of Multiplicative Processes in Auditory Spatial Tuning. J. Neurosci. 2004, 24, 8907–8910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regan, M.P.; Regan, D. A frequency domain technique for characterizing nonlinearities in biological systems. J. Theor. Biol. 1988, 133, 293–317. [Google Scholar] [CrossRef]
- Campbell, D.; Farmer, D.; Crutchfield, J.; Jen, E. Experimental mathematics: The role of computation in nonlinear science. Commun. ACM 1985, 28, 374–384. [Google Scholar] [CrossRef]
- Wagemans, J.; Elder, J.H.; Kubovy, M.; Palmer, S.E.; Peterson, M.A.; Singh, M.; von der Heydt, R. A century of Gestalt psychology in visual perception: I. Perceptual grouping and figure–ground organization. Psychol. Bull. 2012, 138, 1172. [Google Scholar] [CrossRef] [Green Version]
- Yau, J.M.; DeAngelis, G.; Angelaki, D.E. Dissecting neural circuits for multisensory integration and crossmodal processing. Philos. Trans. R. Soc. B: Biol. Sci. 2015, 370, 20140203. [Google Scholar] [CrossRef] [PubMed]
- Joassin, F.; Pesenti, M.; Maurage, P.; Verreckt, E.; Bruyer, R.; Campanella, S. Cross-modal interactions between human faces and voices involved in person recognition. Cortex 2011, 47, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Helbig, H.B.; Ernst, M.O.; Ricciardi, E.; Pietrini, P.; Thielscher, A.; Mayer, K.M.; Schultz, J.; Noppeney, U. The neural mechanisms of reliability weighted integration of shape information from vision and touch. Neuroimage 2012, 60, 1063–1072. [Google Scholar] [CrossRef] [Green Version]
- Sarko, D.K.; Nidiffer, A.R.; Powers, A.R., III; Ghose, D.; Hillock-Dunn, A.; Fister, M.C.; Krueger, J.; Wallace, M.T. Spatial and temporal features of multisensory processes. In The Neural Bases of Multisensory Processes; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Costantini, M.; Robinson, J.; Migliorati, D.; Donno, B.; Ferri, F.; Northoff, G. Temporal limits on rubber hand illusion reflect individuals’ temporal resolution in multisensory perception. Cognition 2016, 157, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, N.P. The law of inverse effectiveness in neurons and behaviour: Multisensory integration versus normal variability. Neuropsychologia 2007, 45, 3340–3345. [Google Scholar] [CrossRef]
- Stein, B.E.; Stanford, T.R. Multisensory integration: Current issues from the perspective of the single neuron. Nat. Rev. Neurosci. 2008, 9, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Vroomen, J. Intersensory binding across space and time: A tutorial review. Atten. Percept. Psychophys. 2013, 75, 790–811. [Google Scholar] [CrossRef] [Green Version]
- De Keyser, R.; Legrain, V. Steady-stade evoked potentials to research multisensory integration. In Proceedings of the Neurocog, Leuven, Belgium, 28–29 November 2016. [Google Scholar]
- Jacoby, O.; Hall, S.E.; Mattingley, J.B. A crossmodal crossover: Opposite effects of visual and auditory perceptual load on steady-state evoked potentials to irrelevant visual stimuli. Neuroimage 2012, 61, 1050–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuś, R.; Spustek, T.; Zieleniewska, M.; Duszyk, A.; Rogowski, P.; Suffczyński, P. Integrated trimodal SSEP experimental setup for visual, auditory and tactile stimulation. J. Neural Eng. 2017, 14, 066002. [Google Scholar] [CrossRef]
- Colon, E.; Legrain, V.; Huang, G.; Mouraux, A. Frequency tagging of steady-state evoked potentials to explore the crossmodal links in spatial attention between vision and touch. Psychophysiology 2015, 52, 1498–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covic, A.; Keitel, C.; Porcu, E.; Schröger, E.; Müller, M.M. Audio-visual synchrony and spatial attention enhance processing of dynamic visual stimulation independently and in parallel: A frequency-tagging study. Neuroimage 2017, 161, 32–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talsma, D.; Senkowski, D.; Soto-Faraco, S.; Woldorff, M.G. The multifaceted interplay between attention and multisensory integration. Trends Cogn. Sci. 2010, 14, 400–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Burg, E.; Olivers, C.N.; Bronkhorst, A.W.; Theeuwes, J. Pip and pop: Nonspatial auditory signals improve spatial visual search. J. Exp. Psychol. Hum. Percept. Perform. 2008, 34, 1053. [Google Scholar] [CrossRef] [PubMed]
- Rutishauser, U.; Walther, D.; Koch, C.; Perona, P. Is bottom-up attention useful for object recognition? In Proceedings of the 2004 IEEE Computer Society Conference on Computer Vision and Pattern Recognition, Washington, DC, USA, 27 June–2 July 2004; Volume 2.
- Kreutzer, S.; Gereon, R. Fink, and Ralph Weidner. Attention modulates visual size adaptation. J. Vis. 2015, 15, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lien, M.C.; Ruthruff, E.; Goodin, Z.; Remington, R.W. Contingent attentional capture by top-down control settings: Converging evidence from event-related potentials. J. Exp. Psychol. Hum. Percept. Perform. 2008, 34, 509. [Google Scholar] [CrossRef] [PubMed]
- Benedek, M.; Bergner, S.; Könen, T.; Fink, A.; Neubauer, A.C. EEG alpha synchronization is related to top-down processing in convergent and divergent thinking. Neuropsychologia 2011, 49, 3505–3511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Long, J.; Yu, T.; Yu, Z.; Wang, C.; Zhang, H.; Guan, C. An EEG-based BCI system for 2-D cursor control by combining Mu/Beta rhythm and P300 potential. IEEE Trans. Biomed. Eng. 2010, 57, 2495–2505. [Google Scholar] [CrossRef] [PubMed]
- Farwell, L.; Donchin, E. Talking off the top of your head: Toward a mental prosthesis utilizing event-related brain potentials. Electroencephalogr. Clin. Neurophysiol. 1988, 70, 510–523. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Pan, J.; Long, J.; Yu, T.; Wang, F.; Yu, Z.; Wu, W. Multimodal BCIs: Target detection, multidimensional control, and awareness evaluation in patients with disorder of consciousness. Proc. IEEE 2015, 104, 332–352. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Description | Basis of Neural Processing | Output-IMs | Comment |
|---|---|---|---|---|
| 2 | A single input signal is processed linearly | Neurons transmit signals linearly | Fundamental frequency | Linear processing of signal cannot yield harmonics and IMs |
| Nonlinear half rectification of a single signal | Neuron firing rate is selectively inhibited to 0 | Fundamental frequency and 2nd order harmonics | Nonlinear processing of a single signal yields harmonics | |
| Two signals are first half rectified and then added | Nonlinear processing of multiple parallel signals | Fundamental frequency and 2nd order harmonics | Nonlinear processing of multiple signals without interaction terms cannot yield IMs | |
| Two signals are first half rectified and then multiplied | Nonlinear Sequence Processing of Multiple Serial Signals | Fundamental frequency, 2nd order harmonics, 2nd order IM and 3rd order IM | Nonlinear processing of multiple signals with interaction terms yields IMs | |
| Nonlinear half squaring of a single signal | Neuron firing rate is selectively inhibited to 0 | Fundamental frequency | ||
| Two signals processed to half squaring nonlinearity and then added | Nonlinear processing of multiple parallel signals | Fundamental frequency and 2nd order harmonics | ||
| Two signals processed to half squaring nonlinearity and multiplied | Nonlinear Sequence Processing of Multiple Serial Signals | Fundamental frequency, 2nd order harmonics and 2nd order IM | ||
| Nonlinear square wave of a single signal | Output of ON/OFF neurons | Fundamental frequency, 3rd order harmonics and 5th order harmonics | ||
| Two signals processed to squaring wave and then added | Nonlinear processing of multiple parallel signals | Fundamental frequency, 3rd order harmonics and 5th order harmonics | ||
| Two signals processed to squaring wave and then multiplied | Nonlinear Sequence Processing of Multiple Serial Signals | All IMs (low-order and high-order IM) | The interaction of square wave signals can generate many IMs | |
| Sum of the two signals as the input of logistic function | Sum of multiple neuron signals as input for logical selection | Fundamental frequency and 3rd order IM | ||
| Difference of the two signals as the input of logistic function | Difference of neuron signals as input for logical selection | Fundamental frequency and 3rd order IM |
| Model ID | Model Function |
|---|---|
| Model 1 | |
| Model 2 | |
| Model 3 | |
| Model 4 | |
| Model 5 | |
| Model 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, S.; Zhou, X.; Chen, L. Intermodulation from Unisensory to Multisensory Perception: A Review. Brain Sci. 2022, 12, 1617. https://doi.org/10.3390/brainsci12121617
Xu S, Zhou X, Chen L. Intermodulation from Unisensory to Multisensory Perception: A Review. Brain Sciences. 2022; 12(12):1617. https://doi.org/10.3390/brainsci12121617
Chicago/Turabian StyleXu, Shen, Xiaolin Zhou, and Lihan Chen. 2022. "Intermodulation from Unisensory to Multisensory Perception: A Review" Brain Sciences 12, no. 12: 1617. https://doi.org/10.3390/brainsci12121617