
Modeling Brain Functional Connectivity Patterns during an Isometric Arm Force Exertion Task at Different Levels of Perceived Exertion: A Graph Theoretical Approach †
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Methodological Pipeline
2.2. Participant Selection and Ethical Code
2.3. Apparatus
2.4. Experimental Design
2.5. EEG Data Acquisition
2.6. EEG Pipeline Analysis
2.7. Estimation of Functional Connectivity of EEG Cortical Sources
2.8. Graph Analysis and Computation of Measures
3. Statistical Analyses
3.1. Isometric Force
3.2. Rate of Perceived Physical Comfort
3.3. Source Functional Connectivity Estimations
3.4. Brain Network Analysis
4. Results
4.1. Anthropometric Characteristics
4.2. Isometric Arm Forces
4.3. Rate of Perceived Physical Comfort
4.4. Functional Connectivity
4.4.1. Functional Brain Patterns
4.4.2. Multiple Comparisons of Functional Connectivity
4.5. Brain Network Results
4.5.1. Topological Differences in the Global Network for Alpha Coherence Network
4.5.2. Topological Differences in the Global Network for Beta Coherence Network
4.5.3. Topological Differences in the Local Network
Betweenness Centrality Results
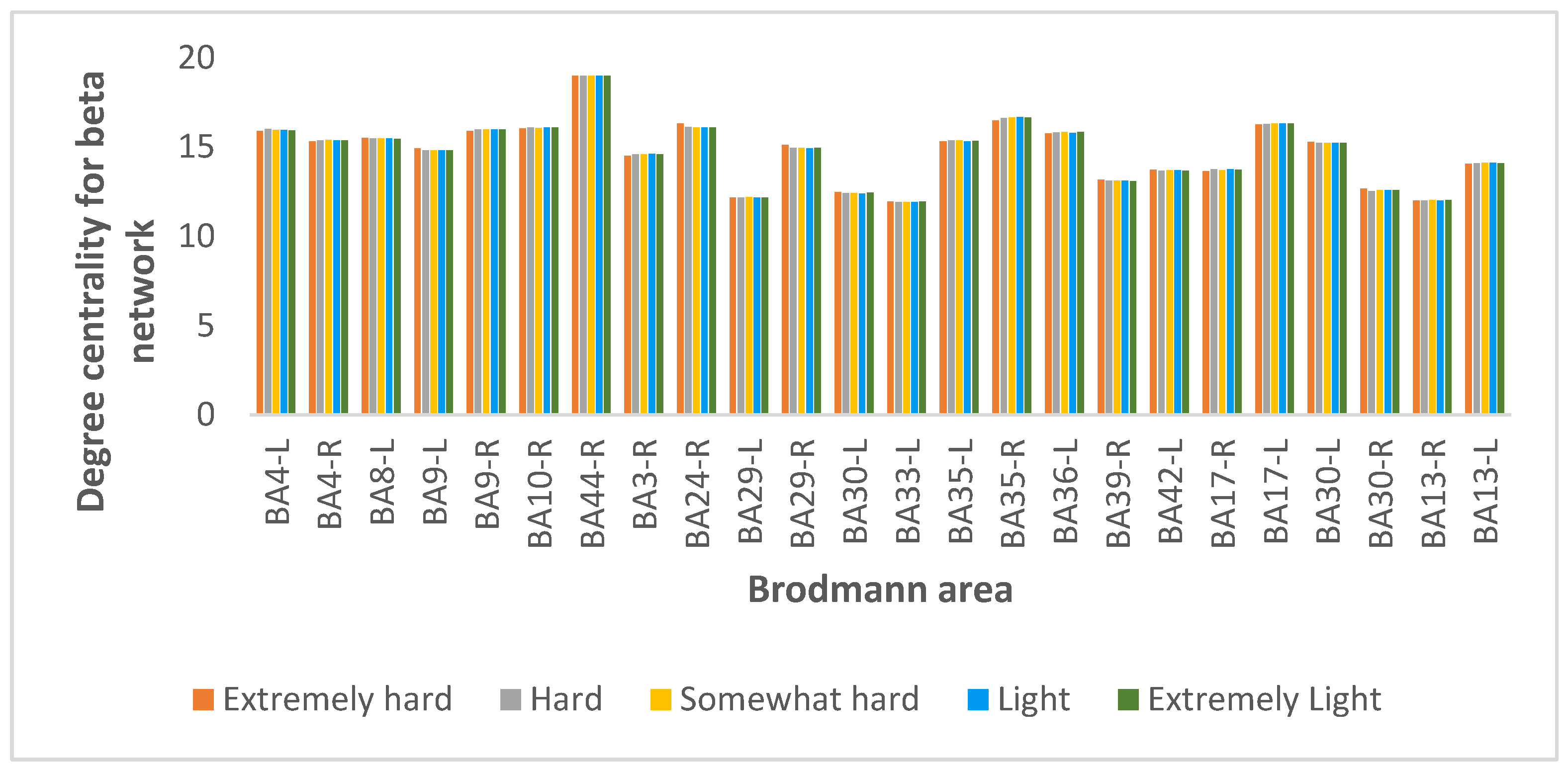
Degree Centrality Results
Nodal Efficiency
5. Discussion
5.1. Force Measures and RPPC Scores
5.2. Functional Brain Patterns
5.3. Brain Network
5.3.1. Global Measures
5.3.2. Local Measures
6. Limitations and Future Implications
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Morree, H.M.; Klein, C.; Marcora, S.M. Perception of Effort Reflects Central Motor Command during Movement Execution. Psychophysiology 2012, 49, 1242–1253. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O. The Human Connectome: A Complex Network. Ann. N. Y. Acad. Sci. 2011, 1224, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O.; Tononi, G.; Kötter, R. The Human Connectome: A Structural Description of the Human Brain. PLoS Comput. Biol. 2005, 1, 245–251. [Google Scholar] [CrossRef]
- Friston, K. Functional and Effective Connectivity: A Review. Brain Connect. 2011, 1, 13–36. [Google Scholar] [CrossRef] [PubMed]
- Baccalá, L.; Sameshima, K. Partial Directed Coherence: A New Concept in Neural Structure Determination. Biol. Cybern. 2001, 84, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Nunez, P.; Srinivasan, R.; Westdorp, A.; Wijesinghe, R.; Tucker, D.; Silberstein, R.; Cadusch, P. EEG Coherency I: Statistics, Reference Electrode, Volume Conduction, Laplacians, Cortical Imaging, and Interpretation at Multiple Scales. Electroencephalogr. Clin. Neurophysiol. 1997, 103, 499–515. [Google Scholar] [CrossRef]
- Lachaux, J.; Rodriguez, E.; Martinerie, J.; Varela, F. Measuring Phase Synchrony in Brain Signals. Hum. Brain Mapp. 1999, 8, 194–208. [Google Scholar] [CrossRef]
- Vinck, M.; Oostenveld, R.; Van Wingerden, M.; Battaglia, F.; Pennartz, C.M.A. An Improved Index of Phase-Synchronization for Electrophysiological Data in the Presence of Volume-Conduction, Noise and Sample-Size Bias. Neuroimage 2011, 55, 1548–1565. [Google Scholar] [CrossRef]
- García-Prieto, J.; Bajo, R.; Pereda, E. Efficient Computation of Functional Brain Networks: Toward Real-Time Functional Connectivity. Front. Neuroinform. 2017, 11, 1–18. [Google Scholar] [CrossRef]
- Pereda, E.; Quiroga, R.Q.; Bhattacharya, J. Nonlinear Multivariate Analysis of Neurophysiological Signals. Prog. Neurobiol. 2005, 77, 1–37. [Google Scholar] [CrossRef]
- Stam, C.; Dijk, B. Synchronization Likelihood: An Unbiased Measure of Generalized Synchronization in Multivariate Data Sets. Phys. D Nonlinear Phenom. 2002, 163, 236–251. [Google Scholar] [CrossRef]
- Wendling, F.; Ansari-Asl, K.; Bartolomei, F.; Senhadji, L. From EEG Signals to Brain Connectivity: A Model-Based Evaluation of Interdependence Measures. J. Neurosci. Methods 2009, 183, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, L.E.; Karwowski, W. A Graph Theory-Based Modeling of Functional Brain Connectivity Based on EEG: A Systematic Review in the Context of Neuroergonomics. IEEE Access 2020, 8, 155103–155135. [Google Scholar] [CrossRef]
- Jiang, Z.; Wang, X.; Kisiel-sajewicz, K.; Yan, J.H.; Yue, G.H. Strengthened Functional Connectivity in the Brain during Muscle Fatigue. Neuroimage 2012, 60, 728–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comani, S.; Fronso, S.D.; Castronovo, A.; Schmid, M.; Bortoli, L.; Conforto, S.; Robazza, C.; Bertollo, M. Attentional Focus and Functional Connectivity in Cycling: An EEG Case Study. In Proceedings of the XIII Mediterranean Conference on Medical and Biological Engineering and Computing, Seville, Spain, 25–28 September 2013; Springer: Cham, Switzerland, 2014; pp. 137–140. [Google Scholar]
- Wingfield, G.; Marino, F.; Skein, M. The Influence of Knowledge of Performance Endpoint on Pacing Strategies, Perception of Effort, and Neural Activity during 30-Km Cycling Time Trials. Physiol. Rep. 2018, 6, e13892. [Google Scholar] [CrossRef]
- Porter, S.; Silverberg, N.; Virji-Babul, N. Cortical Activity and Network Organization Underlying Physical and Cognitive Exertion in Active Young Adult Athletes: Implications for Concussion. J. Sci. Med. Sport 2019, 22, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.A.; Meng, L.F.; Ouyang, Y.; Chan, H.L.; Chang, Y.J.; Chen, S.W.; Liaw, J.W. Resistance-Induced Brain Activity Changes during Cycle Ergometer Exercises. BMC Sports Sci. Med. Rehabil. 2021, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Tamburro, G.; di Fronso, S.; Robazza, C.; Bertollo, M.; Comani, S. Modulation of Brain Functional Connectivity and Efficiency During an Endurance Cycling Task: A Source-Level EEG and Graph Theory Approach. Front. Hum. Neurosci. 2020, 14, 1–10. [Google Scholar] [CrossRef]
- Brümmer, V.; Schneider, S.; Abel, T.; Vogt, T.; Strüder, H.K. Brain Cortical Activity Is Influenced by Exercise Mode and Intensity. Med. Sci. Sport. Exerc. 2011, 43, 1863–1872. [Google Scholar] [CrossRef]
- Weng, T.B.; Pierce, G.L.; Darling, W.G.; Falk, D.; Magnotta, V.A.; Voss, M.W. The Acute Effects of Aerobic Exercise on the Functional Connectivity of Human Brain Networks. Brain Plast. 2017, 2, 171–190. [Google Scholar] [CrossRef]
- Pichardo-Rivas, K.A.; Gutiérrez, D. On the Functional Connectivity between Heart, Muscle, and Frontal Brain Cortex during Exercise Fatigability. Comput. Methods Biomech. Biomed. Engin. 2021, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Upadhyay, N.; Martin, J.A.; Rojas, S.; Strüder, H.K.; Boecker, H. Modulation of Distinct Intrinsic Resting State Brain Networks by Acute Exercise Bouts of Differing Intensity. Brain Plast. 2019, 5, 39–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büchel, D.; Sandbakk, Ø.; Baumeister, J. Exploring Intensity-Dependent Modulations in EEG Resting-State Network Efficiency Induced by Exercise. Eur. J. Appl. Physiol. 2021, 121, 2423–2435. [Google Scholar] [CrossRef] [PubMed]
- Diestel, R. Graph Theory; Springer: Berlin/Heidelberg, Germany, 1997. [Google Scholar]
- Danielle, S.B.; Bullmore, E.D. Small-World Brain Networks. Neuroscientist 2006, 12, 512–523. [Google Scholar] [CrossRef]
- Stam, C.; Reijneveld, J. Graph Theoretical Analysis of Complex Networks in the Brain. Nonlinear Biomed. Phys. 2007, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Bullmore, E.; Sporns, O. Complex Brain Networks: Graph Theoretical Analysis of Structural and Functional Systems. Nat. Rev. Neurosci. 2009, 10, 186. [Google Scholar] [CrossRef]
- Kaiser, M. A Tutorial in Connectome Analysis: Topological and Spatial Features of Brain Networks. Neuroimage 2011, 57, 892–907. [Google Scholar] [CrossRef] [Green Version]
- Boccaletti, S.; Latora, V.; Moreno, Y.; Chavez, M.; Hwang, D.U. Complex Network:Structure and Dynamics. Phys. Rep. 2006, 424, 175–308. [Google Scholar] [CrossRef]
- Churchill, N.W.; Madsen, K.; Mørup, M. The Functional Segregation and Integration Model: Mixture Model Representations of Consistent and Variable Group-Level Connectivity in FMRI. Neural Comput. 2016, 28, 2250–2290. [Google Scholar] [CrossRef]
- van den Heuvel, M.P.; Sporns, O. Network Hubs in the Human Brain. Trends Cogn. Sci. 2013, 17, 683–696. [Google Scholar] [CrossRef]
- Farahani, F.V.; Karwowski, W.; Lighthall, N.R. Application of Graph Theory for Identifying Connectivity Patterns in Human Brain Networks: A Systematic Review. Front. Neurosci. 2019, 13, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Rubinov, M.; Sporns, O. Complex Network Measures of Brain Connectivity: Uses and Interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O. Structure and Function of Complex Brain Networks. Dialogues Clin. Neurosci. 2013, 15, 247–262. [Google Scholar] [CrossRef]
- Farahani, F.V.; Fafrowicz, M.; Karwowski, W.; Douglas, P.K.; Domagalik, A.; Beldzik, E.; Oginska, H.; Winslow, B. Effects of Chronic Sleep Restriction on the Brain Functional Network, as Revealed by Graph Theory. Front. Neurosci. 2019, 13, 1087. [Google Scholar] [CrossRef]
- Dai, Z.; De Souza, J.; Lim, J.; Ho, P.M.; Chen, Y.; Li, J.; Thakor, N.; Bezerianos, A.; Sun, Y. Eeg Cortical Connectivity Analysis of Working Memory Reveals Topological Reorganization in Theta and Alpha Bands. Front. Hum. Neurosci. 2017, 11, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecchio, F.; Miraglia, F.; Maria Rossini, P. Connectome: Graph Theory Application in Functional Brain Network Architecture. Clin. Neurophysiol. Pract. 2017, 2, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.E.J. Modularity and Community Structure in Networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, M.E.J.; Girvan, M. Finding and Evaluating Community Structure in Networks. Phys. Rev. E 2004, 69, 026113. [Google Scholar] [CrossRef] [Green Version]
- Zuo, X.-N.; Ehmke, R.; Mennes, M.; Imperati, D.; Castellanos, F.X.; Sporns, O.; Milham, M.P. Network Centrality in the Human Functional Connectome. Cereb. Cortex 2012, 22, 1862–1875. [Google Scholar] [CrossRef]
- Watts, D.; Strogatz, S. Collective Dynamics of ‘Small-World’ Networks. Nature 1998, 393, 440–442. [Google Scholar] [CrossRef]
- Latora, V.; Marchiori, M. Efficient Behavior of Small-World Networks. Phys. Rev. Lett. 2001, 87, 198701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achard, S.; Bullmore, E. Efficiency and Cost of Economical Brain Functional Networks. PLoS Comput. Biol. 2007, 3, 0174–0183. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhao, M.; Yang, Y.; Gao, J.; Rao, N.; Lin, P. The Reorganization of Human Brain Networks Modulated by Driving Mental Fatigue. IEEE J. Biomed. Health Inform. 2017, 21, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Taya, F.; Sun, Y.; Desouza, J.; Thakor, N.V.; Bezerianos, A. Assessing Small-Worldness of Dynamic Functional Brain Connectivity during Complex Tasks. Proc. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. EMBS 2015, 2015, 2904–2907. [Google Scholar] [CrossRef]
- Davahli, M.R.; Karwowski, W.; Fiok, K.; Murata, A.; Sapkota, N.; Farahani, F.V.; Al-Juaid, A.; Marek, T.; Taiar, R. The COVID-19 Infection Diffusion in the US and Japan: A Graph-Theoretical Approach. Biology 2022, 11, 125. [Google Scholar] [CrossRef]
- Jafari, Z.; Kolb, B.E.; Mohajerani, M.H. Neural Oscillations and Brain Stimulation in Alzheimer’s Disease. Prog. Neurobiol. 2020, 194, 101878. [Google Scholar] [CrossRef]
- Hatlestad-Hall, C.; Bruña, R.; Syvertsen, M.R.; Erichsen, A.; Andersson, V.; Vecchio, F.; Miraglia, F.; Rossini, P.M.; Renvall, H.; Taubøll, E.; et al. Source-Level EEG and Graph Theory Reveal Widespread Functional Network Alterations in Focal Epilepsy. Clin. Neurophysiol. 2021, 132, 1663–1676. [Google Scholar] [CrossRef]
- Vecchio, F.; Miraglia, F.; Alú, F.; Orticoni, A.; Judica, E.; Cotelli, M.; Rossini, P.M. Contribution of Graph Theory Applied to EEG Data Analysis for Alzheimer’s Disease Versus Vascular Dementia Diagnosis. J. Alzheimer’s Dis. 2021, 82, 871–879. [Google Scholar] [CrossRef]
- Stam, C.; Haan, W.; Daffertshofer, A.; Jones, B.; Manshanden, I.; Walsum, A.; Cappellen, V.; Montez, T.; Verbunt, J.; Munck, J.; et al. Graph Theoretical Analysis of Magnetoencephalographic Functional Connectivity in Alzheimer ’ s Disease. Brain A J. Neurol. 2009, 132, 213–224. [Google Scholar] [CrossRef]
- Stam, C.J. Modern Network Science of Neurological Disorders. Nat. Rev. Neurosci. 2014, 15, 683–695. [Google Scholar] [CrossRef]
- Bassett, D.S.; Bullmore, E.T. Human Brain Networks in Health and Disease. Curr. Opin. Neurol. 2009, 22, 340–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffa, A.; Baumann, P.S.; Thiran, J.-P.; Hagmann, P. Structural Connectomics in Brain Diseases. Neuroimage 2013, 80, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Langer, N.; Pedroni, A.; Jäncke, L. The Problem of Thresholding in Small-World Network Analysis. PLoS ONE 2013, 8, e53199. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, M.P.; Stam, C.J.; Boersma, M.; Hulshoff Pol, H.E. Small-World and Scale-Free Organization of Voxel-Based Resting-State Functional Connectivity in the Human Brain. Neuroimage 2008, 43, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Mierlo, P.; Höller, Y.; Focke, N.; Vulliemoz, S. Network Perspectives on Epilepsy Using EEG MEG Source Connectivity. Front. Neurol. 2019, 10, 721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miraglia, F.; Vecchio, F.; Pappalettera, C.; Nucci, L.; Cotelli, M.; Judica, E.; Ferreri, F.; Rossini, P.M. Brain Connectivity and Graph Theory Analysis in Alzheimer’s and Parkinson’s Disease: The Contribution of Electrophysiological Techniques. Brain Sci. 2022, 12, 402. [Google Scholar] [CrossRef]
- Brunner, C.; Billinger, M.; Seeber, M.; Mullen, T.R.; Makeig, S.; Lansky, P.; Cohen, M.X.; Özkurt, T.E. Volume Conduction Influences Scalp-Based Connectivity Estimates. Front. Comput. Neurosci. 2016, 10, 121. [Google Scholar] [CrossRef] [Green Version]
- Astolfi, L.; Cincotti, F.; Mattia, D.; Marciani, M.G.; Baccala, L.A.; Fallani, F.D.V.; Salinari, S.; Ursino, M.; Zavaglia, M.; Ding, L.; et al. Comparison of Different Cortical Connectivity Estimators for High-Resolution EEG Recordings. Hum. Brain Mapp. 2007, 28, 143–157. [Google Scholar] [CrossRef] [Green Version]
- De Vico Fallani, F.; Richiardi, J.; Chavez, M.; Achard, S. Graph Analysis of Functional Brain Networks: Practical Issues in Translational Neuroscience. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130521. [Google Scholar] [CrossRef]
- Hjelmervik, H.; Hausmann, M.; Osnes, B.; Westerhausen, R.; Specht, K. Resting States Are Resting Traits–An FMRI Study of Sex Differences and Menstrual Cycle Effects in Resting State Cognitive Control Networks. PLoS ONE 2014, 9, e103492. [Google Scholar] [CrossRef]
- Zhang, J.; Dong, X.; Wang, L.; Zhao, L.; Weng, Z.; Zhang, T.; Sui, J.; Go, R.; Huang, Q.; Wu, J.; et al. Gender Differences in Global Functional Connectivity during Facial Emotion Processing: A Visual MMN Study. Front. Behav. Neurosci. 2018, 12, 220. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Karwowski, W.; Fafrowicz, M.; Hancock, P. Neuroergonomics Applications of Electroencephalography in Physical Activities: A Systematic Review. Front. Hum. Neurosci. 2019, 13, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, J.; Zhang, Y.; Tang, Y.; Yang, Y. Brain Differences Between Men and Women: Evidence From Deep Learning. Front. Neurosci. 2019, 13, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazlee, N.; Waiter, G.D.; Sandu, A. Age-associated Sex and Asymmetry Differentiation in Hemispheric and Lobar Cortical Ribbon Complexity across Adulthood: A UK Biobank Imaging Study. Hum. Brain Mapp. 2022. [Google Scholar] [CrossRef]
- Gao, Y.; Tang, Y.; Zhang, H.; Yang, Y.; Dong, T.; Jia, Q. Sex Differences of Cerebellum and Cerebrum: Evidence from Graph Convolutional Network. Interdiscip. Sci. Comput. Life Sci. 2022, 14, 532–544. [Google Scholar] [CrossRef]
- Ismail, L. Topological Changes in the Functional Brain Networks Induced by Isometric Force Exertions Using a Graph Theoretical Approach: An EEG-Based Neuroergonomics Study. Ph.D. Dissertation, University of Central Florida, Orlando, FL, USA, 2021. [Google Scholar]
- Pascual-Marqui, R.D. Discrete, 3D Distributed, Linear Imaging Methods of Electric Neuronal Activity. Part 1: Exact, Zero Error Localization. arXiv 2007, arXiv:0710.3341. [Google Scholar]
- Pascual-Marqui, R.D.; Michel, C.M.; Lehmann, D. Low Resolution Electromagnetic Tomography: A New Method for Localizing Electrical Activity in the Brain. Int. J. Psychophysiol. 1994, 18, 49–65. [Google Scholar] [CrossRef]
- van den Heuvel, M.P.; de Lange, S.C.; Zalesky, A.; Seguin, C.; Yeo, B.T.T.; Schmidt, R. Proportional Thresholding in Resting-State FMRI Functional Connectivity Networks and Consequences for Patient-Control Connectome Studies: Issues and Recommendations. Neuroimage 2017, 152, 437–449. [Google Scholar] [CrossRef]
- Jackson, A. Preemployment Physical Evaluation. Exerc. Sport Sci. Rev. 1994, 22, 53–90. [Google Scholar] [CrossRef]
- Chaffin, D.; Herrin, G.; Keyserling, W. An Updated Position. J. Occup. Environ. Med. 1978, 20, 403–408. [Google Scholar]
- Mital, A.; Kumar, S. Human Muscle Strength Definitions, Measurement, and Usage: Part I—Guidelines for the Practitioner. Int. J. Ind. Ergon. 1998, 22, 101–121. [Google Scholar] [CrossRef]
- Borg, G.A. Psychophysical Bases of Perceived Exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Naveenkumar, R.; Shankar, S.; Nithyaprakash, R.; Hariprasath, D.; Moulidaman, G.; Prakash, C. Evaluation of the Efficacy in Ergonomically Designed Handsaw to Minimize the Muscular Load on Arm and Shoulder Muscles among Carpenters. J. Inst. Eng. Ser. C 2022, 103, 1125–1132. [Google Scholar] [CrossRef]
- Karwowski, W.; Marras, W.S. The Occupational Ergonomics Handbook; Crc Press: Boca Raton, FL, USA, 1998. [Google Scholar]
- Karwowski, W. The Scale of Comfort in Physical Exertions. Unpublished Report. Fl, USA, 2018. [Google Scholar]
- Light, G.A.; Williams, L.E.; Minow, F.; Sprock, J.; Rissling, A.; Sharp, R.; Swerdlow, N.R.; Braff, D.L. Electroencephalography (EEG) and Event-Related Potentials (ERP’s) with Human Participants. In Current Protocols in Neuroscience; 2010; pp. 6–25. ISBN 0471142301. [Google Scholar]
- Heisz, J.J.; McIntosh, A.R. Applications of EEG Neuroimaging Data: Event-Related Potentials, Spectral Power, and Multiscale Entropy. J. Vis. Exp. 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Psychology Software Tools, Pittsburgh, PA, E.-P. 3. 0. R. Available online: https://support.pstnet.com/ (accessed on 7 July 2020).
- CGX Software. Available online: https://www.cgxsystems.com/software (accessed on 10 August 2020).
- Ortiz-Echeverri, C.J.; Salazar-Colores, S.; Rodríguez-Reséndiz, J.; Gómez-Loenzo, R.A. A New Approach for Motor Imagery Classification Based on Sorted Blind Source Separation, Continuous Wavelet Transform, and Convolutional Neural Network. Sensors 2019, 19, 4541. [Google Scholar] [CrossRef] [Green Version]
- Ibrahimi, D.; Mendiola-Santibanez, J.D.; Martinez, E.C.; Rodriguez, J.; Pacheco, I.T. Cortical Activity at Baseline and During Light Stimulation in Patients With Strabismus and Amblyopia. IEEE Access 2021, 9, 22430–22446. [Google Scholar] [CrossRef]
- Urigüen, J.; Zapirain, B. EEG Artifact Removal-State-of-the-Art and Guidelines. J. Neural Eng. 2015, 12, 031001. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An Open Source Toolbox for Analysis of Single-Trial EEG Dynamics Including Independent Component Analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Shamlo, N.; Mullen, T.; Kothe, C.; Su, K.; Robbins, K. The PREP Pipeline: Standardized Preprocessing for Large-Scale EEG Analysis. Front. Neuroinform. 2015, 9, 1–19. [Google Scholar] [CrossRef]
- Garcés Correa, A.; Laciar, E.; Patĩo, H.; Valentinuzzi, M. Artifact Removal from EEG Signals Using Adaptive Filters in Cascade. J. Phys. Conf. Ser. 2007, 90, 12081. [Google Scholar] [CrossRef]
- Christiano, L.; Fitzgerald, T. The Band Pass Filter. Int. Econ. Rev. 2003, 44, 435–465. [Google Scholar] [CrossRef]
- Maess, B.; Schröger, E.; Widmann, A. High-Pass Filters and Baseline Correction in M/EEG Analysis. Commentary on: “How Inappropriate High-Pass Filters Can Produce Artefacts and Incorrect Conclusions in ERP Studies of Language and Cognition”. J. Neurosci. Methods 2016, 266, 164–165. [Google Scholar] [CrossRef] [PubMed]
- Winkler, I.; Debener, S.; Muller, K.-R.; Tangermann, M. On the Influence of High-Pass Filtering on ICA-Based Artifact Reduction in EEG-ERP. In Proceedings of the 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Milan, Italy, 25–29 August 2015; IEEE; pp. 4101–4105. [Google Scholar]
- Chang, C.-Y.; Hsu, S.-H.; Pion-Tonachini, L.; Jung, T.-P. Evaluation of Artifact Subspace Reconstruction for Automatic EEG Artifact Removal. In Proceedings of the 2018 40th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Honolulu, HI, USA, 18–21 July 2018; IEEE; pp. 1242–1245. [Google Scholar]
- Mullen, T.; Kothe, C.; Chi, Y.M.; Ojeda, A.; Kerth, T.; Makeig, S.; Cauwenberghs, G.; Jung, T.-P. Real-Time Modeling and 3D Visualization of Source Dynamics and Connectivity Using Wearable EEG. In Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Osaka, Japan, 3–7 July 2013; pp. 2184–2187. [Google Scholar]
- Mullen, T.R.; Kothe, C.A.E.; Chi, Y.M.; Ojeda, A.; Kerth, T.; Makeig, S.; Jung, T.-P.; Cauwenberghs, G. Real-Time Neuroimaging and Cognitive Monitoring Using Wearable Dry EEG. IEEE Trans. Biomed. Eng. 2015, 62, 2553–2567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, J.; Kreutz-Delgado, K.; Makeig, S. AMICA: An Adaptive Mixture of Independent Component Analyzers with Shared Components. Swart. Cent. Comput. Neurosci. Univ. Calif. San Diego CA Tech. Rep. 2011, 1–15. [Google Scholar]
- Delorme, A.; Palmer, J.; Onton, J.; Oostenveld, R.; Makeig, S. Independent EEG Sources Are Dipolar. PLoS ONE 2012, 7, e30135. [Google Scholar] [CrossRef]
- Hsu, S.H.; Pion-Tonachini, L.; Palmer, J.; Miyakoshi, M.; Makeig, S.; Jung, T.P. Modeling Brain Dynamic State Changes with Adaptive Mixture Independent Component Analysis. Neuroimage 2018, 183, 47–61. [Google Scholar] [CrossRef]
- Pion-Tonachini, L.; Kreutz-Delgado, K.; Makeig, S. ICLabel: An Automated Electroencephalographic Independent Component Classifier, Dataset, and Website. Neuroimage 2019, 198, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Pion-Tonachini, L.; Kreutz-Delgado, K.; Makeig, S. The ICLabel Dataset of Electroencephalographic (EEG) Independent Component (IC) Features. Data Br. 2019, 25, 104101. [Google Scholar] [CrossRef]
- Oostendorp, T.F.; van Oosterom, A. Source Parameter Estimation in Inhomogeneous Volume Conductors of Arbitrary Shape. IEEE Trans. Biomed. Eng. 1989, 36, 382–391. [Google Scholar] [CrossRef]
- Oostenveld, R.; Oostendorp, T.F. Validating the Boundary Element Method for Forward and Inverse EEG Computations in the Presence of a Hole in the Skull. Hum. Brain Mapp. 2002, 17, 179–192. [Google Scholar] [CrossRef]
- Piazza, C.; Miyakoshi, M.; Akalin-Acar, Z.; Cantiani, C.; Reni, G.; Bianchi, A.M.; Makeig, S. An Automated Function for Identifying EEG Independent Components Representing Bilateral Source Activity. In IFMBE Proceedings; Springer: Cham, Switzerland, 2016; Volume 57, pp. 105–109. ISBN 9783319327013. [Google Scholar]
- Nguyen, P.; Li, X.; Hayashi, Y.; Yano, S.; Kondo, T. Estimation of Brain Dynamics Under Visuomotor Task Using Functional Connectivity Analysis Based on Graph Theory. In Proceedings of the 2019 IEEE 19th International Conference on Bioinformatics and Bioengineering (BIBE), Athens, Greece, 28–30 October 2019; pp. 577–582. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D.; Esslen, M.; Kochi, K.; Lehmann, D. Functional Imaging with Low-Resolution Brain Electromagnetic Tomography (LORETA): A Review. Methods Find Exp Clin Pharmacol. 2002, 24, 91–95. [Google Scholar] [PubMed]
- Pascual-Marqui, R.D. Standardized Low-Resolution Brain Electromagnetic Tomography (SLORETA): Technical Details. Methods Find Exp. Clin. Pharmacol. 2002, 24 (Suppl. D), 5–12. [Google Scholar]
- Fuchs, M.; Kastner, J.; Wagner, M.; Hawes, S.; Ebersole, J.S. A Standardized Boundary Element Method Volume Conductor Model Integral Equation Using Analytically Integrated Elements. Clin. Neurophysiol. 2002, 113, 702–712. [Google Scholar] [CrossRef]
- Mazziotta, J.; Toga, A.; Evans, A.; Fox, P.; Lancaster, J.; Zilles, K.; Woods, R.; Paus, T.; Simpson, G.; Pike, B.; et al. A Probabilistic Atlas and Reference System for the Human Brain: International Consortium for Brain Mapping (ICBM). Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2001, 356, 1293–1322. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, J.L.; Woldorff, M.G.; Parsons, L.M.; Liotti, M.; Freitas, C.S.; Rainey, L.; Kochunov, P.V.; Nickerson, D.; Mikiten, S.A.; Fox, P.T. Automated Talairach Atlas Labels for Functional Brain Mapping. Hum. Brain Mapp. 2000, 10, 120–131. [Google Scholar] [CrossRef]
- Canuet, L.; Tellado, I.; Couceiro, V.; Fraile, C.; Fernandez-Novoa, L.; Ishii, R.; Takeda, M.; Cacabelos, R. Resting-State Network Disruption and APOE Genotype in Alzheimer’s Disease: A Lagged Functional Connectivity Study. PLoS ONE 2012, 7, e46289. [Google Scholar] [CrossRef]
- Hata, M.; Kazui, H.; Tanaka, T.; Ishii, R.; Canuet, L.; Pascual-Marqui, R.D.; Aoki, Y.; Ikeda, S.; Kanemoto, H.; Yoshiyama, K.; et al. Functional Connectivity Assessed by Resting State EEG Correlates with Cognitive Decline of Alzheimer’s Disease-An ELORETA Study. Clin. Neurophysiol. 2016, 127, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Canuet, L.; Ishii, R.; Pascual-Marqui, R.D.; Iwase, M.; Kurimoto, R.; Aoki, Y.; Ikeda, S.; Takahashi, H.; Nakahachi, T.; Takeda, M. Resting-State EEG Source Localization and Functional Connectivity in Schizophrenia-like Psychosis of Epilepsy. PLoS ONE 2011, 6, e27863. [Google Scholar] [CrossRef] [Green Version]
- Di Lorenzo, G.; Daverio, A.; Ferrentino, F.; Santarnecchi, E.; Ciabattini, F.; Monaco, L.; Lisi, G.; Barone, Y.; Di Lorenzo, C.; Niolu, C.; et al. Altered Resting-State EEG Source Functional Connectivity in Schizophrenia: The Effect of Illness Duration. Front. Hum. Neurosci. 2015, 9, 234. [Google Scholar] [CrossRef] [Green Version]
- Shreekantiah Umesh, D.; Tikka, S.K.; Goyal, N.; Nizamie, S.H.; Sinha, V.K. Resting State Theta Band Source Distribution and Functional Connectivity in Remitted Schizophrenia. Neurosci. Lett. 2016, 630, 199–202. [Google Scholar] [CrossRef]
- Olbrich, S.; Olbrich, H.; Adamaszek, M.; Jahn, I.; Hegerl, U.; Stengler, K. Altered EEG Lagged Coherence during Rest in Obsessive-Compulsive Disorder. Clin. Neurophysiol. 2013, 124, 2421–2430. [Google Scholar] [CrossRef] [PubMed]
- Lanzone, J.; Imperatori, C.; Assenza, G.; Ricci, L.; Farina, B.; Di Lazzaro, V.; Tombini, M. Power Spectral Differences between Transient Epileptic and Global Amnesia: An Eloreta Quantitative Eeg Study. Brain Sci. 2020, 10, 613. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Marqui, R.D.; Biscay, R.J.; Valdes-Sosa, P.A.; Bosch-Bayard, J.; Riera-Diaz, J.J. Cortical Current Source Connectivity by Means of Partial Coherence Fields. arXiv 2011, arXiv:1108.0251. [Google Scholar] [CrossRef]
- Jatoi, M.A.; Kamel, N.; Malik, A.S.; Faye, I. EEG Based Brain Source Localization Comparison of sLORETA and eLORETA. Australas. Phys. Eng. Sci. Med. 2014, 37, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Nichols, T.; Holmes, A. Nonparametric Permutation Tests for Functional Neuroimaging. Hum. Brain Mapp. 2004, 15, 887–910. [Google Scholar] [CrossRef]
- Andrew, C.; Pfurtscheller, G. Lack of Bilateral Coherence of Post-Movement Central Beta Oscillations in the Human Electroencephalogram. Neurosci. Lett. 1999, 273, 89–92. [Google Scholar] [CrossRef]
- Nolte, G.; Bai, O.; Wheaton, L.; Mari, Z.; Vorbach, S.; Hallett, M. Identifying True Brain Interaction from EEG Data Using the Imaginary Part of Coherency. Clin. Neurophysiol. 2004, 115, 2292–2307. [Google Scholar] [CrossRef]
- Canteroa, J.L.; Atienzaa, M.; Salasa, R.M.; Gomez, C.M. Alpha EEG Coherence in Different Brain States: An Electrophysiological Index of the Arousal Level in Human Subjects. Neurosci. Lett. 1999, 271, 159–162. [Google Scholar] [CrossRef]
- Storti, S.; Formaggio, E.; Manganotti, P.; Menegaz, G. Brain Network Connectivity and Topological Analysis during Voluntary Arm Movements. Clin. EEG Neurosci. 2016, 47, 276–290. [Google Scholar] [CrossRef]
- Sauseng, P.; Klimesch, W.; Schabus, M.; Doppelmayr, M. Fronto-Parietal EEG Coherence in Theta and Upper Alpha Reflect Central Executive Functions of Working Memory. Int. J. Psychophysiol. 2005, 57, 97–103. [Google Scholar] [CrossRef]
- Bowyer, S.M. Coherence a Measure of the Brain Networks: Past and Present. Neuropsychiatr. Electrophysiol. 2016, 2, 1–12. [Google Scholar] [CrossRef]
- Wang, R.; Wang, J.; Yu, H.; Wei, X.; Yang, C.; Deng, B. Decreased Coherence and Functional Connectivity of Electroencephalograph in Alzheimer’s Disease. Chaos An Interdiscip. J. Nonlinear Sci. 2014, 24, 033136. [Google Scholar] [CrossRef] [PubMed]
- Adler, G.; Brassen, S.; Jajcevic, A. EEG Coherence in Alzheimer?S Dementia. J. Neural Transm. 2003, 110, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Walter, D. Coherence as a Measure of Relationship between EEG Records. Electroencephalogr. Clin. Neurophysiol. 1968, 24, 282. [Google Scholar]
- Pascual-Marqui, R.D.; Lehmann, D.; Koukkou, M.; Kochi, K.; Anderer, P.; Saletu, B.; Tanaka, H.; Hirata, K.; John, E.R.; Prichep, L.; et al. Assessing Interactions in the Brain with Exact Low-Resolution Electromagnetic Tomography. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2011, 369, 3768–3784. [Google Scholar] [CrossRef]
- Brier, M.R.; Thomas, J.B.; Fagan, A.M.; Hassenstab, J.; Holtzman, D.M.; Benzinger, T.L.; Morris, J.C.; Ances, B.M. Functional Connectivity and Graph Theory in Preclinical Alzheimer’s Disease. Neurobiol. Aging 2014, 35, 757–768. [Google Scholar] [CrossRef] [Green Version]
- Berlot, R.; Metzler-Baddeley, C.; Ikram, M.A.; Jones, D.K.; O’Sullivan, M.J. Global Efficiency of Structural Networks Mediates Cognitive Control in Mild Cognitive Impairment. Front. Aging Neurosci. 2016, 8, 292. [Google Scholar] [CrossRef] [Green Version]
- Farahani, F.V.; Karwowski, W.; D’Esposito, M.; Betzel, R.F.; Douglas, P.K.; Sobczak, A.M.; Bohaterewicz, B.; Marek, T.; Fafrowicz, M. Diurnal Variations of Resting-State FMRI Data: A Graph-Based Analysis. Neuroimage 2022, 256, 119246. [Google Scholar] [CrossRef]
- Freeman, L. A Set of Measures of Centrality Based on Betweenness. Sociometry 1977, 40, 35–41. [Google Scholar] [CrossRef]
- Wang, J.; Zuo, X.; He, Y. Graph-Based Network Analysis of Resting-State Functional MRI. Front. Syst. Neurosci. 2010, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sporns, O.; Honey, C.J.; Kötter, R. Identification and Classification of Hubs in Brain Networks. PLoS ONE 2007, 2, e1049. [Google Scholar] [CrossRef] [PubMed]
- Zinn, M.L.; Zinn, M.A.; Jason, L.A. Intrinsic Functional Hypoconnectivity in Core Neurocognitive Networks Suggests Central Nervous System Pathology in Patients with Myalgic Encephalomyelitis: A Pilot Study. Appl. Psychophysiol. Biofeedback 2016, 41, 283–300. [Google Scholar] [CrossRef] [PubMed]
- Ponomareva, N.; Andreeva, T.; Protasova, M.; Konovalov, R.; Krotenkova, M.; Malina, D.; Mitrofanov, A.; Fokin, V.; Illarioshkin, S.; Rogaev, E. Genetic Association Between Alzheimer’s Disease Risk Variant of the PICALM Gene and EEG Functional Connectivity in Non-Demented Adults. Front. Neurosci. 2020, 14, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, T.E.; Holmes, A.P. Nonparametric Permutation Tests for Functional Neuroimaging: A Primer with Examples. Hum. Brain Mapp. 2002, 15, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Sture Holm A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [CrossRef]
- Dallaway, N.; Leo, S.; Ring, C. How Am I Doing? Performance Feedback Mitigates the Effects of Mental Fatigue on Endurance Exercise Performance. Psychol. Sport Exerc. 2022, 62, 102210. [Google Scholar] [CrossRef]
- Wang, L.; Fang, M.; Hu, J. The Effect of Vibration Massage on Fatigue of the Upper Trapezius Muscle during Different Tasks. Symmetry 2022, 14, 2158. [Google Scholar] [CrossRef]
- Xia, M.; Wang, J.; He, Y. BrainNet Viewer: A Network Visualization Tool for Human Brain Connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef] [Green Version]
- Misselhorn, J.; Friese, U.; Engel, A.K. Frontal and Parietal Alpha Oscillations Reflect Attentional Modulation of Cross-Modal Matching. Sci. Rep. 2019, 9, 5030. [Google Scholar] [CrossRef] [Green Version]
- Johnston, J.; Rearick, M.; Slobounov, S. Movement-Related Cortical Potentials Associated with Progressive Muscle Fatigue in a Grasping Task. Clin. Neurophysiol. 2001, 112, 68–77. [Google Scholar] [CrossRef]
- Mackenzie, T.N.; Bailey, A.Z.; Mi, P.Y.; Tsang, P.; Jones, C.B.; Nelson, A.J. Human Area 5 Modulates Corticospinal Output during Movement Preparation. Neuroreport 2016, 27, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Vogt, B.A. Pain and Emotion Interactions in Subregions of the Cingulate Gyrus. Nat. Rev. Neurosci. 2005, 6, 533–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tops, M.; Boksem, M.A.S. A Potential Role of the Inferior Frontal Gyrus and Anterior Insula in Cognitive Control, Brain Rhythms, and Event-Related Potentials. Front. Psychol. 2011, 2, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, C.S. Anterior Cingulate Cortex, Error Detection, and the Online Monitoring of Performance. Science 1998, 280, 747–749. [Google Scholar] [CrossRef]
- Etkin, A.; Egner, T.; Kalisch, R. Emotional Processing in Anterior Cingulate and Medial Prefrontal Cortex. Trends Cogn. Sci. 2011, 15, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Vogt, B. Cingulate Neurobiology and Disease; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Erickson, K.I.; Leckie, R.L.; Weinstein, A.M. Physical Activity, Fitness, and Gray Matter Volume. Neurobiol. Aging 2014, 35, S20–S28. [Google Scholar] [CrossRef] [Green Version]
- van Praag, H.; Christie, B.R.; Sejnowski, T.J.; Gage, F.H. Running Enhances Neurogenesis, Learning, and Long-Term Potentiation in Mice. Proc. Natl. Acad. Sci. USA 1999, 96, 13427–13431. [Google Scholar] [CrossRef] [Green Version]
- Loprinzi, P.D.; Kane, C.J. Exercise and Cognitive Function. Mayo Clin. Proc. 2015, 90, 450–460. [Google Scholar] [CrossRef] [Green Version]
- Storti, S.F.; Galazzo, I.B.; Caliandro, P.; Iacovelli, C.; Menegaz, G. Connectivity Modulations Induced by Reaching&grasping Movements. In Proceedings of the 2018 26th European Signal Processing Conference (EUSIPCO), Rome, Italy, 3–7 September 2018; pp. 1392–1396. [Google Scholar] [CrossRef]
- Sciaraffa, N.; Borghini, G.; Aricò, P.; Di Flumeri, G.; Colosimo, A.; Bezerianos, A.; Thakor, N.V.; Babiloni, F. Brain Interaction during Cooperation: Evaluating Local Properties of Multiple-Brain Network. Brain Sci. 2017, 7, 90. [Google Scholar] [CrossRef]
- Kar, S.; Routray, A.; Nayak, B.P. Functional Network Changes Associated with Sleep Deprivation and Fatigue during Simulated Driving: Validation Using Blood Biomarkers. Clin. Neurophysiol. 2011, 122, 966–974. [Google Scholar] [CrossRef]
- Han, C.; Sun, X.; Yang, Y.; Che, Y.; Qin, Y. Brain Complex Network Characteristic Analysis of Fatigue during Simulated Driving Based on Electroencephalogram Signals. Entropy 2019, 21, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, B.L.; Dai, Z.; Thakor, N.; Bezerianos, A.; Sun, Y. Connectome Pattern Alterations with Increment of Mental Fatigue in One-Hour Driving Simulation. In Proceedings of the 2017 39th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Jeju, Korea, 11–15 July 2017; pp. 4355–4358. [Google Scholar] [CrossRef]
- Vecchio, F.; Miraglia, F.; Marra, C.; Quaranta, D.; Vita, M.G.; Bramanti, P.; Rossini, P.M. Human Brain Networks in Cognitive Decline: A Graph Theoretical Analysis of Cortical Connectivity from EEG Data. J. Alzheimer’s Dis. 2014, 41, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Breckel, T.P.K.; Thiel, C.M.; Bullmore, E.T.; Zalesky, A.; Patel, A.X.; Giessing, C. Long-Term Effects of Attentional Performance on Functional Brain Network Topology. PLoS ONE 2013, 8, e74125. [Google Scholar] [CrossRef]
- Klimesch, W. Alpha-Band Oscillations, Attention, and Controlled Access to Stored Information. Trends Cogn. Sci. 2012, 16, 606–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitzbichler, M.; Henson, R.; Smith, M.; Nathan, P.; Bullmore, E. Cognitive Effort Drives Workspace Configuration of Human Brain Functional Networks. J. Neurosci. 2011, 31, 8259–8270. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Luo, Y.; Zhang, Z.; Xu, Y.; Jiao, W.; Jiang, Y.; Huang, S.; Wang, C. Effects of Mental Fatigue on Small-World Brain Functional Network Organization. Neural Plast. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Ren, A.; Shang, J.; Lei, Q.; Zhang, Y.; Yin, Z.; Li, J.; von Deneen, K.M.; Huang, L. Combining Partial Directed Coherence and Graph Theory to Analyse Effective Brain Networks of Different Mental Tasks. Front. Hum. Neurosci. 2016, 10, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakkos, I.; Dimitrakopoulos, G.N.; Gao, L.; Zhang, Y.; Qi, P.; Matsopoulos, G.K.; Thakor, N.; Bezerianos, A.; Sun, Y. Mental Workload Drives Different Reorganizations of Functional Cortical Connectivity Between 2D and 3D Simulated Flight Experiments. IEEE Trans. Neural Syst. Rehabil. Eng. 2019, 27, 1704–1713. [Google Scholar] [CrossRef]
- Kim, J.; Cho, K.-H. Robustness Analysis of Network Modularity. IEEE Trans. Control Netw. Syst. 2016, 3, 348–357. [Google Scholar] [CrossRef] [Green Version]
- Stevens, A.A.; Tappon, S.C.; Garg, A.; Fair, D.A. Functional Brain Network Modularity Captures Inter- and Intra-Individual Variation in Working Memory Capacity. PLoS ONE 2012, 7, e30468. [Google Scholar] [CrossRef] [Green Version]
- Fallani, F.; Astolfi, L.; Cincotti, F.; Mattia, D.; Tocci, A.; Salinari, S.; Marciani, M.G.; Witte, H.; Colosimo, A.; Babiloni, F. Brain Network Analysis from High-Resolution EEG Recordings by the Application of Theoretical Graph Indexes. IEEE Trans. Neural Syst. Rehabil. Eng. 2008, 16, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.H.; Lin, P.; Hallett, M. Reorganization of Brain Functional Small-World Networks during Finger Movements. Hum. Brain Mapp. 2012, 33, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Kar, S.; Routray, A. Effect of Sleep Deprivation on Functional Connectivity of EEG Channels. IEEE Trans. Syst. Man, Cybern. Part ASystems Humans 2013, 43, 666–672. [Google Scholar] [CrossRef]
- Sengupta, A.; Datta, S.; Kar, S.; Routray, A. EEG Synchronization and Brain Networks: A Case Study in Fatigue. In Proceedings of the International Conference on Medical Imaging, m-Health and Emerging Communication Systems (MedCom), Greater Noida, India, 7–8 November 2014; pp. 278–282. [Google Scholar]
- Storti, S.; Formaggio, E.; Manganotti, P.; Menegaz, G. Cortical Network Modulation during Paced Arm Movements. In Proceedings of the 2015 23rd European Signal Processing Conference (EUSIPCO), Nice, France, 31 August–4 September 2015; pp. 2596–2600. [Google Scholar]
- Gao, Y.; Ren, L.; Li, R.; Zhang, Y. Electroencephalogram–Electromyography Coupling Analysis in Stroke Based on Symbolic Transfer Entropy. Front. Neurol. 2018, 8, 716. [Google Scholar] [CrossRef] [Green Version]
- Baig, M.Z.; Kavakli, M. Connectivity Analysis Using Functional Brain Networks to Evaluate Cognitive Activity during 3D Modelling. Brain Sci. 2019, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Chakarov, V.; Naranjo, J.R.; Schulte-Mönting, J.; Omlor, W.; Huethe, F.; Kristeva, R. Beta-Range EEG-EMG Coherence With Isometric Compensation for Increasing Modulated Low-Level Forces. J. Neurophysiol. 2009, 102, 1115–1120. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Ni, Y.; Sweeney, J.; He, B. Sparse Cortical Current Density Imaging in Motor Potentials Induced by Finger Movement. J. Neural Eng. 2011, 8, 036008. [Google Scholar] [CrossRef] [Green Version]
- Gwin, J.; Ferris, D. Beta- and Gamma-Range Human Lower Limb Corticomuscular Coherence. Front. Hum. Neurosci. 2012, 6, 258. [Google Scholar] [CrossRef] [Green Version]
- Negro, F.; Farina, D. Linear Transmission of Cortical Oscillations to the Neural Drive to Muscles Is Mediated by Common Projections to Populations of Motoneurons in Humans. J. Physiol. 2011, 589, 629–637. [Google Scholar] [CrossRef]
- Slobounov, S.; Hallett, M.; Newell, K.M. Perceived Effort in Force Production as Reflected in Motor-Related Cortical Potentials. Clin. Neurophysiol. 2004, 115, 2391–2402. [Google Scholar] [CrossRef]
- Bola, M.; Sabel, B. Dynamic Reorganization of Brain Functional Networks during Cognition. Neuroimage 2015, 114, 398–413. [Google Scholar] [CrossRef] [PubMed]
- Toppi, J.; De Vico Fallani, F.; Vecchiato, G.; Maglione, A.G.; Cincotti, F.; Mattia, D.; Salinari, S.; Babiloni, F.; Astolfi, L. How the Statistical Validation of Functional Connectivity Patterns Can Prevent Erroneous Definition of Small-World Properties of a Brain Connectivity Network. Comput. Math. Methods Med. 2012, 2012, 1–13. [Google Scholar] [CrossRef] [PubMed]
- van Wijk, B.C.M.; Stam, C.J.; Daffertshofer, A. Comparing Brain Networks of Different Size and Connectivity Density Using Graph Theory. PLoS ONE 2010, 5, e13701. [Google Scholar] [CrossRef]
- Hutchison, R.M.; Womelsdorf, T.; Allen, E.A.; Bandettini, P.A.; Calhoun, V.D.; Corbetta, M.; Della Penna, S.; Duyn, J.H.; Glover, G.H.; Gonzalez-Castillo, J.; et al. Dynamic Functional Connectivity: Promise, Issues, and Interpretations. Neuroimage 2013, 80, 360–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massullo, C.; Imperatori, C.; De Vico Fallani, F.; Ardito, R.B.; Adenzato, M.; Palmiero, L.; Carbone, G.A.; Farina, B. Decreased Brain Network Global Efficiency after Attachment Memories Retrieval in Individuals with Unresolved/Disorganized Attachment-Related State of Mind. Sci. Rep. 2022, 12, 4725. [Google Scholar] [CrossRef] [PubMed]
- Borghini, G.; Vecchiato, G.; Toppi, J.; Astolfi, L.; Maglione, A.; Isabella, R.; Caltagirone, C.; Kong, W.; Wei, D.; Zhou, Z.; et al. Assessment of Mental Fatigue during Car Driving by Using High Resolution EEG Activity and Neurophysiologic Indices. In Proceedings of the 2012 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, San Diego, CA, USA, 28 August–1 September 2012; Volume 70, pp. 6442–6445. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Mean | SD |
|---|---|---|
| Age (year) | 27.4 | 6.2 |
| Body Weight (kg) | 60.2 | 11 |
| Shoulder Height (cm) | 135.85 | 7.50 |
| Hip Height (cm) | 98.04 | 6.08 |
| Knee Height (cm) | 51.65 | 2.84 |
| Arm Height (cm) | 106.26 | 5.90 |
| Knuckle Height (cm) | 73.98 | 6.28 |
| Body Height (cm) | 163.00 | 7.260 |
| MVC (N) | 115.00 | 47.60 |
| Predefined Level of Physical Exertion | Isometric Arm Force (N) | ||||||
|---|---|---|---|---|---|---|---|
| Mean | SD | Range | % MVC | ||||
| Min | Max | Mean | Min | Max | |||
| Extremely hard | 67.35 | 35.25 | 2 | 18 | 49% | 3% | 99% |
| Hard | 41.83 | 18.9 | 3 | 28 | 30% | 4% | 60% |
| Somewhat hard | 34.58 | 16.7 | 3 | 66 | 25% | 2% | 48% |
| Light | 13.61 | 6.76 | 6 | 82 | 10% | 2% | 20% |
| Extremely light | 8.04 | 5.32 | 4 | 136 | 4% | 1% | 13% |
| Source | Df | Adj SS | Adj MS | F-Value | p-Value |
|---|---|---|---|---|---|
| Participant | 11 | 11,374 | 1034.0 | 4.05 | 0.00 |
| Exertion level | 4 | 27,108 | 6777.085 | 26.54 | 0.00 |
| Error | 44 | 11,236 | 255.4 | ||
| Total | 59 | 49,718 |
| Exertion Level | Mean | Grouping |
|---|---|---|
| Extremely hard | 67.4 | A |
| Hard | 41.83 | B |
| Somewhat hard | 34.57 | B |
| Light | 13.62 | C |
| Extremely light | 8.04 | C |
| Difference in Exertion Level | Difference in Mean | SE of Difference | Simultaneous 95% CI | T-Value | Adj p-Value |
|---|---|---|---|---|---|
| EL-EH | −59.31 | 6.52 | (−77.85, −40.76) | −9.09 | 0.000 |
| H-EH | −25.52 | 6.52 | (−44.06, −6.97) | −3.91 | 0.003 |
| L-EH | −53.73 | 6.52 | (−72.28, −35.19) | −8.24 | 0.000 |
| SWH-EH | −32.77 | 6.52 | (−51.32, −14.23) | −5.02 | 0.000 |
| H-EL | 33.79 | 6.52 | (15.25, 52.34) | 5.18 | 0.000 |
| L-EL | 5.58 | 6.52 | (−12.97, 24.12) | 0.85 | NS |
| SWH-EL | 26.53 | 6.52 | (7.99, 45.08) | 4.07 | 0.002 |
| L-H | −28.22 | 6.52 | (−46.76, −9.67) | −4.33 | 0.001 |
| SWH-H | −7.26 | 6.52 | (−25.80, 11.29) | −1.11 | NS |
| SWH-L | 20.96 | 6.52 | (2.41, 39.50) | 3.21 | 0.020 |
| Exertion Level | RPPC Mean | RPPC Standard Deviation |
|---|---|---|
| Extremely light | 8.80 | 1.48 |
| Light | 8.16 | 1.63 |
| Somewhat hard | 5.47 | 1.68 |
| Hard | 5 | 2.11 |
| Extremely hard | 3.9 | 2.30 |
| Source | Df | Adj SS | Adj MS | F-Value | p-Value |
|---|---|---|---|---|---|
| Participants | 11 | 61.71 | 5.60 | 7.56 | 0.000 |
| Exertion levels | 4 | 119.36 | 29.84 | 40.23 | 0.000 |
| Errors | 44 | 32.64 | 0.7417 | ||
| Total | 59 | 213.71 |
| Exertion Level | Mean | Grouping |
|---|---|---|
| Extremely hard | 4.583 | A |
| Hard | 5.375 | AB |
| Somewhat hard | 5.729 | B |
| Light | 7.750 | C |
| Extremely light | 8.23 | C |
| Difference in Exertion Level | Difference in Mean | SE of Difference | Simultaneous 95% CI | T-Value | Adj p-Value |
|---|---|---|---|---|---|
| EL-EH | 3.646 | 0.352 | (2.646, 4.645) | 10.37 | 0.000 |
| H-EH | 0.792 | 0.352 | (−0.208, 1.791) | 2.25 | 0.180 |
| L-EH | 3.167 | 0.352 | (2.167, 4.166) | 9.01 | 0.000 |
| SWH-EH | 1.146 | 0.352 | (0.146, 2.145) | 3.26 | 0.017 |
| H-EL | −2.854 | 0.352 | (−3.854, −1.855) | −8.12 | 0.000 |
| L-EL | −0.479 | 0.352 | (−1.479, 0.520) | −1.36 | NS |
| SWH-EL | −2.500 | 0.352 | (−3.499, −1.501) | −7.11 | 0.000 |
| L-H | 2.375 | 0.352 | (1.376, 3.374) | 6.75 | 0.000 |
| SWH-H | 0.354 | 0.352 | (−0.645, 1.354) | 1.01 | NS |
| SWH-L | −2.021 | 0.352 | (−3.020, −1.021) | −5.75 | 0.000 |
| Nodal Centrality | Frequency Band | Extremely Hard | Hard | Somewhat Hard | Light | Extremely Light |
|---|---|---|---|---|---|---|
| Betweenness centrality | Alpha | BA 10 | BA 11 | BA 11 | BA 11 | BA 11 |
| Betweenness centrality | Beta | BA 17 | BA 17 | BA 17 | BA 17 | BA 17 |
| Degree centrality | Alpha | BA 11 | BA 11 | BA 11 | BA 11 | BA 11 |
| Degree centrality | Beta | BA 44 | BA 44 | BA 44 | BA 44 | BA 44 |
| Nodal efficiency | Alpha | BA 11, 29 | BA 11, 29 | BA 11, 29 | BA 11, 29 | BA 11, 29 |
| Nodal efficiency | Beta | BA 44 | BA 44 | BA 44 | BA 44 | BA 44 |
| Reference | Task | Global Measures | Local Measures | Number of Participants | ROI | Frequency Bands |
|---|---|---|---|---|---|---|
| Our study | Isometric arm exertion | Eg, El, CC, PL, SW | Bw, D, Enodal | 12 females | 84 sources based | Alpha and beta |
| Fallani et al. [167] | Foot dorsal flexion | Eg, Eloc | N/A | 5 | 16 sources based | Alpha |
| Jin et al. [168] | Finger-tapping | Eg | Enodal | 12 males 3 females | Scalp based | Alpha and beta |
| Kar et al. [169] | Physical activity | CC, PL | D | 12 males | Scalp based | Alpha and theta |
| Sengupta et al. [170] | Physical exercise | CC, PL | N/A | 12 males | Scalp based | N/A |
| Storti et al. [171] | Arm movement | CC | Dc | 7 males 3 females | Scalp based | Alpha and beta |
| Storti et al. [153] | Reach and grasp | Weighted CC, weighted PL | N/A | 10 | Scalp based | Alpha, beta, delta, and theta |
| Huang et al. [163] | Play task | Eg, Eloc | D | 19 males | Scalp based | Beta and theta |
| Porter et al. [17] | Cycling | CC | N/A | 8 males 5 females | Scalp based | Theta |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ismail, L.; Karwowski, W.; Farahani, F.V.; Rahman, M.; Alhujailli, A.; Fernandez-Sumano, R.; Hancock, P.A. Modeling Brain Functional Connectivity Patterns during an Isometric Arm Force Exertion Task at Different Levels of Perceived Exertion: A Graph Theoretical Approach. Brain Sci. 2022, 12, 1575. https://doi.org/10.3390/brainsci12111575
Ismail L, Karwowski W, Farahani FV, Rahman M, Alhujailli A, Fernandez-Sumano R, Hancock PA. Modeling Brain Functional Connectivity Patterns during an Isometric Arm Force Exertion Task at Different Levels of Perceived Exertion: A Graph Theoretical Approach. Brain Sciences. 2022; 12(11):1575. https://doi.org/10.3390/brainsci12111575
Chicago/Turabian StyleIsmail, Lina, Waldemar Karwowski, Farzad V. Farahani, Mahjabeen Rahman, Ashraf Alhujailli, Raul Fernandez-Sumano, and P. A. Hancock. 2022. "Modeling Brain Functional Connectivity Patterns during an Isometric Arm Force Exertion Task at Different Levels of Perceived Exertion: A Graph Theoretical Approach" Brain Sciences 12, no. 11: 1575. https://doi.org/10.3390/brainsci12111575