
Central Serotonin/Noradrenaline Transporter Availability and Treatment Success in Patients with Obesity

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
1.1. Serotonin Transporter (5-HTT)
1.2. Noradrenaline Transporter (NAT)
1.3. Clinical Relevance of 5-HTT and NAT in Obesity
2. Materials and Methods
2.1. Participants
2.2. Study Design
2.3. BDI Questionnaire [30]; German Version [31]
2.4. BIS/BAS Questionnaire [32]; German Version [33]
2.5. MRI
2.6. PET
2.7. Imaging Data Processing and Analysis
2.8. Statistical Analysis
3. Results
3.1. Demographic Characteristics of Study Participants
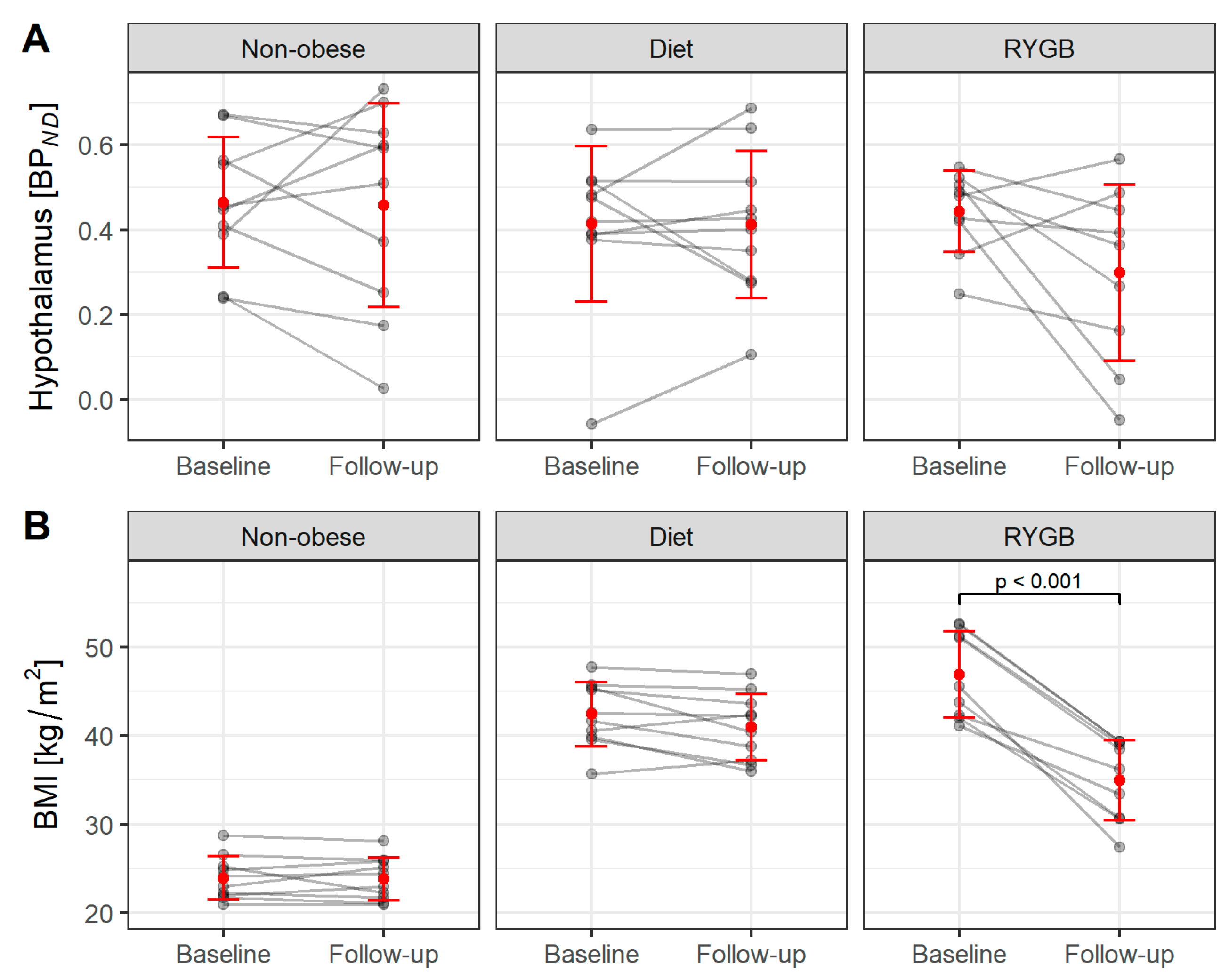
3.2. Changes in BMI and BPND during Diet and 6 Months after RYGB Surgery in the 5-HTT Cohort ([11C]DASB)
3.3. Changes in Body Weight and BMI during Diet and 6 Months after RYGB Surgery in the NAT Cohort ([11C]MRB)
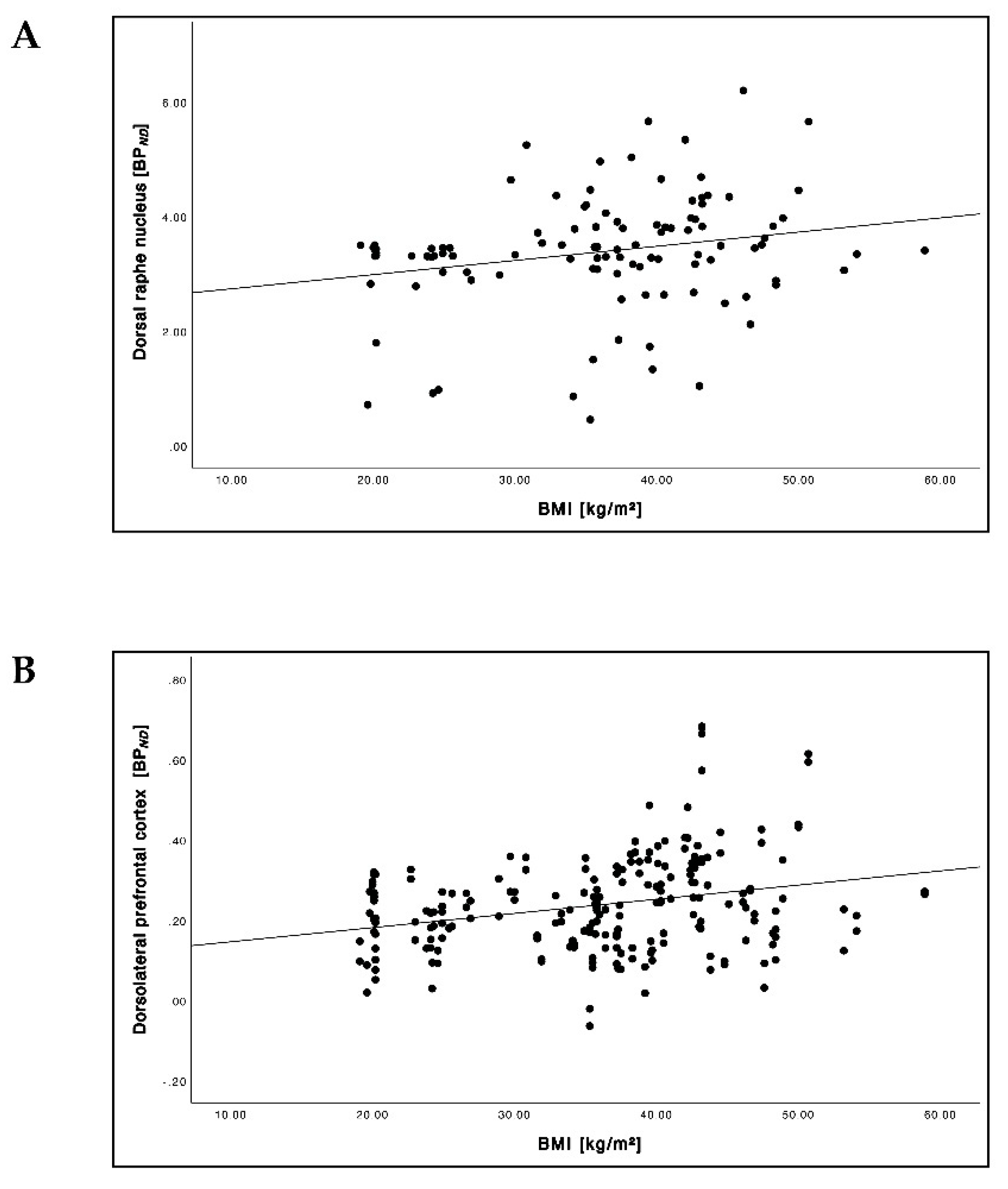
3.4. Relationship between the BMI and 5-HTT BPND
3.5. Relationship between the BMI and NAT BPND
3.6. Relationship between Pre-Interventional 5-HTT BPND and ΔBMI
3.7. Relationship between the BAS Reward Responsiveness Scores and 5-HTT BPND
4. Discussion
4.1. Group Characteristics
4.2. Relationship between BMI and 5-HTT Availability
4.3. Relationship between BMI and NAT Availability
4.4. Prediction of Treatment Success
4.5. 5-HTT Availability and Reward Responsiveness
4.6. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Van Galen, K.A.; ter Horst, K.W.; Booij, J.; la Fleur, S.E.; Serlie, M.J. The role of central dopamine and serotonin in human obesity: Lessons learned from molecular neuroimaging studies. Metabolism 2018, 85, 325–339. [Google Scholar] [CrossRef]
- Horstmann, A.; Fenske, W.K.; Hankir, M.K. Argument for a non-linear relationship between severity of human obesity and dopaminergic tone. Obes. Rev. 2015, 16, 821–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesse, S.; Rullmann, M.; Luthardt, J.; Winter, K.; Hankir, M.K.; Becker, G.A.; Zientek, F.; Reissig, G.; Regenthal, R.; Drabe, M.; et al. Central serotonin transporter availability in highly obese individuals compared with non-obese controls: A [11C]DASB positron emission tomography study. Eur. J. Nucl. Med. Mol. Imaging 2016, 43, 1096–1104. [Google Scholar] [CrossRef]
- Hesse, S.; van de Giessen, E.; Zientek, F.; Petroff, D.; Winter, K.; Dickson, J.C.; Tossici-Bolt, L.; Sera, T.; Asenbaum, S.; Darcourt, J.; et al. Association of central serotonin transporter availability and body mass index in healthy Europeans. Eur. Neuropsychopharmacol. 2014, 24, 1240–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesse, S.; Becker, G.A.; Rullmann, M.; Bresch, A.; Luthardt, J.; Hankir, M.K.; Zientek, F.; Reißig, G.; Patt, M.; Arelin, K.; et al. Central noradrenaline transporter availability in highly obese, non-depressed individuals. Eur. J. Nucl. Med. Mol. Imaging 2017, 44, 1056–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vettermann, F.J.; Rullmann, M.; Becker, G.A.; Luthardt, J.; Zientek, F.; Patt, M.; Meyer, P.M.; McLeod, A.; Brendel, M.; Blüher, M.; et al. Noradrenaline transporter availability on [11C]MRB PET predicts weight loss success in highly obese adults. Eur. J. Nucl. Med. Mol. Imaging 2018, 45, 1618–1625. [Google Scholar] [CrossRef] [Green Version]
- Soeder, J.M.; Luthardt, J.; Rullmann, M.; Becker, G.A.; Hankir, M.K.; Patt, M.; Meyer, P.M.; Schütz, T.; Ding, Y.; Hilbert, A.; et al. Central noradrenergic neurotransmission and weight loss 6 months after gastric bypass surgery in patients with severe obesity. Obes. Surg. 2021, 31, 4868–4876. [Google Scholar] [CrossRef]
- Li, C.S.; Potenza, M.N.; Lee, D.E.; Planeta, B.; Gallezot, J.D.; Labaree, D.; Henry, S.; Nabulsi, N.; Sinha, R.; Ding, Y.; et al. Decreased norepinephrine transporter availability in obesity: Positron Emission Tomography imaging with (S,S)-[11C]O-methylreboxetine. Neuroimage 2014, 86, 306–310. [Google Scholar] [CrossRef] [Green Version]
- Wurtman, R.J.; Wurtman, J.J. Brain serotonin, carbohydrate-craving, obesity and depression. Obes Res. 1995, 3 (Suppl. 4), 477S–480S. [Google Scholar] [CrossRef] [PubMed]
- Garfield, A.S.; Heisler, L.K. Pharmacological targeting of the serotonergic system for the treatment of obesity. J. Physiol. 2009, 587, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Dalley, J.W.; Roiser, J.P. Dopamine, serotonin and impulsivity. Neuroscience 2012, 215, 42–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.; Zhou, J.; Liu, Z. Reward processing by the dorsal raphe nucleus: 5-HT and beyond. Learn. Mem. 2015, 22, 452–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faulkner, P.; Deakin, J.F. The role of serotonin in reward, punishment and behavioural inhibition in humans: Insights from studies with acute tryptophan depletion. Neurosci. Biobehav. Rev. 2014, 46 Pt 3, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Bari, A.; Robbins, T.W. Inhibition and impulsivity: Behavioral and neural basis of response control. Prog. Neurobiol. 2013, 108, 44–79. [Google Scholar] [PubMed]
- Roberts, A.C. The importance of serotonin for orbitofrontal function. Biol. Psychiatry. 2011, 69, 1185–1191. [Google Scholar] [CrossRef]
- Fisher, P.M.; Meltzer, C.C.; Price, J.C.; Coleman, R.L.; Ziolko, S.K.; Becker, C.; Moses-Kolko, E.L.; Berga, S.L.; Hariri, A.R. Medial prefrontal cortex 5-HT2A density is correlated with amygdala reactivity, response habituation, and functional coupling. Cereb. Cortex 2009, 19, 2499–2507. [Google Scholar] [CrossRef]
- Haahr, M.E.; Rasmussen, P.M.; Madsen, K.; Marner, L.; Ratner, C.; Gillings, N.; Baaré, W.F.C.; Knudsen, G.M. Obesity is associated with high serotonin 4 receptor availability in the brain reward circuitry. Neuroimage 2012, 61, 884–888. [Google Scholar] [CrossRef]
- Huang, X.F.; Huang, X.; Han, M.; Chen, F.; Storlien, L.; Lawrence, A.J. 5-HT2A/2C receptor and 5-HT transporter densities in mice prone or resistant to chronic high-fat diet-induced obesity: A quantitative autoradiography study. Brain Res. 2004, 1018, 227–235. [Google Scholar] [CrossRef]
- Hummerich, R.; Reischl, G.; Ehrlichmann, W.; Machulla, H.J.; Heinz, A.; Schloss, P. DASB -in vitro binding characteristics on human recombinant monoamine transporters with regard to its potential as positron emission tomography (PET) tracer. J. Neurochem. 2004, 90, 1218–1226. [Google Scholar] [CrossRef]
- Huppelsberg, J.; Walter, K. Kurzlehrbuch Physiologie (3. Aufl); Thieme: Stuttgart, Germany, 2009. [Google Scholar]
- Leibowitz, S.F. Reciprocal hunger-regulating circuits involving alpha- and beta-adrenergic receptors located, respectively, in the ventromedial and lateral hypothalamus. Proc. Natl. Acad. Sci. USA 1970, 67, 1063–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Date, Y.; Shimbara, T.; Koda, S.; Toshinai, K.; Ida, T.; Murakami, N.; Miyazato, M.; Kokame, K.; Ishizuka, Y.; Ishida, Y.; et al. Peripheral ghrelin transmits orexigenic signals through the noradrenergic pathway from the hindbrain to the hypothalamus. Cell Metab. 2006, 4, 323–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J. Hunger, ghrelin and the gut. Brain Res. 2018, 1693 Pt B, 154–158. [Google Scholar] [CrossRef]
- Scheen, A.J. Sibutramine on cardiovascular outcome. Diabetes Care 2011, 34 (Suppl. 2), S114–S119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.H.; Chang, C.S.; Yang, Y.K.; Shen, L.H.; Yao, W.J. Comparison of brain serotonin transporter using [I-123]-ADAM between obese and non-obese young adults without an eating disorder. PLoS ONE 2017, 12, e0170886. [Google Scholar] [CrossRef] [Green Version]
- Haahr, M.E.; Hansen, D.L.; Fisher, P.M.; Svarer, C.; Stenbaek, D.S.; Madsen, K.; Madsen, J.; Holst, J.J.; Baaré, W.F.C.; Hojgaard, L.; et al. Central 5-HT neurotransmission modulates weight loss following gastric bypass surgery in obese individuals. J. Neurosci. 2015, 35, 5884–5889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.; Jang, M.; Lee, Y.; Ha, J.; Park, S. Amino acid metabolites and slow weight loss in the early postoperative period after sleeve gastrectomy. J. Clin. Med. 2020, 9, 2348. [Google Scholar] [CrossRef]
- Falkai, P.; Wittchen, H.U. (Eds.) Diagnostisches und Statistisches Manual Psychischer Störungen DSM-5; Hogrefe: Göttingen, Germany, 2015. [Google Scholar]
- Beck, A.T.; Ward, C.H.; Mendelson, J.; Mock, J.; Erbaugh, J. An inventory for measuring depression. Arch. Gen. Psychiatry 1961, 4, 564–571. [Google Scholar] [CrossRef] [Green Version]
- Hautzinger, M. Das Beck-Depressionsinventar (BDI) in der Klinik. Nervenarzt 1991, 62, 689–696. [Google Scholar]
- Carver, C.S.; White, T.L. Behavioral inhibition, behavioral activation, and affective responses to impending reward and punishment: The BIS/BAS Scales. J. Personal. Soc. Psychol. 1994, 67, 319–333. [Google Scholar] [CrossRef]
- Strobel, A.; Beauduce, A.; Debener, S.; Brocke, B. Eine deutschsprachige Version des BIS/BAS-Fragebogens von Carver und White. Z. Für Differ. Diagn. Psychol. 2001, 22, 216–227. [Google Scholar] [CrossRef]
- Drabe, M.; Rullmann, M.; Luthardt, J.; Boettcher, Y.; Regenthal, R.; Plötz, T.; Becker, G.A.; Patt, M.; Schinke, C.; Bergh, F.T.; et al. Serotonin transporter gene promoter methylation status correlates with in vivo prefrontal 5-HTT availability and reward function in human obesity. Transl. Psychiatry 2017, 7, e1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, A.A.; Ginovart, N.; Schmidt, M.; Meyer, J.H.; Threlkeld, P.G.; Houle, S. Novel radiotracers for imaging the serotonin transporter by positron emission tomography: Synthesis, radiosynthesis, and in vitro and ex vivo evaluation of 11C-labeled 2-(phenylthio)araalkylamines. J. Med. Chem. 2000, 43, 3103–3110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, K.S.; Ding, Y.S. Synthesis, enantiomeric resolution, and selective C-11 methylation of a highly selective radioligand for imaging the norepinephrine transporter with positron emission tomography. Chirality 2004, 16, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Ichise, M.; Liow, J.S.; Lu, J.Q.; Takano, A.; Model, K.; Toyama, H.; Suhara, T.; Suzuki, K.; Innis, R.B.; Carson, R.E. Linearized reference tissue parametric imaging methods: Application to [11C]DASB positron emission tomography studies of the serotonin transporter in human brain. J. Cereb. Blood Flow Metab. 2003, 23, 1096–1112. [Google Scholar] [CrossRef] [Green Version]
- Bertoldo, A.; Cobelli, C. Physiological modelling of positron emission tomography images. In Modelling Methodology for Physiology and Medicine; Carson, E., Cobelli, C., Eds.; Elsevier: London, UK, 2014; pp. 417–448. [Google Scholar]
- Innis, R.B.; Cunningham, V.J.; Delforge, J.; Fujita, M.; Gjedde, A.; Gunn, R.N.; Holden, J.; Houle, S.; Huang, S.; Ichise, M.; et al. Consensus nomenclature for in vivo imaging of reversibly binding radioligands. J. Cereb. Blood Flow Metab. 2007, 27, 1533–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milaneschi, Y.; Simmons, W.K.; van Rossum, E.F.C.; Penninx, B.W. Depression and obesity: Evidence of shared biological mechanisms. Mol. Psychiatry 2019, 24, 18–33. [Google Scholar] [CrossRef]
- Northoff, G.; Wiebking, C.; Feinberg, T.; Panksepp, J. The ‘resting-state hypothesis’ of major depressive disorder-a translational subcortical-cortical framework for a system disorder. Neurosci. Biobehav. Rev. 2011, 35, 29–1945. [Google Scholar] [CrossRef]
- Melasch, J.; Rullmann, M.; Hilbert, A.; Luthardt, J.; Becker, G.A.; Patt, M.; Villringer, A.; Arelin, K.; Meyer, P.M.; Lobsien, D.; et al. The central nervous norepinephrine network links a diminished sense of emotional well-being to an increased body weight. Int. J. Obes. 2016, 40, 779–787. [Google Scholar] [CrossRef] [Green Version]
- Dayan, D.; Kuriansky, J.; Abu-Abeid, S. Weight Regain Following Roux-en-Y Gastric Bypass: Etiology and Surgical Treatment. Isr. Med. Assoc. J. 2019, 21, 823–828. [Google Scholar]
- Spies, M.; Knudsen, G.M.; Lanzenberger, R.; Kasper, S. The serotonin transporter in psychiatric disorders: Insights from PET imaging. Lancet Psychiatry 2015, 2, 743–755. [Google Scholar] [CrossRef]
- Li, Y.; Zhong, W.; Wang, D.; Feng, Q.; Liu, Z.; Zhou, J.; Jia, C.; Hu, F.; Zeng, J.; Guo, Q.; et al. Serotonin neurons in the dorsal raphe nucleus encode reward signals. Nat. Commun. 2016, 7, 10503. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Obesity | Diet | RYGB | p # | ||||
|---|---|---|---|---|---|---|---|
| BS | FU | BS | FU | BS | FU | ||
| n | 23 (13 females) | 38 (25 females) | 21 (12 females) | 0.735 | |||
| Age (years) | 35.2 ± 8.5 | 35.5 ± 8.4 | 36.4 ± 9.8 | 36.9 ± 9.9 | 49.8 ± 10.7 | 50.4 ± 10.8 | <0.001 |
| BMI (kg/m2) | 23.1 ± 2.6 | 23.1 ± 2.5 | 41.8 ± 4.6 | 40.5 ± 5.5 | 44.8 ± 5.0 | 33.7 ± 4.8 | <0.001 |
| p * | 0.969 | 0.010 | <0.001 | ||||
| BDI | 3.4 ± 4.9 | 2.4 ± 3.6 | 9.7 ± 8.0 | 8.0 ± 6.5 | 8.6 ± 5.6 | 7.4 ± 4.6 | 0.002 |
| p * | 0.251 | 0.197 | 0.255 | ||||
| Non-obesity | Diet | RYGB | p # | |||||
|---|---|---|---|---|---|---|---|---|
| BS | FU | BS | FU | BS | FU | |||
| n | 23 (13 females) | 38 (25 females) | 21 (12 females) | |||||
| BPND | DRN | 3.1 ± 0.7 | 3.0 ± 0.8 | 3.5 ± 0.9 | 3.6 ± 1.1 | 3.3 ± 1.2 | 3.4 ± 1.3 | 0.272 |
| p * | 0.679 | 0.478 | 0.820 | |||||
| dlPFC | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 | <0.001 | |
| p * | 0.024 | 0.820 | 0.711 | |||||
| OFC | 0.4 ± 0.1 | 0.4 ± 0.1 | 0.5 ± 0.2 | 0.5 ± 0.2 | 0.3 ± 0.2 | 0.4 ± 0.2 | <0.001 | |
| p * | 0.739 | 0.482 | 0.186 | |||||
| Non-obesity | Diet | RYGB | p # | |||||
|---|---|---|---|---|---|---|---|---|
| BS | FU | BS | FU | BS | FU | |||
| n | 10 (4 females) | 10 (4 females) | 9 (7 females) | |||||
| BPND | Hypothalamus | 0.5 ± 0.2 | 0.5 ± 0.3 | 0.4 ± 0.2 | 0.4 ± 0.2 | 0.4 ± 0.2 | 0.3 ± 0.2 | 0.452 |
| p * | 0.910 | 0.963 | 0.820 | |||||
| Non-Obesity | Diet | RYGB | p # | ||||
|---|---|---|---|---|---|---|---|
| BS | FU | BS | FU | BS | FU | ||
| n | 12 (9 females) | 28 (21 females) | 11 (5 females) | ||||
| BAS Reward | 17.2 ± 1.9 | 17.9 ± 1.8 | 16.3 ± 2.3 | 16.2 ± 2.1 | 15.0 ± 2.9 | 14.3 ± 1.9 | 0.091 |
| p * | 0.082 | 0.757 | 0.863 | ||||
| Diet | RYGB | |||||||
|---|---|---|---|---|---|---|---|---|
| Pearson’s r | p | Pearson’s r | p | |||||
| BS | FU | BS | FU | BS | FU | BS | FU | |
| dlPFC | −0.60 | −0.61 | <0.001 | <0.001 | 0.48 | 0.17 | 0.025 | 0.442 |
| OFC | −0.44 | −0.46 | <0.001 | <0.001 | 0.67 | 0.59 | <0.001 | 0.004 |
| ACC | −0.61 | −0.48 | <0.001 | <0.001 | 0.43 | 0.39 | 0.045 | 0.074 |
| Insula | −0.49 | −0.51 | <0.001 | <0.001 | 0.53 | 0.01 | 0.011 | 0.956 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Griebsch, N.-I.; Kern, J.; Hansen, J.; Rullmann, M.; Luthardt, J.; Helfmeyer, S.; Dekorsy, F.J.; Soeder, M.; Hankir, M.K.; Zientek, F.; et al. Central Serotonin/Noradrenaline Transporter Availability and Treatment Success in Patients with Obesity. Brain Sci. 2022, 12, 1437. https://doi.org/10.3390/brainsci12111437
Griebsch N-I, Kern J, Hansen J, Rullmann M, Luthardt J, Helfmeyer S, Dekorsy FJ, Soeder M, Hankir MK, Zientek F, et al. Central Serotonin/Noradrenaline Transporter Availability and Treatment Success in Patients with Obesity. Brain Sciences. 2022; 12(11):1437. https://doi.org/10.3390/brainsci12111437
Chicago/Turabian StyleGriebsch, Nora-Isabell, Johanna Kern, Jonas Hansen, Michael Rullmann, Julia Luthardt, Stephanie Helfmeyer, Franziska J. Dekorsy, Marvin Soeder, Mohammed K. Hankir, Franziska Zientek, and et al. 2022. "Central Serotonin/Noradrenaline Transporter Availability and Treatment Success in Patients with Obesity" Brain Sciences 12, no. 11: 1437. https://doi.org/10.3390/brainsci12111437