
Short-Term Habituation of Auditory N1 in Spoken Word-Forms Is Modulated by Phonological Information

Abstract
:1. Introduction
2. Literature Review
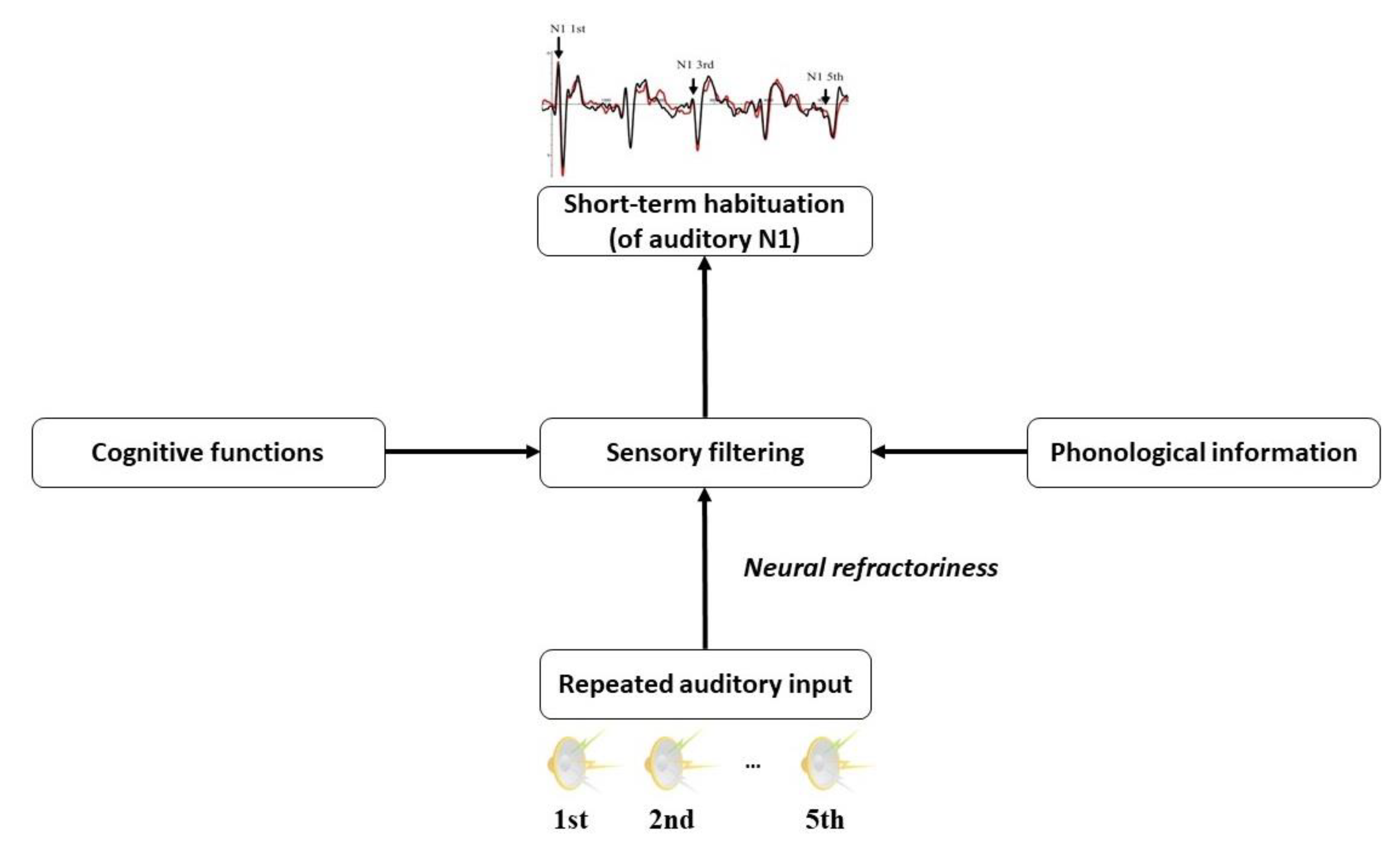
2.1. Short-Term Auditory Habituation
2.2. N1 Habituation Modulated by Cognition
2.3. N1 Habituation and Phonological Information
2.4. The Present Study
3. Materials and Methods
3.1. Participants
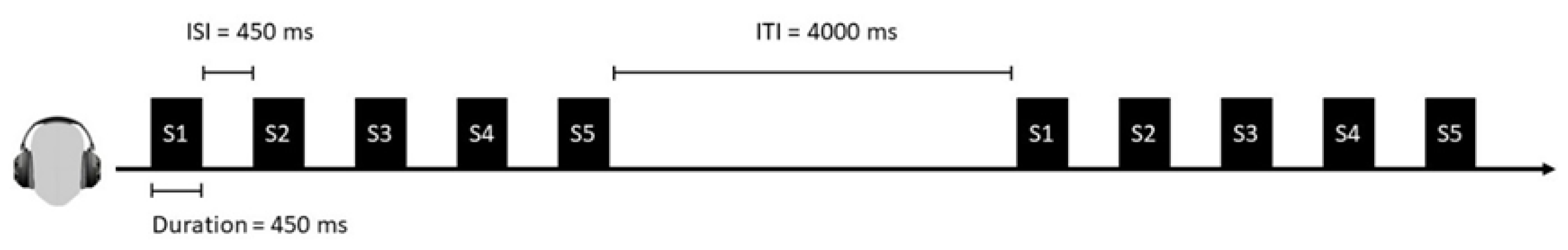
3.2. Design and Materials
3.3. EEG Data Acquisition
3.4. EEG Data Analysis
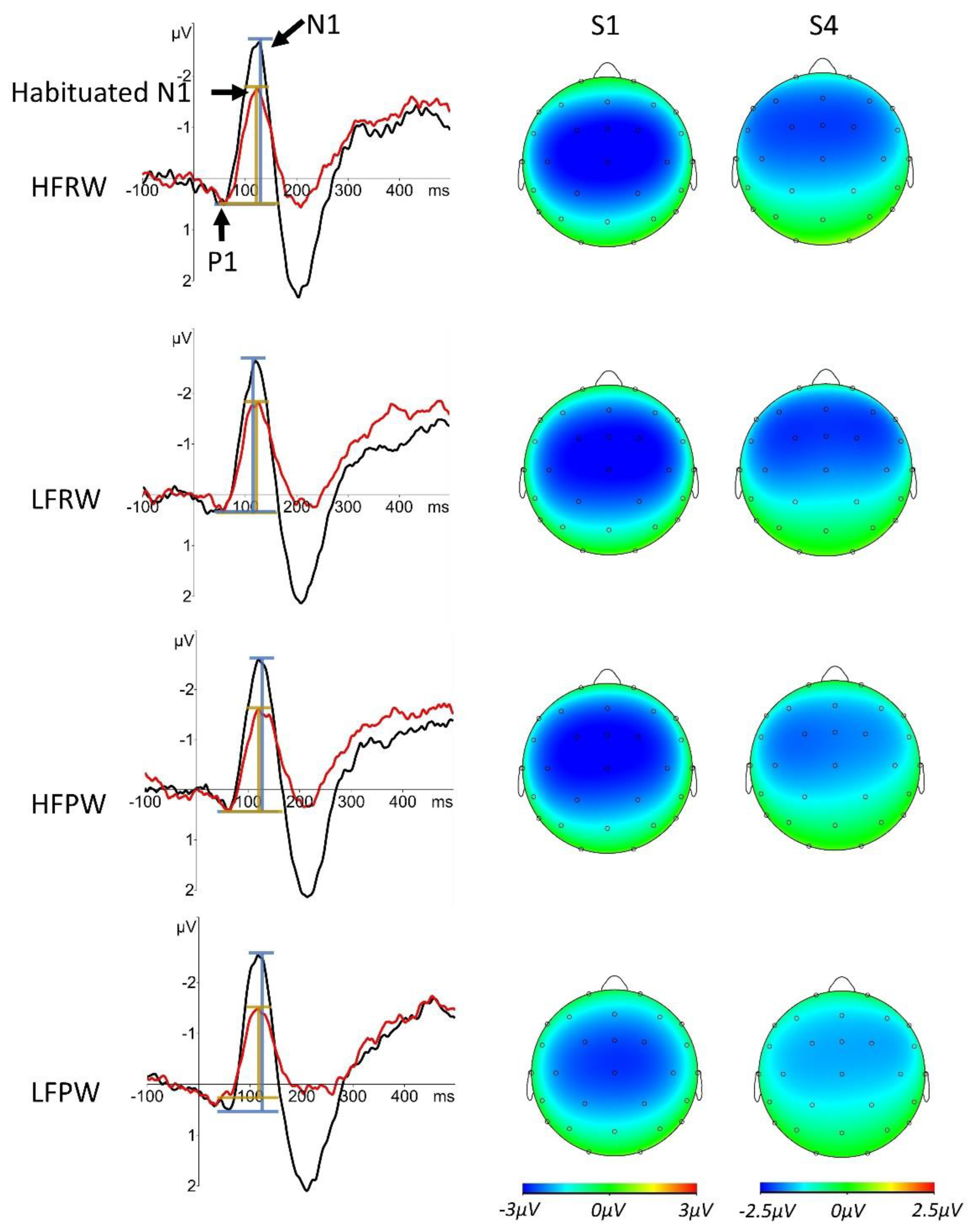
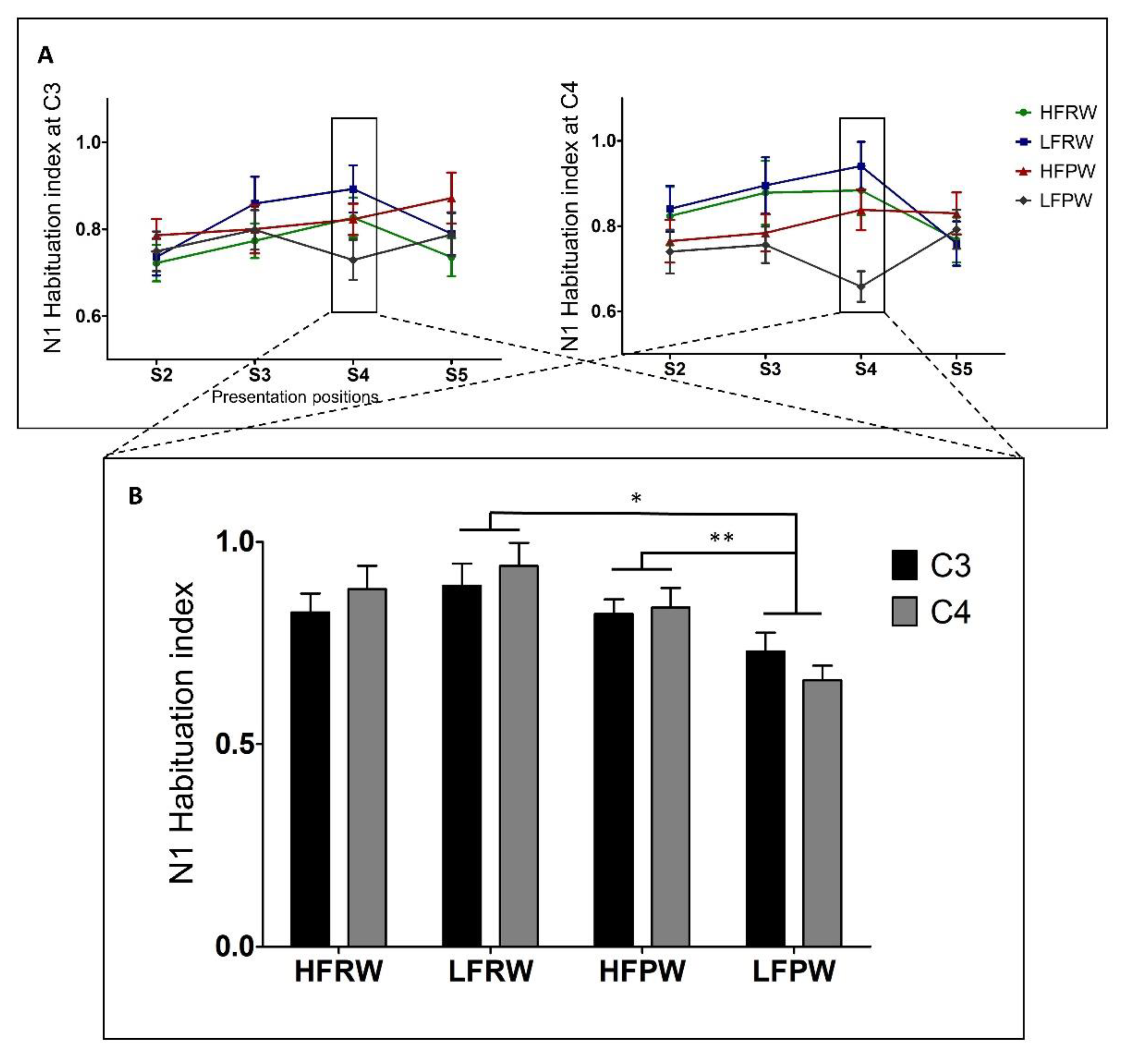
4. Results
5. Discussion
5.1. The Effects of Lexicality and Usage Frequency
5.2. Theoretical Implications
5.3. Limitations and Future Research
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| List1 | List2 | ||||||
|---|---|---|---|---|---|---|---|
| HFRW | HFPW | LFRW | LFPW | HFRW | HFPW | LFRW | LFPW |
| ran2 | mai1 | jiong3 | niang3 | mai3 | ran4 | niang2 | jiong1 |
| chun1 | ru1 | tun1 | diu4 | ru4 | chun4 | diu1 | tun3 |
| che1 | kan2 | mie4 | nie3 | kan4 | che2 | nie1 | mie3 |
| heng2 | lun3 | pie3 | rao1 | lun4 | heng3 | rao4 | pie2 |
| gei3 | nü4 | niao3 | reng3 | nü3 | gei1 | reng2 | niao1 |
| HFRP = high-frequency real word-form, HFPW = high-frequency pseudo word-form, LFRW = low-frequency real word-form, LFPW = low-frequency pseudo word-form. | |||||||
References
- Boutros, N.N.; Gjini, K.; Urbach, H.; Pflieger, M.E. Mapping repetition suppression of the N100 evoked response to the human cerebral cortex. Biol. Psychiatry 2011, 69, 883–889. [Google Scholar] [CrossRef]
- Rosburg, T.; Haueisen, J.; Kreitschmann-Andermahr, I. The dipole location shift within the auditory evoked neuromagnetic field components N100m and mismatch negativity (MMNm). Clin. Neurophysiol. 2004, 115, 906–913. [Google Scholar] [CrossRef]
- De Bruin, N.M.W.J.; Ellenbroek, B.A.; van Schaijk, W.J.; Cools, A.R.; Coenen, A.M.L.; van Luijtelaar, E.L.J.M. Sensory gating of auditory evoked potentials in rats: Effects of repetitive stimulation and the interstimulus interval. Biol. Psychol. 2001, 55, 195–213. [Google Scholar] [CrossRef]
- Larsson, J.; Smith, A.T. fMRI repetition suppression: Neuronal adaptation or stimulus expectation? Cereb. Cortex 2012, 22, 567–576. [Google Scholar] [CrossRef]
- Matsuzaki, N.; Nagasawa, T.; Juhász, C.; Sood, S.; Asano, E. Independent predictors of neuronal adaptation in human primary visual cortex measured with high-gamma activity. Neuroimage 2012, 59, 1639–1646. [Google Scholar] [CrossRef]
- Garrido, M.I.; Kilner, J.M.; Kiebel, S.J.; Stephan, K.E.; Baldeweg, T.; Friston, K.J. Repetition suppression and plasticity in the human brain. Neuroimage 2009, 48, 269–279. [Google Scholar] [CrossRef]
- Groves, P.M.; Thompson, R.F. Habituation: A dual-process theory. Psychol. Rev. 1970, 77, 419. [Google Scholar] [CrossRef]
- Mildner, V. The Cognitive Neuroscience of Human Communication; Lawrence Erlbaum Associates: New York, NY, USA, 2008. [Google Scholar]
- Budd, T.W.; Barry, R.J.; Gordon, E.; Rennie, C.; Michie, P.T. Decrement of the N1 auditory event-related potential with stimulus repetition: Habituation vs. refractoriness. Int. J. Psychophysiol. 1998, 31, 51–68. [Google Scholar] [CrossRef]
- Teismann, I.K.; Sörös, P.; Manemann, E.; Ross, B.; Pantev, C.; Knecht, S. Responsiveness to repeated speech stimuli persists in left but not right auditory cortex. NeuroReport 2004, 15, 1267–1270. [Google Scholar] [CrossRef]
- Muenssinger, J.; Stingl, K.T.; Matuz, T.; Binder, G.; Ehehalt, S.; Preissl, H. Auditory habituation to simple tones: Reduced evidence for habituation in children compared to adults. Front. Hum. Neurosci. 2013, 7, 377. [Google Scholar] [CrossRef] [Green Version]
- Rosburg, T.; Haueisen, J.; Sauer, H. Habituation of the auditory evoked field component N100m and its dependence on stimulus duration. Clin. Neurophysiol. 2002, 113, 421–428. [Google Scholar] [CrossRef]
- Fruhstorfer, H.; Soveri, P.; Järvilehto, T. Short-term habituation of the auditory evoked response in man. Electroencephalogr. Clin. Neurophysiol. 1970, 28, 153–161. [Google Scholar] [CrossRef]
- Woods, D.L.; Elmasian, R. The habituation of event-related potentials to speech sounds and tones. Electroencephalogr. Clin. Neurophysiol. Evoked Potentials Sect. 1986, 65, 447–459. [Google Scholar] [CrossRef]
- Lijffijt, M.; Lane, S.D.; Meier, S.L.; Boutros, N.N.; Burroughs, S.; Steinberg, J.L.; Gerard Moeller, F.; Swann, A.C. P50, N100, and P200 sensory gating: Relationships with behavioral inhibition, attention, and working memory. Psychophysiology 2009, 46, 1059–1068. [Google Scholar] [CrossRef]
- Rosburg, T.; Sörös, P. The response decrease of auditory evoked potentials by repeated stimulation–Is there evidence for an interplay between habituation and sensitization? Clin. Neurophysiol. 2016, 127, 397–408. [Google Scholar] [CrossRef]
- Rust, J. Habituation and the orienting response in the auditory cortical evoked potential. Psychophysiology 1977, 14, 123–126. [Google Scholar] [CrossRef]
- Rosburg, T.; Zimmerer, K.; Huonker, R. Short-term habituation of auditory evoked potential and neuromagnetic field components in dependence of the interstimulus interval. Exp. Brain Res. 2010, 205, 559–570. [Google Scholar] [CrossRef]
- Joos, K.; Gilles, A.; Van de Heyning, P.; De Ridder, D.; Vanneste, S. From sensation to percept: The neural signature of auditory event-related potentials. Neurosci. Biobehav. Rev. 2014, 42, 148–156. [Google Scholar] [CrossRef]
- Fruhstorfer, H. Habituation and dishabituation of the human vertex response. Electroencephalogr. Clin. Neurophysiol. 1971, 30, 306–312. [Google Scholar] [CrossRef]
- Ritter, W.; Vaughan Jr, H.G.; Costa, L.D. Orienting and habituation to auditory stimuli: A study of short terms changes in average evoked responses. Electroencephalogr. Clin. Neurophysiol. 1968, 25, 550–556. [Google Scholar] [CrossRef]
- Sara, Y.; Mozhayeva, M.G.; Liu, X.; Kavalali, E.T. Fast vesicle recycling supports neurotransmission during sustained stimulation at hippocampal synapses. J. Neurosci. 2002, 22, 1608–1617. [Google Scholar] [CrossRef]
- Wang, P.; Knösche, T.R. A realistic neural mass model of the cortex with laminar-specific connections and synaptic plasticity–evaluation with auditory habituation. PLoS ONE 2013, 8, e77876. [Google Scholar] [CrossRef]
- Marklund, E.; Gustavsson, L.; Kallioinen, P.; Schwarz, I.-C. N1 Repetition-Attenuation for Acoustically Variable Speech and Spectrally Rotated Speech. Front. Hum. Neurosci. 2020, 14, 534804. [Google Scholar] [CrossRef]
- Yue, J.; Alter, K.; Howard, D.; Bastiaanse, R. Early access to lexical-level phonological representations of Mandarin word-forms: Evidence from auditory N1 habituation. Lang. Cogn. Neurosci. 2017, 32, 1148–1163. [Google Scholar] [CrossRef]
- Öhman, A.; Lader, M. Selective attention and “habituation” of the auditory averaged evoked response in humans. Physiol. Behav. 1972, 8, 79–85. [Google Scholar] [CrossRef]
- Font-Alaminos, M.; Cornella, M.; Costa-Faidella, J.; Hervás, A.; Leung, S.; Rueda, I.; Escera, C. Increased subcortical neural responses to repeating auditory stimulation in children with autism spectrum disorder. Biol. Psychol. 2020, 149, 107807. [Google Scholar] [CrossRef]
- Herrmann, B.; Maess, B.; Johnsrude, I.S. Aging affects adaptation to sound-level statistics in human auditory cortex. J. Neurosci. 2018, 38, 1989–1999. [Google Scholar] [CrossRef]
- Todorovic, A.; van Ede, F.; Maris, E.; de Lange, F.P. Prior expectation mediates neural adaptation to repeated sounds in the auditory cortex: An MEG study. J. Neurosci. 2011, 31, 9118–9123. [Google Scholar] [CrossRef]
- Gansonre, C.; Højlund, A.; Leminen, A.; Bailey, C.; Shtyrov, Y. Task-free auditory EEG paradigm for probing multiple levels of speech processing in the brain. Psychophysiology 2018, 55, e13216. [Google Scholar] [CrossRef]
- Shtyrov, Y.; Lenzen, M. First-pass neocortical processing of spoken language takes only 30 msec: Electrophysiological evidence. Cogn. Neurosci. 2017, 8, 24–38. [Google Scholar] [CrossRef]
- Slowiaczek, L.M.; Hamburger, M. Prelexical facilitation and lexical interference in auditory word recognition. J. Exp. Psychol. Learn. Mem. Cogn. 1992, 18, 1239. [Google Scholar] [CrossRef]
- Wagenmakers, E.J.M.; Zeelenberg, R.; Steyvers, M.; Shiffrin, R.; Raaijmakers, J.G. Nonword repetition in lexical decision: Support for two opposing processes. Q. J. Exp. Psychol. 2004, 57, 1191–1210. [Google Scholar] [CrossRef]
- Howard, D.; Patterson, K.; Wise, R.; Brown, W.D.; Friston, K.; Weiller, C.; Frackowiak, R. The cortical localization of the lexicons: Positron emission tomography evidence. Brain 1992, 115, 1769–1782. [Google Scholar] [CrossRef]
- Myers, E.B.; Blumstein, S.E.; Walsh, E.; Eliassen, J. Inferior frontal regions underlie the perception of phonetic category invariance. Psychol. Sci. 2009, 20, 895–903. [Google Scholar] [CrossRef]
- Orfanidou, E.; Marslen-Wilson, W.D.; Davis, M.H. Neural response suppression predicts repetition priming of spoken words and pseudowords. J. Cogn. Neurosci. 2006, 18, 1237–1252. [Google Scholar] [CrossRef]
- Pulvermüller, F.; Kujala, T.; Shtyrov, Y.; Simola, J.; Tiitinen, H.; Alku, P.; Alho, K.; Martinkauppi, S.; Ilmoniemi, R.J.; Näätänen, R. Memory traces for words as revealed by the mismatch negativity. Neuroimage 2001, 14, 607–616. [Google Scholar] [CrossRef]
- Shtyrov, Y.; Kimppa, L.; Pulvermüller, F.; Kujala, T. Event-related potentials reflecting the frequency of unattended spoken words: A neuronal index of connection strength in lexical memory circuits? Neuroimage 2011, 55, 658–668. [Google Scholar] [CrossRef]
- Jacobsen, T.; Horváth, J.; Schröger, E.; Lattner, S.; Widmann, A.; Winkler, I. Pre-attentive auditory processing of lexicality. Brain Lang. 2004, 88, 54–67. [Google Scholar] [CrossRef]
- Yue, J.; Bastiaanse, R.; Alter, K. Cortical plasticity induced by rapid Hebbian learning of novel tonal word-forms: Evidence from mismatch negativity. Brain Lang. 2014, 139, 10–22. [Google Scholar] [CrossRef]
- Aleksandrov, A.A.; Memetova, K.S.; Stankevich, L.N.; Knyazeva, V.M.; Shtyrov, Y. Referent’s lexical frequency predicts mismatch negativity responses to new words following semantic training. J. Psycholinguist. Res. 2020, 49, 187–198. [Google Scholar] [CrossRef]
- Silva, S.; Vigário, M.; Fernandez, B.L.; Jerónimo, R.; Alter, K.; Frota, S. The sense of sounds: Brain responses to phonotactic frequency, phonological grammar and lexical meaning. Front. Psychol. 2019, 10, 681. [Google Scholar] [CrossRef]
- Duanmu, S. The Phonology of Standard Chinese; Oxford University Press: Oxford, England, 2007. [Google Scholar]
- Yue, J. Tone-Word Recognition in Mandarin Chinese: Influences of Lexical-Level Representations. Ph.D. Thesis, University of Groningen, Groningen, The Netherlands, 2016. [Google Scholar]
- Dufour, S. Phonological priming in auditory word recognition: When both controlled and automatic processes are responsible for the effects. Can. J. Exp. Psychol. Rev. Can. De Psychol. Expérimentale 2008, 62, 33. [Google Scholar] [CrossRef]
- Ziegler, J.C.; Tan, L.H.; Perry, C.; Montant, M. Phonology matters: The phonological frequency effect in written Chinese. Psychol. Sci. 2000, 11, 234–238. [Google Scholar] [CrossRef]
- Balota, D.A.; Cortese, M.J.; Sergent-Marshall, S.D.; Spieler, D.H.; Yap, M.J. Visual word recognition of single-syllable words. J. Exp. Psychol. Gen. 2004, 133, 283. [Google Scholar] [CrossRef]
- Perea, M.; Rosa, E.; Gómez, C. The frequency effect for pseudowords in the lexical decision task. Percept. Psychophys. 2005, 67, 301–314. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- McEnery, A.; Xiao, Z. The Lancaster Corpus of Mandarin Chinese: A Corpus for Monolingual and Contrastive Language Study. In Proceedings of the Fourth International Conference on Language Resources and Evaluation (LREC), Centro Cultural de Belem, Lisbon, Portugal, 26–28 May 2004; pp. 1175–1178. [Google Scholar]
- Shu, H.; Chen, X.; Anderson, R.C.; Wu, N.; Xuan, Y. Properties of school Chinese: Implications for learning to read. Child Dev. 2003, 74, 27–47. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.X.; Kong, L. An ERP study on the time course of phonological and semantic activation in Chinese word recognition. Int. J. Psychophysiol. 2009, 73, 235–245. [Google Scholar] [CrossRef]
- Wiener, S.; Ito, K. Do syllable-specific tonal probabilities guide lexical access? Evidence from Mandarin, Shanghai and Cantonese speakers. Lang. Cogn. Neurosci. 2015, 30, 1048–1060. [Google Scholar] [CrossRef]
- Wiener, S.; Turnbull, R. Constraints of tones, vowels and consonants on lexical selection in Mandarin Chinese. Lang. Speech 2016, 59, 59–82. [Google Scholar] [CrossRef] [Green Version]
- Boersma, P.; Weenink, D. Praat: Doing Phonetics by Computer (Version 5.3.39); Institute of Phonetic Sciences of the University of Amsterdam: Amsterdam, The Netherlands, 2013; Available online: http://www.praat.org (accessed on 15 September 2022).
- McClaskey, C.M.; Dias, J.W.; Dubno, J.R.; Harris, K.C. Reliability of measures of N1 peak amplitude of the compound action potential in younger and older adults. J. Speech Lang. Heart Res. 2018, 61, 2422–2430. [Google Scholar] [CrossRef]
- Toyokura, M. Influence of stimulus intensity on waveform of sympathetic skin response evoked by magnetic stimulation. Clin. Neurophysiol. 2003, 114, 1423–1430. [Google Scholar] [CrossRef]
- Swink, S.; Stuart, A. Auditory long latency responses to tonal and speech stimuli. J. Speech Lang. Heart Res. 2012, 55, 447–459. [Google Scholar] [CrossRef]
- Grau, C.; Fuentemilla, L.; Marco-Pallarés, J. Functional neural dynamics underlying auditory event-related N1 and N1 suppression response. Neuroimage 2007, 36, 522–531. [Google Scholar] [CrossRef]
- Pulvermüller, F.; Shtyrov, Y. Language outside the focus of attention: The mismatch negativity as a tool for studying higher cognitive processes. Prog. Neurobiol. 2006, 79, 49–71. [Google Scholar] [CrossRef]
- Rankin, C.H.; Abrams, T.; Barry, R.J.; Bhatnagar, S.; Clayton, D.F.; Colombo, J.; Coppola, G.; Geyer, M.A.; Glanzman, D.L.; Marsland, S. Habituation revisited: An updated and revised description of the behavioral characteristics of habituation. Neurobiol. Learn. Mem. 2009, 92, 135–138. [Google Scholar] [CrossRef]
- Gaskell, M.G.; Marslen-Wilson, W.D. Phonological variation and inference in lexical access. J. Exp. Psychol. Hum. Percept. Perform. 1996, 22, 144. [Google Scholar] [CrossRef]
- McMurray, B.; Jongman, A. What information is necessary for speech categorization? Harnessing variability in the speech signal by integrating cues computed relative to expectations. Psychol. Rev. 2011, 118, 219. [Google Scholar] [CrossRef]
- Apfelbaum, K.S.; McMurray, B. Using variability to guide dimensional weighting: Associative mechanisms in early word learning. Cogn. Sci. 2011, 35, 1105–1138. [Google Scholar] [CrossRef]
- Goswami, U. Early phonological development and the acquisition of literacy. In Handbook of Early Literacy Research; Neuman, S.B., Dickinson, D.K., Eds.; The Guildfor Press: New York, NY, USA, 2001; pp. 111–125. [Google Scholar]
- Aleksandrov, A.A.; Memetova, K.S.; Stankevich, L.N.; Uplisova, K.O. Effects of Russian-language word frequency on mismatch negativity in auditory event-related potentials. Neurosci. Behav. Physiol. 2017, 47, 1043–1050. [Google Scholar] [CrossRef]
- Luo, H.; Ni, J.-T.; Li, Z.-H.; Li, X.-O.; Zhang, D.-R.; Zeng, F.-G.; Chen, L. Opposite patterns of hemisphere dominance for early auditory processing of lexical tones and consonants. Proc. Natl. Acad. Sci. USA 2006, 103, 19558–19563. [Google Scholar] [CrossRef]
- Malins, J.G.; Joanisse, M.F. Setting the tone: An ERP investigation of the influences of phonological similarity on spoken word recognition in Mandarin Chinese. Neuropsychologia 2012, 50, 2032–2043. [Google Scholar] [CrossRef]
- Wang, X.-D.; Gu, F.; He, K.; Chen, L.-H.; Chen, L. Preattentive extraction of abstract auditory rules in speech sound stream: A mismatch negativity study using lexical tones. PLoS ONE 2012, 7, e30027. [Google Scholar] [CrossRef]
- Joanisse, M.F.; Zevin, J.D.; McCandliss, B.D. Brain mechanisms implicated in the preattentive categorization of speech sounds revealed using fMRI and a short-interval habituation trial paradigm. Cereb. Cortex 2007, 17, 2084–2093. [Google Scholar] [CrossRef]
- Vaden Jr, K.I.; Muftuler, L.T.; Hickok, G. Phonological repetition-suppression in bilateral superior temporal sulci. Neuroimage 2010, 49, 1018–1023. [Google Scholar] [CrossRef]
- Kringelbach, M.L.; Deco, G. Brain states and transitions: Insights from computational neuroscience. Cell Rep. 2020, 32, 108128. [Google Scholar] [CrossRef]
- Shtyrov, Y.; Pulvermüller, F. Language in the mismatch negativity design: Motivations, benefits, and prospects. J. Psychophysiol. 2007, 21, 176. [Google Scholar] [CrossRef]
- Kessier, C.; Steinberg, A. Evoked potential variation in schizophrenic subgroups. Biol. Psychiatry 1989, 26, 372–380. [Google Scholar] [CrossRef]
- Choi, W.; Lim, M.; Kim, J.; Chung, C. Habituation deficit of auditory N100m in patients with fibromyalgia. Eur. J. Pain 2016, 20, 1634–1643. [Google Scholar] [CrossRef]
- Ethridge, L.E.; White, S.P.; Mosconi, M.W.; Wang, J.; Byerly, M.J.; Sweeney, J. Reduced habituation of auditory evoked potentials indicate cortical hyper-excitability in Fragile X Syndrome. Transl. Psychiatry 2016, 6, e787. [Google Scholar] [CrossRef]
- Wu, N.; Shu, H. The gating paradigm and spoken word recognition of Chinese (Gating). Acta Psychol. Sin. 2003, 35, 582–590. [Google Scholar]
- Fabiani, M.; Low, K.A.; Wee, E.; Sable, J.J.; Gratton, G. Reduced suppression or labile memory? Mechanisms of inefficient filtering of irrelevant information in older adults. J. Cogn. Neurosci. 2006, 18, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Melara, R.D.; Root, J.C.; Bibi, R.; Ahles, T.A. Sensory filtering and sensory memory in breast cancer survivors. Clin. EEG Neurosci. 2021, 52, 246–253. [Google Scholar] [CrossRef]
- Sambeth, A.; Maes, J.; Quiroga, R.Q.; Coenen, A.M. Effects of stimulus repetitions on the event-related potential of humans and rats. Int. J. Psychophysiol. 2004, 53, 197–205. [Google Scholar] [CrossRef]
- Liberman, A.M.; Cooper, F.S.; Shankweiler, D.P.; Studdert-Kennedy, M. Perception of the speech code. Psychol. Rev. 1967, 74, 431–461. [Google Scholar] [CrossRef]
- Samuel, A.G. Knowing a word affects the fundamental perception of the sounds within it. Psychol. Sci. 2001, 12, 348–351. [Google Scholar] [CrossRef]
- Dahan, D.; Magnuson, J.S. Spoken-word recognition. In Handbook of Psycholinguistics; Traxler, M.J., Gernsbacher, M.A., Eds.; Academic Press: Amsterdam, The Netherlands, 2006; pp. 249–283. [Google Scholar]
- McClelland, J.L.; Elman, J.L. The TRACE model of speech perception. Cogn. Psychol. 1986, 18, 1–86. [Google Scholar] [CrossRef]
- Davis, M.H.; Sohoglu, E. Three functions of prediction error for Bayesian inference in speech perception. In The Cognitive Neurosciences, 6th ed.; Gazzaniga, M.S., Mangun, G.R., Poeppel, D., Eds.; MIT Press: Cambridge, MA, USA, 2020; pp. 177–189. [Google Scholar]




| Condition | Electrode | Amplitude S1 (μv) | Habituation Index S2 | Habituation Index S3 | Habituation Index S4 | Habituation Index S5 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| M | SD | M | SD | M | SD | M | SD | M | SD | ||
| HFRW | C3 | 4.38 | 1.65 | 0.72 | 0.23 | 0.77 | 0.22 | 0.83 | 0.25 | 0.74 | 0.24 |
| C4 | 4.05 | 1.66 | 0.82 | 0.37 | 0.88 | 0.41 | 0.88 | 0.32 | 0.77 | 0.29 | |
| LFRW | C3 | 4.19 | 1.83 | 0.74 | 0.23 | 0.86 | 0.34 | 0.89 | 0.30 | 0.79 | 0.27 |
| C4 | 3.68 | 1.53 | 0.84 | 0.30 | 0.90 | 0.36 | 0.94 | 0.31 | 0.76 | 0.29 | |
| HFPW | C3 | 4.18 | 1.61 | 0.79 | 0.20 | 0.80 | 0.31 | 0.82 | 0.20 | 0.87 | 0.32 |
| C4 | 4.10 | 1.66 | 0.76 | 0.27 | 0.78 | 0.24 | 0.84 | 0.26 | 0.83 | 0.27 | |
| LFPW | C3 | 4.32 | 1.59 | 0.75 | 0.25 | 0.80 | 0.25 | 0.73 | 0.25 | 0.79 | 0.27 |
| C4 | 4.20 | 1.64 | 0.74 | 0.28 | 0.76 | 0.23 | 0.66 | 0.20 | 0.79 | 0.25 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, J.; Wang, P.; Li, J.; Li, Z.; Liang, X.; He, Y. Short-Term Habituation of Auditory N1 in Spoken Word-Forms Is Modulated by Phonological Information. Brain Sci. 2022, 12, 1279. https://doi.org/10.3390/brainsci12101279
Yue J, Wang P, Li J, Li Z, Liang X, He Y. Short-Term Habituation of Auditory N1 in Spoken Word-Forms Is Modulated by Phonological Information. Brain Sciences. 2022; 12(10):1279. https://doi.org/10.3390/brainsci12101279
Chicago/Turabian StyleYue, Jinxing, Peng Wang, Jiayin Li, Zhipeng Li, Xia Liang, and Yifei He. 2022. "Short-Term Habituation of Auditory N1 in Spoken Word-Forms Is Modulated by Phonological Information" Brain Sciences 12, no. 10: 1279. https://doi.org/10.3390/brainsci12101279