
From Action to Cognition: Neural Reuse, Network Theory and the Emergence of Higher Cognitive Functions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Local or Distributed: Two Views of the Structure-Function Relationship
3. Theories of Neural Reuse
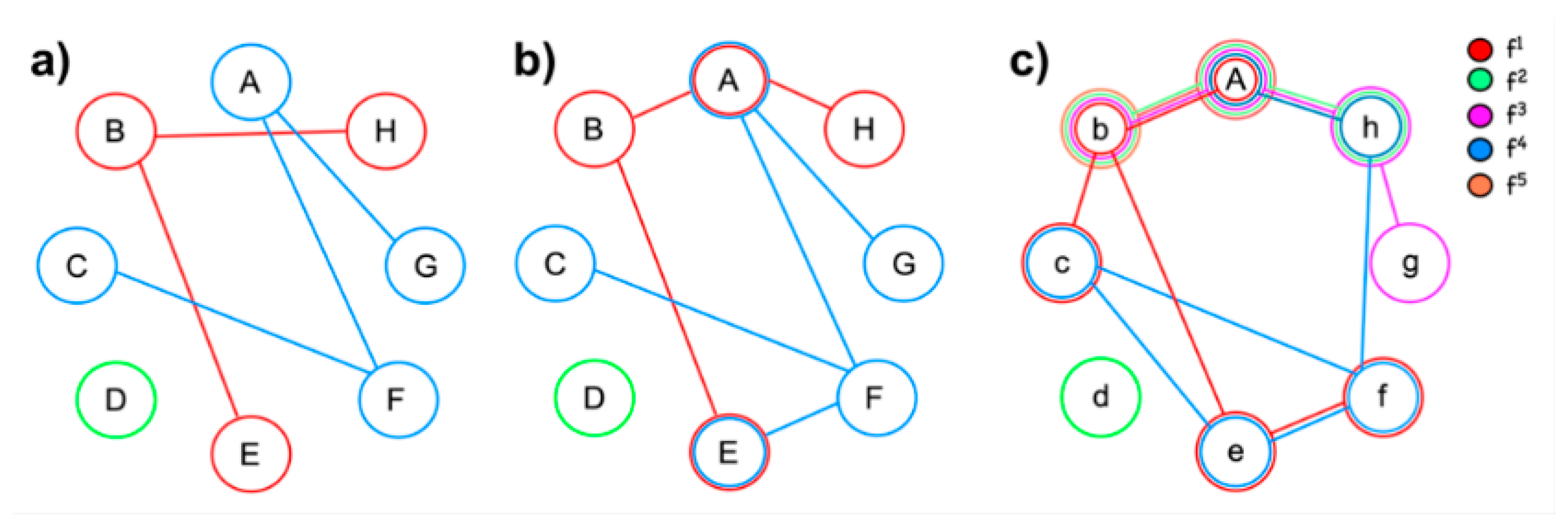
4. Functional Networks Provide the Necessary Conditions for Neural Reuse
- Networks consist of nodes and connections, whereby some nodes (‘hubs’) are of greater importance than others and some connections are stronger than others [63,64,65]. The intrinsic organization of networks enables processing information along a continuum of encapsulation: they may adopt a centralized and modular state (segregation) or become more penetrable to influences from other networks (integration; [66]). Integration and segregation of information processing are thus expressions of the current state, not a fixed feature of networks. Note that the term ‘modular’, when applied to networks, relates to the formation of subgroups of nodes forming a community within a network [67].
- A network structure appears at rest or during activity. The high degree of energy consumption during ‘rest’ indicates that a large quantity of information processing is intrinsic and occurs without external stimulation [68].
- Though the spatial and temporal stability of networks is under debate [69,70], at least some studies using task-based connectivity suggest temporal changes of node weights and network topology over time [71,72,73]. There is also evidence of interindividual variability, yet intraindividual stability of networks [69]. This corresponds to one of Edelman’s requirements for theories of brain function, namely to explain ‘how both perceptual and conceptual categorization can arise as a result of selection upon preexisting variance in structure and function of the nervous system’ ([14], p. 115).
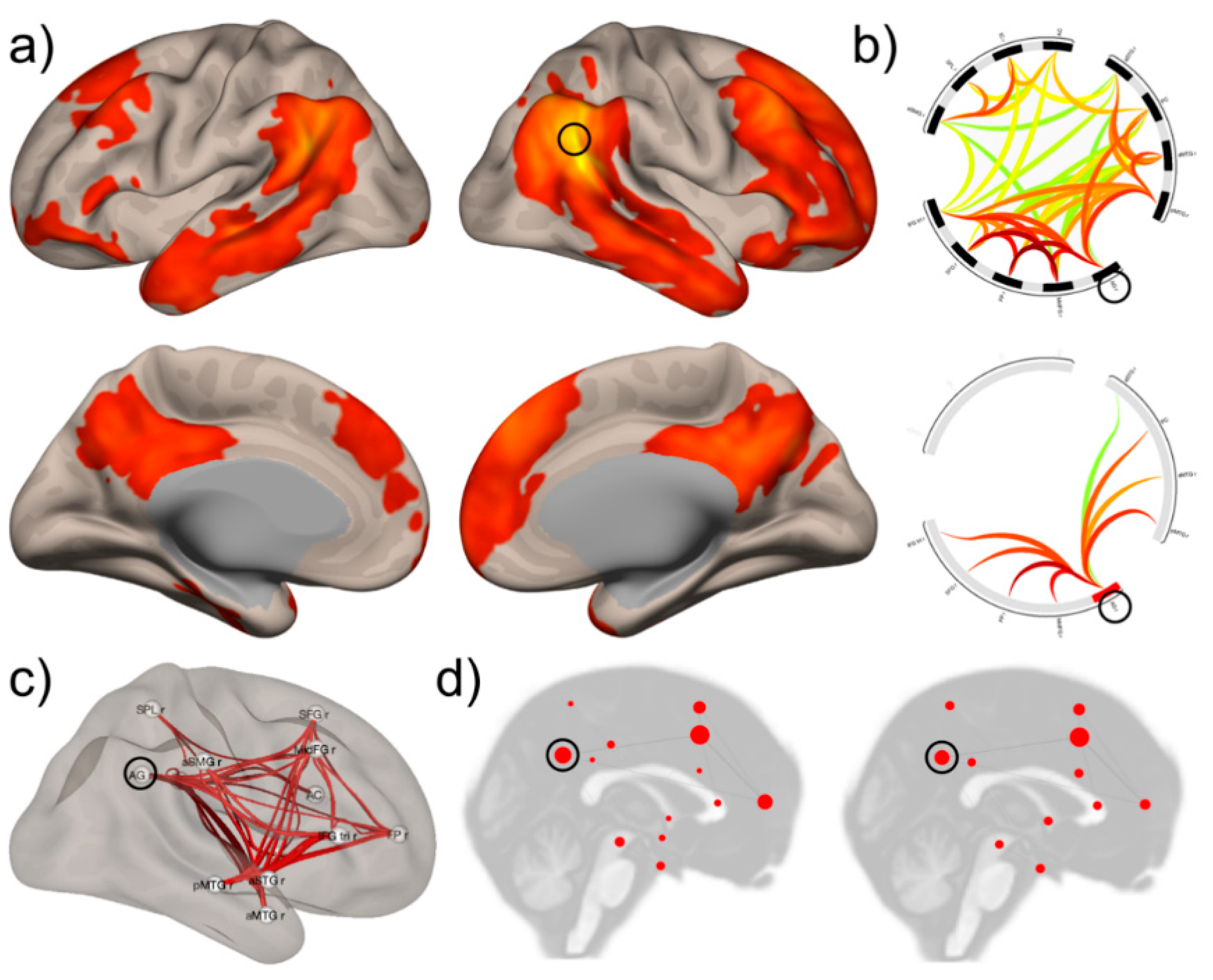
5. Integrating Neural Data and Behavior: The Case of Visual Object Processing
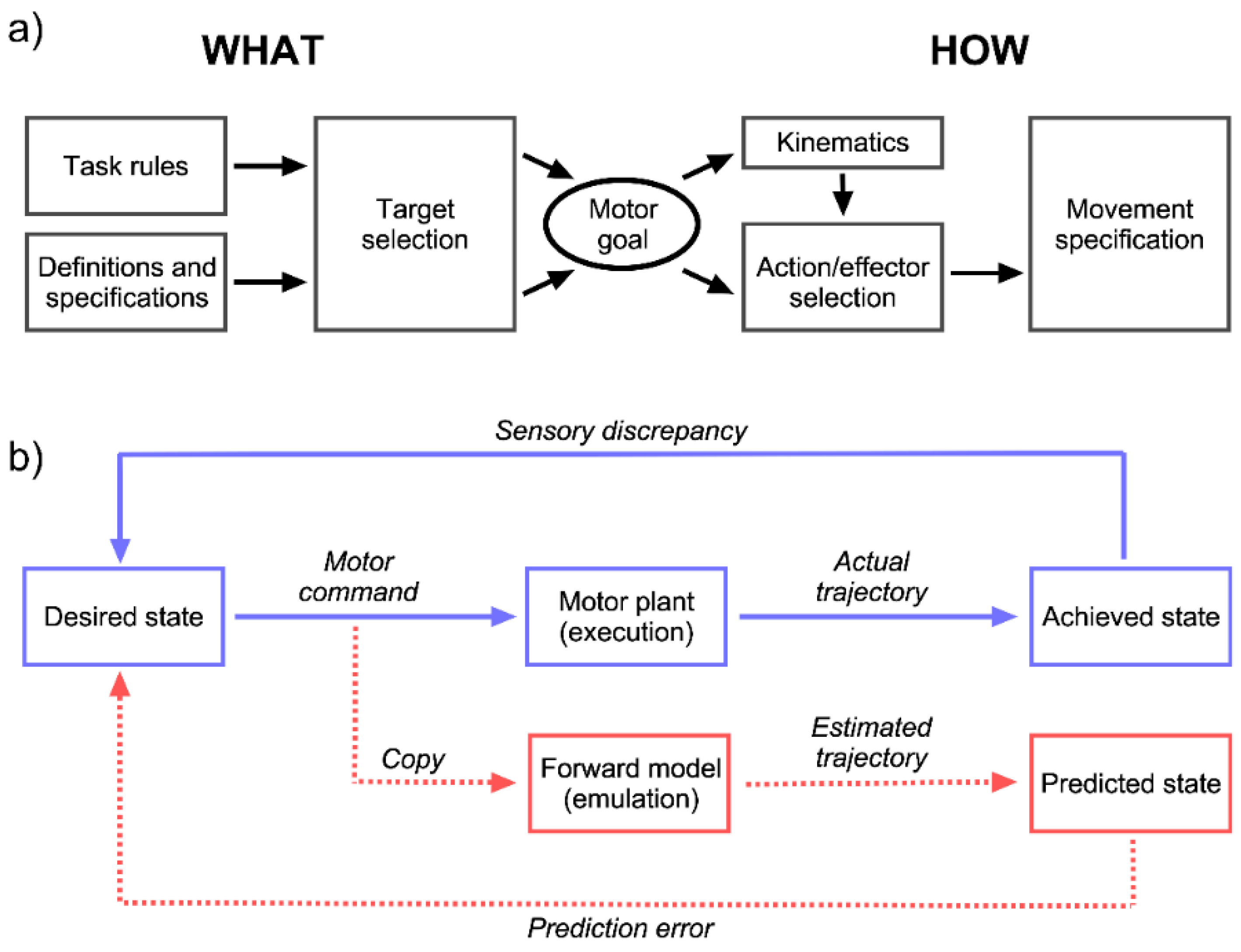
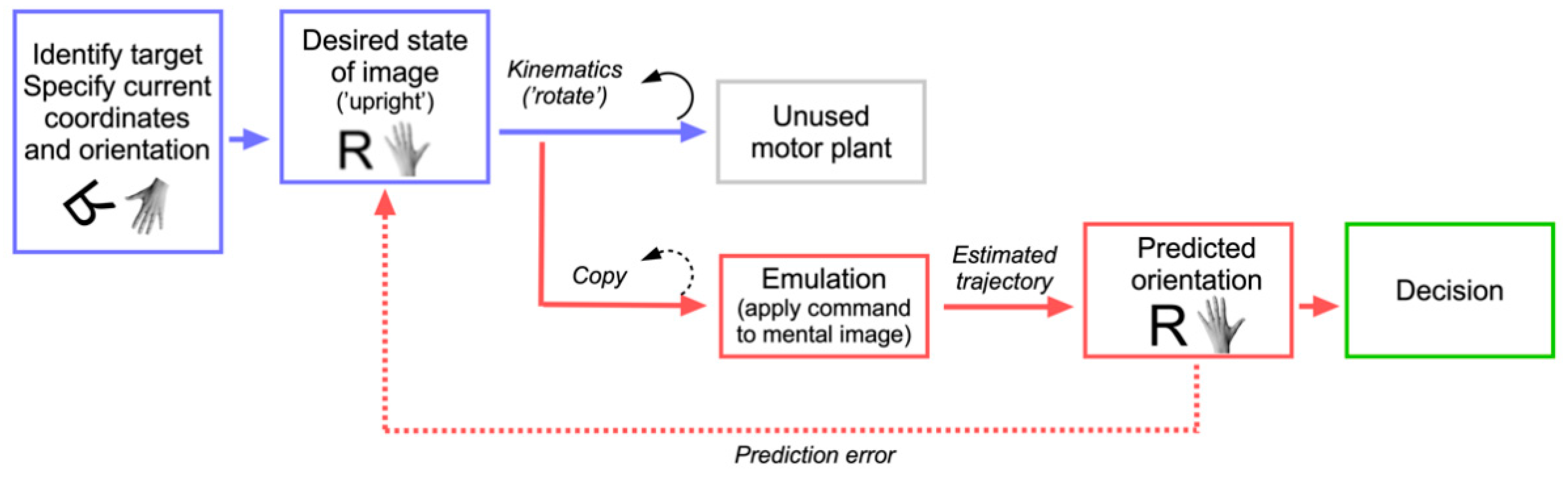
6. A Motor Process for a Cognitive Function: The Emulation Theory of Mental Rotation
- Development of mental rotation ability depends on the maturation of the motor system and only becomes available once the child is capable of replaying actions mentally.
- Mental rotation and motor execution produce comparable patterns of performance. For example, the effect of rotational angle in mental rotation of hands is proportional to its effect when subjects actually perform hand rotations. Hand positions that are difficult or impossible to imitate produce particularly increased reaction times.
- An overlap exists between neural structures involved in motor planning and mental rotation.
- Motor planning and mental rotation use similar resources, resulting in interference when both are performed simultaneously.
- Mental rotation of objects relies on the same kinematic plan as the rotation of body parts, except for the involvement of an effector component in the latter.
7. Conclusions, Limitations and Open Questions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Karmiloff-Smith, A. Development itself is the key to understanding developmental disorders. Trends Cogn. Sci. 1998, 2, 389–398. [Google Scholar] [CrossRef]
- Barsalou, L.W. Grounded cognition. Annu. Rev. Psychol. 2008, 59, 617–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallese, V.; Lakoff, G. The Brain’s concepts: The role of the Sensory-motor system in conceptual knowledge. Cogn. Neuropsychol. 2005, 22, 455–479. [Google Scholar] [CrossRef]
- Dehaene, S.; Cohen, L. Cultural recycling of cortical maps. Neuron 2007, 56, 384–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.L. Neural reuse: A fundamental organizational principle of the brain. Behav. Brain Sci. 2010, 33, 245–313. [Google Scholar] [CrossRef] [Green Version]
- Broca, P. Perte de la parole, ramolissement chronique et destruction partielle du lobe antérieur gauche du cerveau. Bull. Soc. Fr. D’anthropol. 1861, 2, 235–238. [Google Scholar]
- Lissauer, H. Ein Fall von Seelenblindheit nebst einem Beitrag zur Theorie derselben. Arch. Psychiatr. 1890, 21, 222–270. [Google Scholar] [CrossRef]
- Bálint, R. Seelenlähmung des ‘Schauens’, optische Ataxie, räumliche Störung der Aufmerksamkeit. Msschr. Psychiat. Neurol. 1909, 25, 51–66. [Google Scholar] [CrossRef] [Green Version]
- Gerstmann, J. Reine taktile Agnosie. Mschr. Psychiat. Neurol. 1918, 44, 330–343. [Google Scholar] [CrossRef]
- Lashley, K.S. Basic neural mechanisms in behavior. Psychol. Rev. 1930, 37, 1–24. [Google Scholar] [CrossRef]
- Lashley, K.S. Integrative functions of the cerebral cortex. Physiol. Rev. 1933, 13, 1–42. [Google Scholar] [CrossRef] [Green Version]
- Geschwind, N. Disconnection syndromes in animals and man. Part II. Brain 1965, 88, 585–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geschwind, N. Disconnexion syndromes in animals and man. Part I. Brain 1965, 88, 237–294. [Google Scholar] [CrossRef] [Green Version]
- Edelman, G.M. Neural Darwinism: Selection and reentrant signaling in higher brain function. Neuron 1993, 10, 115–125. [Google Scholar] [CrossRef]
- Di Lollo, V.; Enns, J.T.; Rensink, R.A. Competition for consciousness among visual events: The psychophysics of reentrant visual processes. J. Exp. Psychol. Gen. 2000, 129, 481–507. [Google Scholar] [CrossRef]
- Price, C.J.; Friston, K.J. Degeneracy and cognitive anatomy. Trends Cogn. Sci. 2002, 6, 416–421. [Google Scholar] [CrossRef]
- Tononi, G.; Sporns, O.; Edelman, G.M. Measures of degeneracy and redundancy in biological networks. Proc. Natl. Acad. Sci. USA 1999, 96, 3257–3262. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, S. Parallel distributed processing. Nature 1986, 323, 486. [Google Scholar] [CrossRef]
- Farah, M.J. Neuropsychological inference with an interactive brain: A critique of the “locality” assumption. Behav. Brain Sci. 1994, 17, 43–104. [Google Scholar] [CrossRef]
- McClelland, J.L. The organization of memory. A Parallel Distributed Processing perspective. Rev. Neurol. 1994, 150, 570–579. [Google Scholar]
- Bibby, P.A. Distributed Knowledge: In the Head, in the World or in the Interaction? In Models in the Mind. Theory, Perspective and Applications; Academic Press: New York, NY, USA, 1992; pp. 93–99. [Google Scholar]
- Zeki, S.; Shipp, S. The functional logic of cortical connections. Nature 1988, 335, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.K., Jr. Mass function and equipotentiality: A reanalysis of Lashley’s retention data. Psychol. Rep. 1970, 27, 899–902. [Google Scholar] [CrossRef]
- Milner, B.; Corkin, S.; Teuber, H.-L. Further analysis of the hippocampal amnesic syndrome: 14-year follow-up study of H.M. Neuropsychologia 1968, 6, 215–234. [Google Scholar] [CrossRef]
- Scoville, W.B.; Milner, B. Loss of recent memory after bilateral hippocampal lesions. J. Neurol. Neurosurg. Psychiatry 1957, 20, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Bornstein, B.; Kindron, D.P. Prosopagnosia. J. Neurol. Neurosurg. Psychiatry 1959, 22, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Luria, A.R. Disorders of "simultaneous perception” in a case of bilateral occipito-parietal brain injury. Brain 1959, 82, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Kinsbourne, M.; Warrington, E.K. A study of finger agnosia. Brain 1962, 85, 47–66. [Google Scholar] [CrossRef]
- Gazzaniga, M.S.; Bogen, J.E.; Sperry, R.W. Observations on visual perception after disconnexion of the cerebral hemispheres in man. Brain 1965, 88, 221–236. [Google Scholar] [CrossRef] [Green Version]
- Caramazza, A. The Logic of Neuropsychological Research and the Problem of Patient Classification in Aphasia. Brain. Lang. 1984, 21, 9–20. [Google Scholar] [CrossRef]
- Caramazza, A. On Drawing Inferences about the Structure of Normal Cognitive Systems from the Analysis of Patterns of Impaired Performance: The Case for Single-Patient Studies. Brain. Cogn. 1986, 5, 41–66. [Google Scholar] [CrossRef]
- Sternberg, S. The discovery of processing stages: Extensions of Donder’s method. Acta Psychol. 1969, 30, 276–315. [Google Scholar] [CrossRef]
- Sternberg, S. Modular processes in mind and brain. Cogn. Neuropsychol. 2011, 28, 156–208. [Google Scholar] [CrossRef] [Green Version]
- Fodor, J.A. The Modularity of Mind; MIT Press: Cambridge, MA, USA, 1983. [Google Scholar]
- Shallice, T. From Neuropsychology to Mental Structure; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Kanwisher, N.; McDermott, J.; Chun, M.M. The fusiform face area: A module in human extrastriate cortex specialized for face perception. J. Neurosci. 1997, 17, 4302–4311. [Google Scholar] [CrossRef]
- Epstein, R.; Kanwisher, N. A cortical representation of the local visual environment. Nature 1998, 392, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Downing, P.E.; Jiang, Y.; Shuman, M.; Kanwisher, N. A cortical area selective for visual processing of the human body. Science 2001, 293, 2470–2473. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.L.; Young, A.W.; Senior, C.; Brammer, M.; Andrew, C.; Calder, A.J.; Bullmore, E.T.; Perrett, D.I.; Rowland, D.; Williams, S.D.R.; et al. A specific neural substrate for perceiving facial expressions of disgust. Nature 1997, 389, 495–498. [Google Scholar] [CrossRef]
- Anderson, M.L. Massive redeployment, exaptation, and the functional integration of cognitive operations. Synthese 2007, 159, 329–345. [Google Scholar] [CrossRef] [Green Version]
- Quartz, S.R.; Sejnowski, T.J. The neural basis of cognitive development: A constructivist manifesto. Behav. Brain Sci. 1997, 20, 537–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeannerod, M. Neural simulation of action: A unifying mechanism for motor cognition. NeuroImage 2001, 14, S103–S109. [Google Scholar] [CrossRef] [PubMed]
- Thelen, E. Grounded in the world: Developmental origins of the embodied mind. Infancy 2000, 1, 3–28. [Google Scholar] [CrossRef]
- Wilson, M. Six views of embodied cognition. Psychon. Bull. Rev. 2002, 9, 625–636. [Google Scholar] [CrossRef]
- Gallese, V.; Goldman, A. Mirror neurons and the simulation theory of mind-reading. Trends Cogn. Sci. 1998, 2, 493–501. [Google Scholar] [CrossRef]
- Rizzolatti, G.; Arbib, M.A. Language within our grasp. Trends Neurosci. 1998, 21, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Hauk, O.; Johnsrude, I.; Pulvermüller, F. Somatotopic representation of action words in human motor and premotor cortex. Neuron 2004, 41, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Moore, T.; Armstrong, K.M.; Fallah, M. Visuomotor origins of covert spatial attention. Neuron 2003, 40, 671–683. [Google Scholar] [CrossRef] [Green Version]
- Rizzolatti, G.; Riggio, L.; Dascola, I.; Umiltà, C. Reorienting attention across the horizontal and vertical meridians: Evidence in favor of a premotor theory of attention. Neuropsychologia 1987, 25, 31–40. [Google Scholar] [CrossRef]
- Corbetta, M.; Patel, G.; Shulman, G.L. The reorienting system of the human brain: From environment to theory of mind. Neuron 2008, 58, 306–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehaene, S.; Pegado, F.; Braga, L.W.; Ventura, P.; Nunes Filho, G.; Jobert, A.; Dehaene-Lambertz, G.; Kolinsky, R.; Morais, J.; Cohen, L. How learning to read changes the cortical networks for vision and language. Science 2010, 330, 1359–1364. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.L. After Phrenology. Neural Reuse and the Interactive Brain; The MIT Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Anderson, M.L.; Kinnison, J.; Pessoa, L. Describing functional diversity of brain regions and brain networks. NeuroImage 2013, 73, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Poldrack, R.A. Can cognitive processes be inferred from neuroimaging data? Trends Cogn. Sci. 2006, 10, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Bassett, D.S.; Sporns, O. Network neuroscience. Nat. Neurosci. 2017, 20, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Petersen, S.E.; Sporns, O. Brain networks and cognitive architectures. Neuron 2015, 88, 207–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermundstad, A.M.; Bassett, D.S.; Brown, K.W.; Aminoff, E.M.; Clewett, D.; Freeman, S.; Frithsen, A.; Johnson, A.; Tipper, C.M.; Miller, M.B.; et al. Structural foundations of resting-state and task-based functional connectivity in the human brain. Proc. Natl. Acad. Sci. USA 2013, 110, 6169–6174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pessoa, L. Understanding brain networks and brain organization. Phys. Life Rev. 2014, 11, 400–435. [Google Scholar] [CrossRef] [Green Version]
- Bressler, S.L.; Menon, V. Large-scale brain networks in cognition: Emerging methods and principles. Trends Cogn. Sci. 2010, 14, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O.; Tononi, G.; Kotter, R. The human connectome: A structural description of the human brain. PLoS Comput. Biol. 2005, 1, e42. [Google Scholar] [CrossRef] [PubMed]
- Yeo, B.T.; Krienen, F.M.; Sepulcre, J.; Sabuncu, M.R.; Lashkari, D.; Hollinshead, M.; Roffman, J.L.; Smoller, J.W.; Zollei, L.; Polimeni, J.R.; et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J. Neurophysiol. 2011, 106, 1125–1165. [Google Scholar] [CrossRef] [PubMed]
- Modha, D.S.; Singh, R. Network architecture of the long-distance pathways in the macaque brain. Proc. Natl. Acad. Sci. USA 2010, 107, 13485–13490. [Google Scholar] [CrossRef] [Green Version]
- Power, J.D.; Schlaggar, B.L.; Lessov-Schlaggar, C.N.; Petersen, S.E. Evidence for hubs in human functional brain networks. Neuron 2013, 79, 798–813. [Google Scholar] [CrossRef] [Green Version]
- Buckner, R.L.; Sepulcre, J.; Talukdar, T.; Krienen, F.M.; Liu, H.; Hedden, T.; Andrews-Hanna, J.R.; Sperling, R.A.; Johnson, K.A. Cortical hubs revealed by intrinsic functional connectivity: Mapping, assessment of stability, and relation to Alzheimer’s disease. J. Neurosci. 2009, 29, 1860–1873. [Google Scholar] [CrossRef] [Green Version]
- Cole, M.W.; Reynolds, J.R.; Power, J.D.; Repovs, G.; Anticevic, A.; Braver, T.S. Multi-task connectivity reveals flexible hubs for adaptive task control. Nat. Neurosci. 2013, 16, 1348–1355. [Google Scholar] [CrossRef]
- Guggisberg, A.G.; Rizk, S.; Ptak, R.; Di Pietro, M.; Saj, A.; Lazeyras, F.; Lovblad, K.-O.; Schnider, A.; Pignat, J.-M. Two intrinstic coupling types for resting-state integration in the human brain. Brain Topogr. 2015, 28, 318–329. [Google Scholar] [CrossRef]
- van den Heuvel, M.P.; Hulshoff Pol, H.E. Exploring the brain network: A review on resting-state fMRI functional connectivity. Eur. Neuropsychopharmacol. 2010, 20, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E. Two views of brain function. Trends Cogn. Sci. 2010, 14, 180–190. [Google Scholar] [CrossRef]
- Gratton, C.; Laumann, T.O.; Nielsen, A.N.; Greene, D.J.; Gordon, E.M.; Gilmore, A.W.; Nelson, S.M.; Coalson, R.S.; Snyder, A.Z.; Schlaggar, B.L.; et al. Functional Brain Networks Are Dominated by Stable Group and Individual Factors, Not Cognitive or Daily Variation. Neuron 2018, 98, 439–452. [Google Scholar] [CrossRef] [Green Version]
- Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Corbetta, M.; Van Essen, D.C.; Raichle, M.E. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc. Natl. Acad. Sci. USA 2005, 102, 9673–9678. [Google Scholar] [CrossRef] [Green Version]
- Bola, M.; Sabel, B.A. Dynamic reorganization of brain functional networks during cognition. NeuroImage 2015, 114, 398–413. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.S.; Wymbs, N.F.; Porter, M.A.; Mucha, P.J.; Carlson, J.M.; Grafton, S.T. Dynamic reconfiguration of human brain networks during learning. Proc. Natl. Acad. Sci. USA 2011, 108, 7641–7646. [Google Scholar] [CrossRef] [Green Version]
- Dresler, M.; Shirer, W.R.; Konrad, B.N.; Muller, N.C.J.; Wagner, I.C.; Fernandez, G.; Czisch, M.; Greicius, M.D. Mnemonic Training Reshapes Brain Networks to Support Superior Memory. Neuron 2017, 93, 1227–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratton, C.; Nomura, E.M.; Perez, F.; D’Esposito, M. Focal brain lesions to critical locations cause widespread disruption of the modular organization of the brain. J. Cogn. Neurosci. 2012, 24, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- Alstott, J.; Breakspear, M.; Hagmann, P.; Cammoun, L.; Sporns, O. Modeling the impact of lesions in the human brain. PLoS Comput. Biol. 2009, 5, e1000408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, A.R.; Astafiev, S.V.; Lang, C.E.; Connor, L.T.; Rengachary, J.; Strube, M.J.; Pope, D.L.; Shulman, G.L.; Corbetta, M. Resting interhemispheric functional magnetic resonance imaging connectivity predicts performance after stroke. Ann. Neurol. 2010, 67, 365–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubovik, S.; Pignat, J.-M.; Ptak, R.; Aboulafia, T.; Allet, L.; Gillabert, N.; Magnin, C.; Albert, F.; Momjian-Mayor, I.; Nahum, L.; et al. The behavioral significance of coherent resting-state oscillations after stroke. NeuroImage 2012, 61, 249–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ptak, R.; Bourgeois, A.; Cavelti, S.; Doganci, N.; Schnider, A.; Iannotti, G.R. Discrete patterns of cross-hemispheric functional connectivity underlie impairments of spatial cognition after stroke. J. Neurosci. 2020, 40, 6638–6648. [Google Scholar] [CrossRef]
- Ptak, R.; Pedrazzini, E. Insular Cortex Mediates Attentional Capture by Behaviorally Relevant Stimuli after Damage to the Right Temporoparietal Junction. Cereb. Cortex 2021, 31, 4245–4258. [Google Scholar] [CrossRef]
- Gao, W.; Alcauter, S.; Smith, J.K.; Gilmore, J.H.; Lin, W. Development of human brain cortical network architecture during infancy. Brain Struct. Funct. 2015, 220, 1173–1186. [Google Scholar] [CrossRef] [PubMed]
- de Bie, H.M.; Boersma, M.; Adriaanse, S.; Veltman, D.J.; Wink, A.M.; Roosendaal, S.D.; Barkhof, F.; Stam, C.J.; Oostrom, K.J.; Delemarre-van de Waal, H.A.; et al. Resting-state networks in awake five- to eight-year old children. Hum. Brain Mapp. 2012, 33, 1189–1201. [Google Scholar] [CrossRef] [PubMed]
- Farrant, K.; Uddin, L.Q. Asymmetric development of dorsal and ventral attention networks in the human brain. Dev. Cogn. Neurosci. 2015, 12, 165–174. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, A.R. Towards a network theory of cognition. Neural Netw. 2000, 13, 861–870. [Google Scholar] [CrossRef]
- Caplan, D. On the Role of Group Studies in Neuropsychological and Pathopsychological Research. Cogn. Neuropsychol. 1988, 5, 535–548. [Google Scholar] [CrossRef]
- Shallice, T.; Cooper, R.P. The Organisation of Mind; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- McCloskey, M.; Caramazza, A. Theory and Methodology in Cognitive Neuropsychology: A Response to our Critics. Cogn. Neuropsychol. 1988, 5, 583–623. [Google Scholar] [CrossRef]
- Warrington, E.K.; Shallice, T. Category specific semantic impairments. Brain 1984, 107, 829–854. [Google Scholar] [CrossRef]
- Benson, D.F.; Greenberg, J.P. Visual form agnosia. Arch. Neurol. 1969, 20, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Campion, J.; Latto, R. Apperceptive agnosia due to carbon monoxide poisoning. An interpretation based on critical band masking from disseminated lesions. Behav. Brain Res. 1985, 15, 227–240. [Google Scholar] [CrossRef]
- Coltheart, M. Assumptions and Methods in Cognitive Neuropsychology. In The Handbook of Cognitive Neuropsychology; Rapp, B., Ed.; Taylor & Francis: Philadelphia, PA, USA, 2001; pp. 3–21. [Google Scholar]
- McCarthy, R.; Warrington, E.K. Cognitive Neuropsychology. A Clinical Introduction; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Rorden, C.; Karnath, H.-O. Using human brain lesions to infer function: A relic from a past era in the fMRI age? Nat. Rev. Neurosci. 2004, 5, 813–819. [Google Scholar] [CrossRef]
- Farah, M.J. The Cognitive Neuroscience of Vision; Blackwell: Malden, MA, USA, 2000. [Google Scholar]
- Price, C.J. The evolution of cognitive models: From neuropsychology to neuroimaging and back. Cortex 2018, 107, 37–49. [Google Scholar] [CrossRef]
- Gelb, A.; Goldstein, K. Zur Psychologie des optischen Wahrnehmungs- und Erkennungsvorganges. In Psychologische Analysen Hirnpathologischer Fälle; Gelb, A., Goldstein, K., Eds.; Johann Ambrosius Barth: Leipzig, Germany, 1920; Volume 1, pp. 1–142. [Google Scholar]
- Marr, D. Vision; Freeman and Co.: San Francisco, CA, USA, 1982. [Google Scholar]
- Riddoch, M.J.; Humphreys, G.W. A case of integrative visual agnosia. Brain 1987, 110, 1431–1462. [Google Scholar] [CrossRef]
- Riesenhuber, M.; Poggio, T. Hierarchical models of object recognition in cortex. Nat. Neurosci. 1999, 2, 1019–1025. [Google Scholar] [CrossRef]
- Riesenhuber, M.; Poggio, T. Models of object recognition. Nat. Neurosci. 2000, 3, 1199–1204. [Google Scholar] [CrossRef] [PubMed]
- Farah, M.J. Relations among the agnosias. In Case Studies in the Neuropsychology of Vision, Humphreys, G.W., Ed.; Psychology Press: Hove, UK, 1999; pp. 181–200. [Google Scholar]
- Ptak, R. Definition: Visual object agnosia. Cortex 2021, 143, 281. [Google Scholar] [CrossRef]
- Anaki, D.; Kaufman, Y.; Freedman, M.; Moscovitch, M. Associative (prosop)agnosia without (apparent) perceptual deficits: A case-study. Neuropsychologia 2007, 45, 1658–1671. [Google Scholar] [CrossRef]
- Fery, P.; Morais, J. A case study of visual agnosia without perceptual processing or structural descriptions impairment. Cogn. Neuropsychol. 2003, 20, 595–618. [Google Scholar] [CrossRef] [PubMed]
- Behrmann, M.; Kimchi, R. What does visual agnosia tell us about perceptual organization and its relationship to object perception? J. Exp. Psychol. Hum. Percept. Perf. 2003, 29, 19–42. [Google Scholar] [CrossRef]
- Humphreys, G.W.; Riddoch, M.J. Features, objects, action: The cognitive neuropsychology of visual object processing, 1984-2004. Cogn. Neuropsychol. 2006, 23, 156–183. [Google Scholar] [CrossRef] [PubMed]
- Rubens, A.B.; Benson, D.F. Associative visual agnosia. Arch. Neurol. 1971, 24, 305–316. [Google Scholar] [CrossRef]
- Farah, M.J. Visual Agnosia, 2nd ed.; MIT Press: Cambridge, MA, USA, 2004. [Google Scholar]
- Ptak, R.; Turri, F.; Doganci, N. Object Recognition and Visual Agnosia. In Encyclopedia of Behavioral Neuroscience, 2nd ed.; Elsevier: New York, NY, USA, 2022; pp. 587–596. [Google Scholar]
- Teuber, H. Alteration of perception and memory in man. In Analysis of Behavioral Change; Weiskrantz, L., Ed.; Harper & Row: New York, NY, USA, 1968. [Google Scholar]
- Farah, M.J. Patterns of co-occurrence among the associative agnosias: Implications for visual object representation. Cogn. Neuropsychol. 1991, 8, 1–19. [Google Scholar] [CrossRef]
- Feinberg, T.E.; Schindler, R.J.; Ochoa, E.; Kwan, P.C.; Farah, M.J. Associative visual agnosia and alexia without prosopagnosia. Cortex 1994, 30, 395–411. [Google Scholar] [CrossRef]
- Rice, G.E.; Kerry, S.J.; Robotham, R.J.; Leff, A.P.; Lambon Ralph, M.A.; Starrfelt, R. Category-selective deficits are the exception and not the rule: Evidence from a case-series of 64 patients with ventral occipito-temporal cortex damage. Cortex 2021, 138, 266–281. [Google Scholar] [CrossRef]
- Haxby, J.V.; Gobbini, M.I.; Furey, M.L.; Ishai, A.; Schouten, J.L.; Pietrini, P. Distributed and overlapping representations of faces and objects in ventral temporal cortex. Science 2001, 293, 2425–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishai, A.; Ungerleider, L.G.; Martin, A.; Schouten, J.L.; Haxby, J.V. Distributed representation of objects in the human ventral visual pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 9379–9384. [Google Scholar] [CrossRef] [Green Version]
- Denys, K.; Vanduffel, W.; Fize, D.; Nelissen, K.; Peuskens, H.; Van Essen, D.; Orban, G.A. The processing of visual shape in the cerebral cortex of human and nonhuman primates: A functional magnetic resonance imaging study. J. Neurosci. 2004, 24, 2551–2565. [Google Scholar] [CrossRef]
- Grill-Spector, K.; Kushnir, T.; Edelman, S.; Avidan, G.; Itzchak, Y.; Malach, R. Differential processing of objects under various viewing conditions in the human lateral occipital complex. Neuron 1999, 24, 187–203. [Google Scholar] [CrossRef] [Green Version]
- Malach, R.; Reppas, J.B.; Benson, R.R.; Kwong, K.K.; Jiang, H.; Kennedy, W.A.; Ledden, P.J.; Brady, T.J.; Rosen, B.R.; Tootell, R.B. Object-related activity revealed by functional magnetic resonance imaging in human occipital cortex. Proc. Natl. Acad. Sci. USA 1995, 92, 8135–8139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrmann, M.; Plaut, D.C. Distributed circuits, not circumscribed centers, mediate visual recognition. Trends Cogn. Sci. 2013, 17, 210–219. [Google Scholar] [CrossRef]
- Konen, C.S.; Behrmann, M.; Nishimura, M.; Kastner, S. The functional neuroanatomy of object agnosia: A case study. Neuron 2011, 71, 49–60. [Google Scholar] [CrossRef] [Green Version]
- James, T.W.; Culham, J.; Humphrey, G.K.; Milner, A.D.; Goodale, M.A. Ventral occipital lesions impair object recognition but not object-directed grasping: An fMRI study. Brain 2003, 126, 2463–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavina-Pratesi, C.; Kentridge, R.W.; Heywood, C.A.; Milner, A.D. Separate channels for processing form, texture, and color: Evidence from FMRI adaptation and visual object agnosia. Cereb. Cortex 2010, 20, 2319–2332. [Google Scholar] [CrossRef] [Green Version]
- Ptak, R.; Lazeyras, F.; Di Pietro, M.; Schnider, A.; Simon, S.R. Visual object agnosia is associated with a breakdown of object-selective responses in the lateral occipital cortex. Neuropsychologia 2014, 60, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Ptak, R.; Lazeyras, F. Functional connectivity and the failure to retrieve meaning from shape in visual object agnosia. Brain. Cogn. 2019, 131, 94–101. [Google Scholar] [CrossRef]
- Grush, R. The emulation theory of representation: Motor control, imagery, and perception. Behav. Brain Sci. 2004, 27, 377–396. [Google Scholar] [CrossRef] [Green Version]
- Shepard, R.N.; Metzler, J. Mental rotation of three-dimensional objects. Science 1971, 171, 701–703. [Google Scholar] [CrossRef] [Green Version]
- Searle, J.A.; Hamm, J.P. Mental rotation: An examination of assumptions. Wiley Interdiscip. Rev. Cogn. Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Parsons, L.M. Imagined spatial transformations of one’s hands and feet. Cogn. Psychol. 1987, 19, 178–241. [Google Scholar] [CrossRef]
- Wong, A.L.; Haith, A.M.; Krakauer, J.W. Motor Planning. Neuroscientist 2015, 21, 385–398. [Google Scholar] [CrossRef]
- Seidler, R.D.; Noll, D.C.; Thiers, G. Feedforward and feedback processes in motor control. NeuroImage 2004, 22, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Wolpert, D.M.; Diedrichsen, J.; Flanagan, J.R. Principles of sensorimotor learning. Nat. Rev. Neurosci. 2011, 12, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Haar, S.; Donchin, O. A Revised Computational Neuroanatomy for Motor Control. J. Cogn. Neurosci. 2020, 32, 1823–1836. [Google Scholar] [CrossRef]
- Franklin, D.W.; Wolpert, D.M. Computational mechanisms of sensorimotor control. Neuron 2011, 72, 425–442. [Google Scholar] [CrossRef] [Green Version]
- Shadmehr, R.; Krakauer, J.W. A computational neuroanatomy for motor control. Exp. Brain Res. 2008, 185, 359–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickering, M.J.; Clark, A. Getting ahead: Forward models and their place in cognitive architecture. Trends Cogn. Sci. 2014, 18, 451–456. [Google Scholar] [CrossRef]
- Wolpert, D.M.; Ghahramani, Z. Computational principles of movement neuroscience. Nat. Neurosci. 2000, 3 (Suppl. S3), 1212–1217. [Google Scholar] [CrossRef]
- Haar, S.; Dinstein, I.; Shelef, I.; Donchin, O. Effector-Invariant Movement Encoding in the Human Motor System. J. Neurosci. 2017, 37, 9054–9063. [Google Scholar] [CrossRef] [Green Version]
- Adolph, K.E.; Franchak, J.M. The development of motor behavior. Wiley Interdiscip. Rev. Cogn. Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarty, M.E.; Clifton, R.K.; Ashmead, D.H.; Lee, P.; Goubet, N. How infants use vision for grasping objects. Child. Dev. 2001, 72, 973–987. [Google Scholar] [CrossRef] [PubMed]
- Frick, A.; Daum, M.M.; Walser, S.; Mast, F.W. Motor processes in children’s mental rotation. J. Cogn. Dev. 2009, 10, 18–40. [Google Scholar] [CrossRef]
- Frick, A.; Hansen, M.A.; Newcombe, N.S. Development of mental rotation in 3- to 5-year-old children. Cogn. Dev. 2013, 28, 386–399. [Google Scholar] [CrossRef] [Green Version]
- Sekiyama, K. Kinesthetic aspects of mental representations in the identification of left and right hands. Percept. Psychophys. 1982, 32, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Coslett, H.B.; Medina, J.; Kliot, D.; Burkey, A.R. Mental motor imagery indexes pain: The hand laterality task. Eur. J. Pain 2010, 14, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- de Lange, F.P.; Helmich, R.C.; Toni, I. Posture influences motor imagery: An fMRI study. NeuroImage 2006, 33, 609–617. [Google Scholar] [CrossRef]
- Hardwick, R.M.; Caspers, S.; Eickhoff, S.B.; Swinnen, S.P. Neural correlates of action: Comparing meta-analyses of imagery, observation, and execution. Neurosci. Biobehav. Rev. 2018, 94, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Grezes, J.; Decety, J. Functional anatomy of execution, mental simulation, observation, and verb generation of actions: A meta-analysis. Hum. Brain Mapp. 2001, 12, 1–19. [Google Scholar] [CrossRef]
- Hanakawa, T.; Dimyan, M.A.; Hallett, M. Motor planning, imagery, and execution in the distributed motor network: A time-course study with functional MRI. Cereb. Cortex 2008, 18, 2775–2788. [Google Scholar] [CrossRef] [Green Version]
- Hetu, S.; Gregoire, M.; Saimpont, A.; Coll, M.P.; Eugene, F.; Michon, P.E.; Jackson, P.L. The neural network of motor imagery: An ALE meta-analysis. Neurosci. Biobehav. Rev. 2013, 37, 930–949. [Google Scholar] [CrossRef]
- Zacks, J.M. Neuroimaging studies of mental rotation: A meta-analysis and review. J. Cogn. Neurosci. 2008, 20, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Cona, G.; Scarpazza, C. Where is the “where” in the brain? A meta-analysis of neuroimaging studies on spatial cognition. Hum. Brain Mapp. 2019, 40, 1867–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ptak, R.; Schnider, A.; Fellrath, J. The Dorsal Frontoparietal Network: A Core System for Emulated Action. Trends Cogn. Sci. 2017, 21, 589–599. [Google Scholar] [CrossRef]
- Kosslyn, S.M.; DiGirolamo, G.J.; Thompson, W.L.; Alpert, N.M. Mental rotation of objects versus hands: Neural mechanisms revealed by positron emission tomography. Psychophysiology 1998, 35, 151–161. [Google Scholar] [CrossRef]
- Jordan, K.; Wustenberg, T.; Heinze, H.J.; Peters, M.; Jancke, L. Women and men exhibit different cortical activation patterns during mental rotation tasks. Neuropsychologia 2002, 40, 2397–2408. [Google Scholar] [CrossRef]
- Milivojevic, B.; Hamm, J.P.; Corballis, M.C. Functional neuroanatomy of mental rotation. J. Cogn. Neurosci. 2009, 21, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Lamm, C.; Windischberger, C.; Moser, E.; Bauer, H. The functional role of dorso-lateral premotor cortex during mental rotation: An event-related fMRI study separating cognitive processing steps using a novel task paradigm. NeuroImage 2007, 36, 1374–1386. [Google Scholar] [CrossRef] [PubMed]
- Tomasino, B.; Gremese, M. Effects of Stimulus Type and Strategy on Mental Rotation Network: An Activation Likelihood Estimation Meta-Analysis. Front. Hum. Neurosci. 2015, 9, 693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ditunno, P.L.; Mann, V.A. Right hemisphere specialization for mental rotation in normals and brain damaged subjects. Cortex 1990, 26, 177–188. [Google Scholar] [CrossRef]
- Ratcliff, G. Spatial thought, mental rotation and the right cerebral hemisphere. Neuropsychologia 1979, 17, 49–54. [Google Scholar] [CrossRef]
- Tomasino, B.; Toraldo, A.; Rumiati, R.I. Dissociation between the mental rotation of visual images and motor images in unilateral brain-damaged patients. Brain. Cogn. 2003, 51, 368–371. [Google Scholar] [CrossRef]
- Tomasino, B.; Rumiati, R.I. Effects of strategies on mental rotation and hemispheric lateralization: Neuropsychological evidence. J. Cogn. Neurosci. 2004, 16, 878–888. [Google Scholar] [CrossRef]
- Buiatti, T.; Mussoni, A.; Toraldo, A.; Skrap, M.; Shallice, T. Two qualitatively different impairments in making rotation operations. Cortex 2011, 47, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Cona, G.; Panozzo, G.; Semenza, C. The role of dorsal premotor cortex in mental rotation: A transcranial magnetic stimulation study. Brain. Cogn. 2017, 116, 71–78. [Google Scholar] [CrossRef]
- Wohlschlager, A.; Wohlschlager, A. Mental and manual rotation. J. Exp. Psychol. Hum. Percept. Perform. 1998, 24, 397–412. [Google Scholar] [CrossRef]
- Wexler, M.; Kosslyn, S.M.; Berthoz, A. Motor processes in mental rotation. Cognition 1998, 68, 77–94. [Google Scholar] [CrossRef] [Green Version]
- Wohlschlager, A. Mental object rotation and the planning of hand movements. Percept. Psychophys. 2001, 63, 709–718. [Google Scholar] [CrossRef]
- Nico, D.; Daprati, E.; Rigal, F.; Parsons, L.; Sirigu, A. Left and right hand recognition in upper limb amputees. Brain 2004, 127, 120–132. [Google Scholar] [CrossRef] [Green Version]
- Menendez Granda, M.; Iannotti, G.R.; Darque, A.; Ptak, R. Does mental rotation emulate motor processes? An electrophysiological study of objects and body parts. J. Prepr. 2021. [Google Scholar] [CrossRef]
- Thayer, Z.C.; Johnson, B.W. Cerebral processes during visuo-motor imagery of hands. Psychophysiology 2006, 43, 401–412. [Google Scholar] [CrossRef]
- Jansen, P.; Render, A.; Scheer, C.; Siebertz, M. Mental rotation with abstract and embodied objects as stimuli: Evidence from event-related potential (ERP). Exp. Brain Res. 2020, 238, 525–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehaene, S.; Dehaene-Lambertz, G.; Cohen, L. Abstract representation of numbers in the animal and human brain. Trends Neurosci. 1998, 21, 355–361. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Serio, G.; Battaglia, S. Please, don’t do it! Fifteen years of progress of non-invasive brain stimulation in action inhibition. Cortex 2020, 132, 404–422. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Serio, G.; Scarpazza, C.; D’Ausilio, A.; Borgomaneri, S. Frozen in (e)motion: How reactive motor inhibition is influenced by the emotional content of stimuli in healthy and psychiatric populations. Behav. Res. 2021, 146, 103963. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Vitale, F.; Battaglia, S.; Avenanti, A. Early Right Motor Cortex Response to Happy and Fearful Facial Expressions: A TMS Motor-Evoked Potential Study. Brain Sci. 2021, 11, 1203. [Google Scholar] [CrossRef]
- Eichenbaum, H.; Cohen, N.J. Can we reconcile the declarative memory and spatial navigation views on hippocampal function? Neuron 2014, 83, 764–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aisenberg, D.; Henik, A. Reuse or re-function? Behav. Brain Sci. 2010, 33, 266–267. [Google Scholar] [CrossRef]
- Goldinger, S.D.; Papesh, M.H.; Barnhart, A.S.; Hansen, W.A.; Hout, M.C. The poverty of embodied cognition. Psychon Bull. Rev. 2016, 23, 959–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churchland, P.M. Matter and Consciousness; The MIT Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Anderson, B. There is no such thing as attention. Front. Psychol. 2011, 2, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hommel, B.; Chapman, C.S.; Cisek, P.; Neyedli, H.F.; Song, J.H.; Welsh, T.N. No one knows what attention is. Atten Percept. Psychophys 2019, 81, 2288–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genon, S.; Reid, A.; Langner, R.; Amunts, K.; Eickhoff, S.B. How to Characterize the Function of a Brain Region. Trends Cogn. Sci. 2018, 22, 350–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.L. Mining the brain for a new taxonomy of the mind. Philos. Compass 2015, 10, 68–77. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ptak, R.; Doganci, N.; Bourgeois, A. From Action to Cognition: Neural Reuse, Network Theory and the Emergence of Higher Cognitive Functions. Brain Sci. 2021, 11, 1652. https://doi.org/10.3390/brainsci11121652
Ptak R, Doganci N, Bourgeois A. From Action to Cognition: Neural Reuse, Network Theory and the Emergence of Higher Cognitive Functions. Brain Sciences. 2021; 11(12):1652. https://doi.org/10.3390/brainsci11121652
Chicago/Turabian StylePtak, Radek, Naz Doganci, and Alexia Bourgeois. 2021. "From Action to Cognition: Neural Reuse, Network Theory and the Emergence of Higher Cognitive Functions" Brain Sciences 11, no. 12: 1652. https://doi.org/10.3390/brainsci11121652