
Facilitation of Motor Evoked Potentials in Response to a Modified 30 Hz Intermittent Theta-Burst Stimulation Protocol in Healthy Adults


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Participants
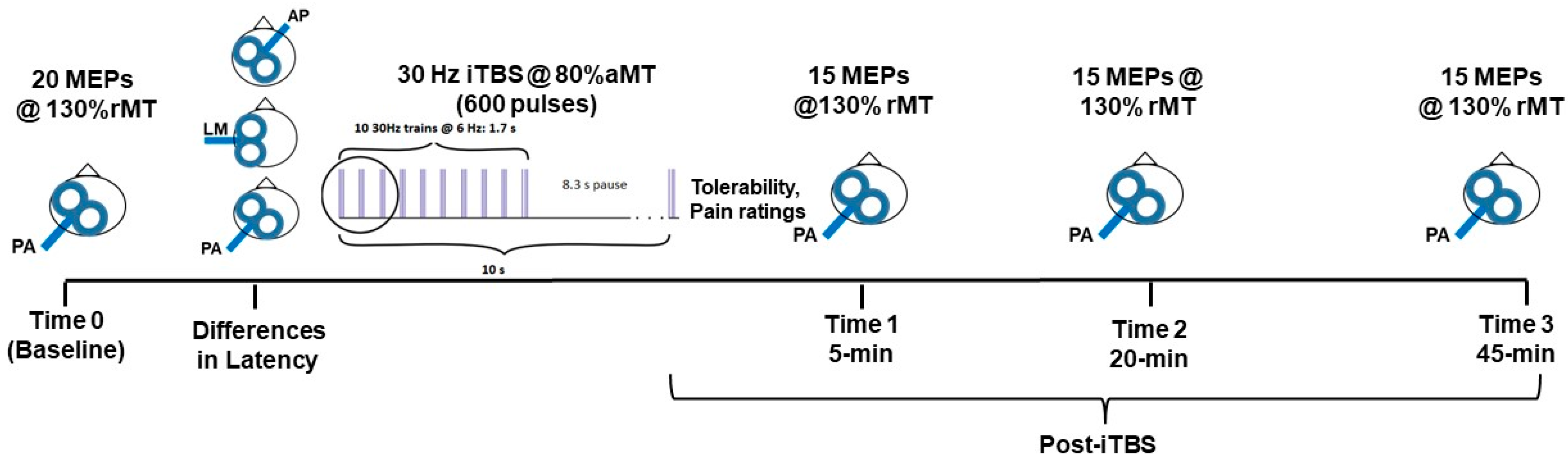
2.2. Experimental Protocol
2.3. Baseline Assessment of Corticospinal Excitability
2.4. Assessment of MEP Latency with Different Coil Orientations
2.5. Modified 30 Hz/6 Hz iTBS Protocol
2.6. Post-iTBS Changes in Corticospinal Excitability, Safety and Tolerability
2.7. Analysis of MEP Data
2.8. Analysis of Responses to iTBS
2.9. Statistical Analysis
3. Results
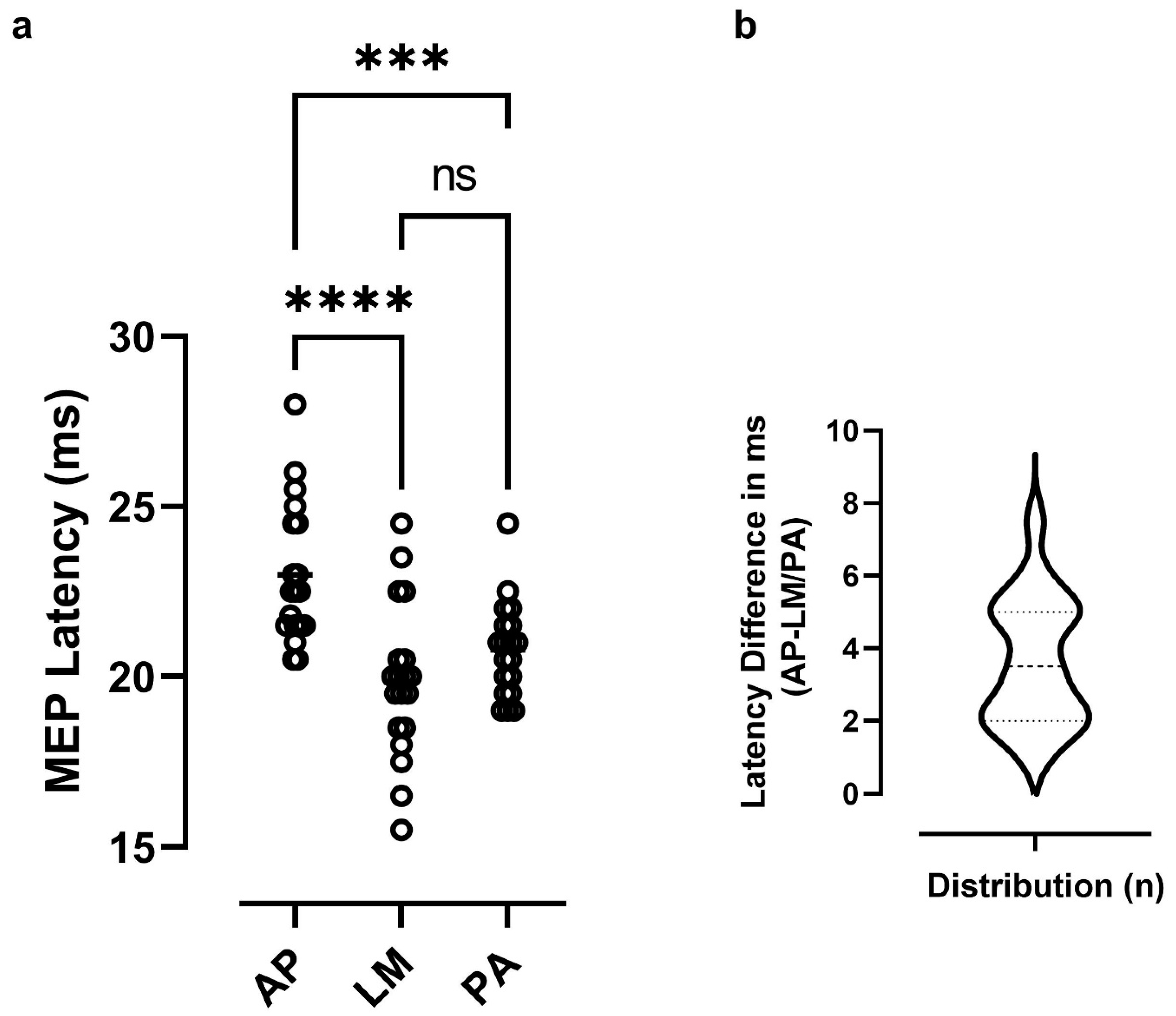
3.1. Baseline Measures of Excitability and Latency Differences
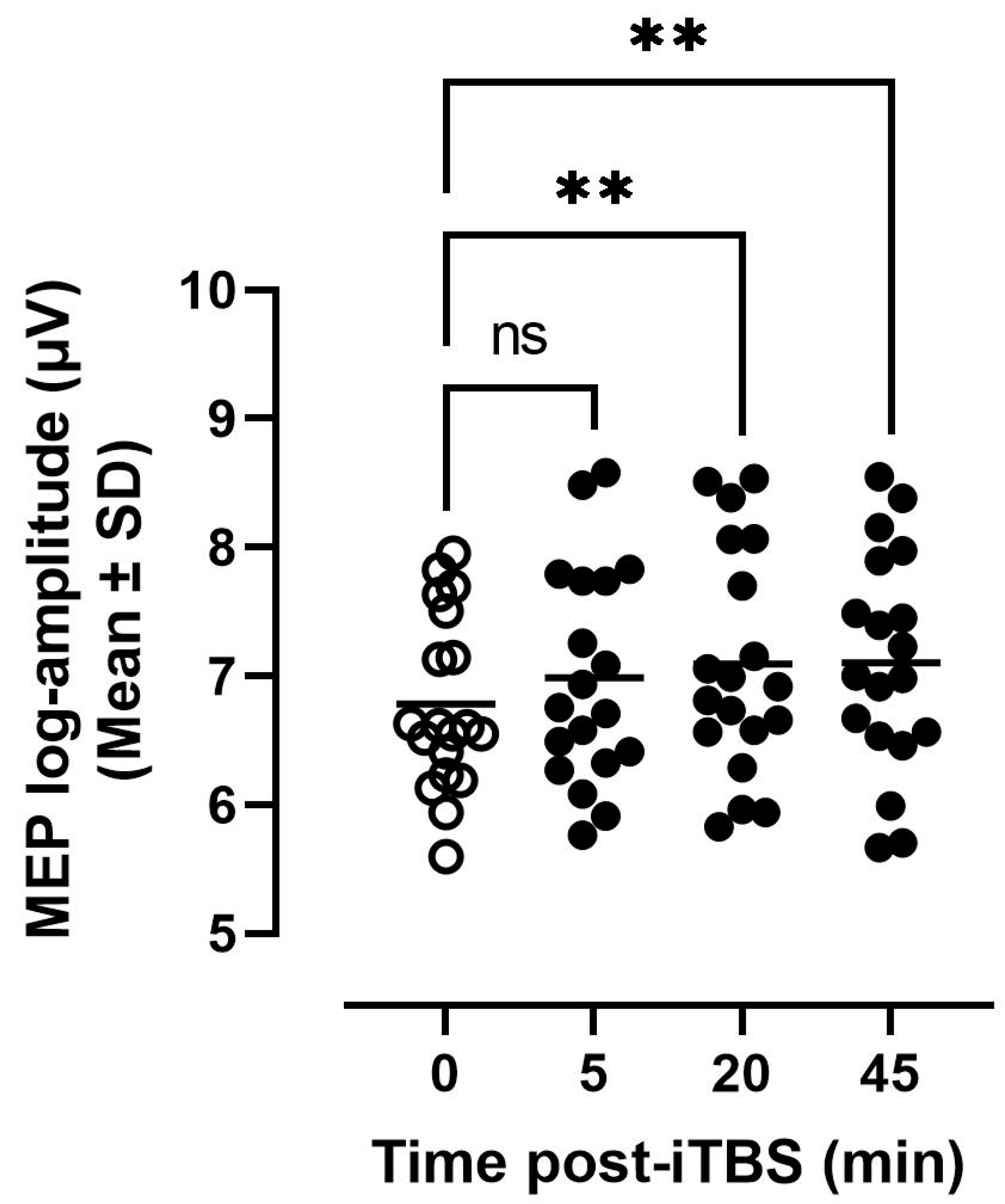
3.2. Tolerability and MEP Modulation in Response to iTBS
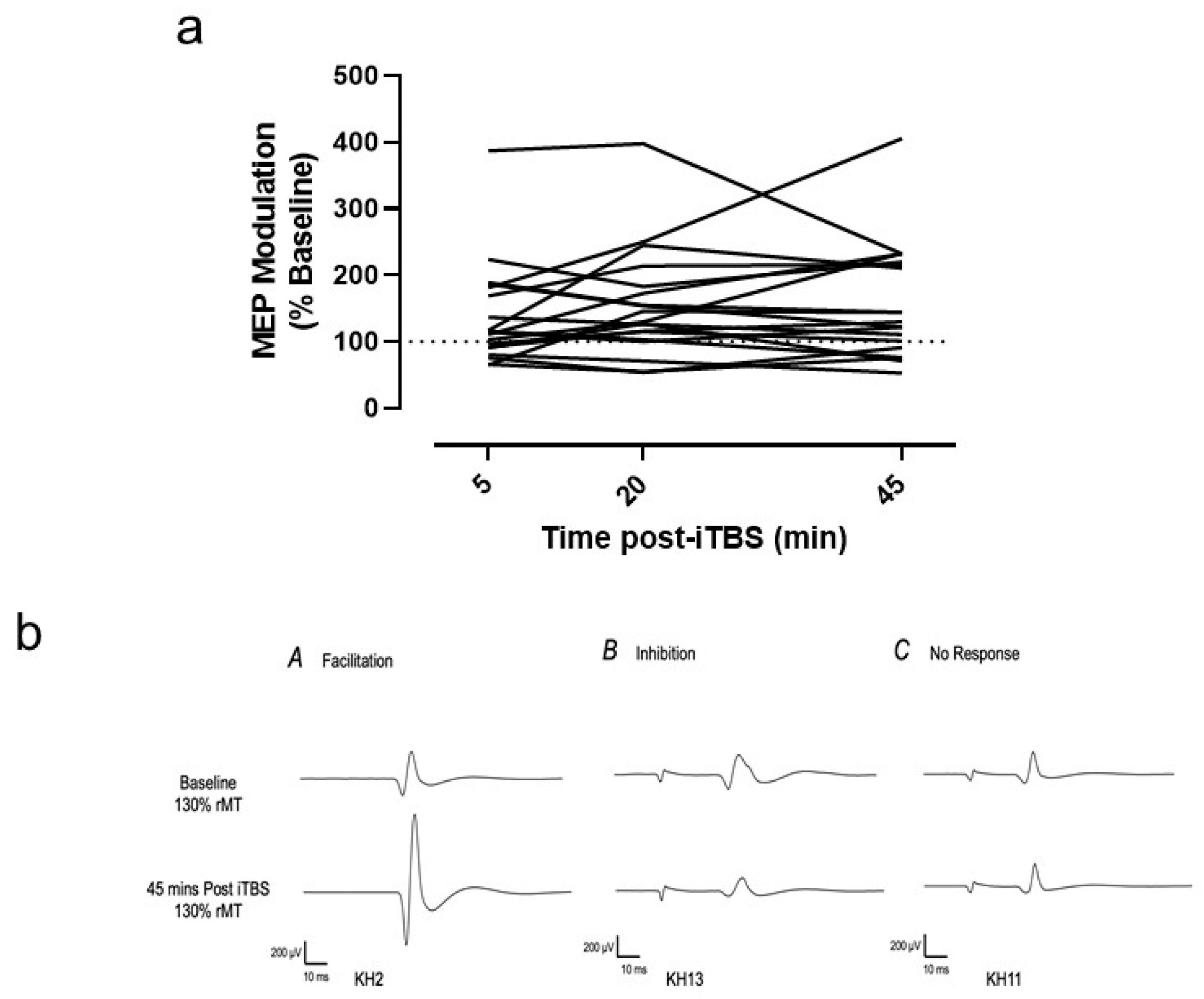
3.3. Variability of Individual Responses
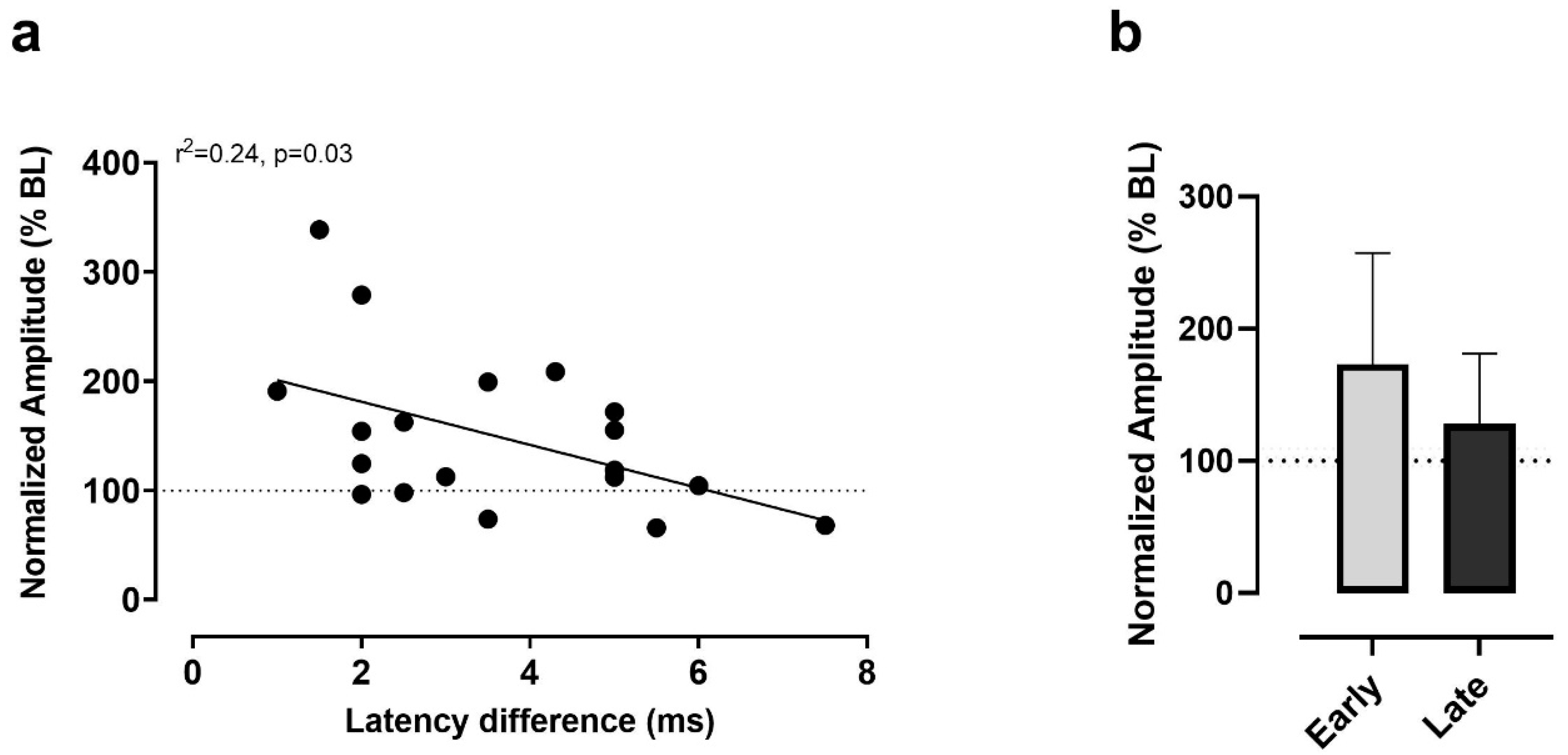
3.4. Latency Differences as Predictors of Responses to iTBS
4. Discussion
4.1. Corticospinal Excitability and Latency Differences at Baseline
4.2. Tolerability, MEP Modulation and Variability of Responses to 30 Hz iTBS
4.3. Predictors of Responses to iTBS from Latency Differences
4.4. Study Limitations
4.5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, Y.-Z.; Edwards, M.J.; Rounis, E.; Bhatia, K.P.; Rothwell, J.C. Theta Burst Stimulation of the Human Motor Cortex. Neuron 2005, 45, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Hess, G.; Aizenman, C.D.; Donoghue, J.P. Conditions for the induction of long-term potentiation in layer II/III horizontal connections of the rat motor cortex. J. Neurophysiol. 1996, 75, 1765–1778. [Google Scholar] [CrossRef]
- Chung, S.W.; Hill, A.T.; Rogasch, N.C.; Hoy, K.; Fitzgerald, P. Use of theta-burst stimulation in changing excitability of motor cortex: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2016, 63, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Suppa, A.; Huang, Y.-Z.; Funke, K.; Ridding, M.C.; Cheeran, B.; Di Lazzaro, V.; Ziemann, U.; Rothwell, J.C. Ten Years of Theta Burst Stimulation in Humans: Established Knowledge, Unknowns and Prospects. Brain Stimul. 2016, 9, 323–335. [Google Scholar] [CrossRef]
- Talelli, P.; Greenwood, R.; Rothwell, J. Exploring Theta Burst Stimulation as an intervention to improve motor recovery in chronic stroke. Clin. Neurophysiol. 2007, 118, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Daskalakis, Z.J. Theta-burst transcranial magnetic stimulation in depression: When less may be more. Brain 2014, 137, 1860–1862. [Google Scholar] [CrossRef] [Green Version]
- Konstantinou, G.N.; Downar, J.; Daskalakis, Z.J.; Blumberger, D.M. Accelerated Intermittent Theta Burst Stimulation in Late-Life Depression: A Possible Option for Older Depressed Adults in Need of ECT During the COVID-19 Pandemic. Am. J. Geriatr. Psychiatry 2020, 28, 1025–1029. [Google Scholar] [CrossRef]
- Corp, D.T.; Bereznicki, H.G.K.; Clark, G.M.; Youssef, G.J.; Fried, P.J.; Jannati, A.; Davies, C.B.; Gomes-Osman, J.; Stamm, J.; Chung, S.W.; et al. Large-Scale Analysis of Interin-dividual Variability in Theta-Burst Stimulation Data: Results from the ‘Big Tms Data Collaboration’. Brain Stimul. 2020, 13, 1476–1488. [Google Scholar] [CrossRef]
- Hinder, M.R.; Goss, E.L.; Fujiyama, H.; Canty, A.; Garry, M.; Rodger, J.; Summers, J.J. Inter- and Intra-individual Variability Following Intermittent Theta Burst Stimulation: Implications for Rehabilitation and Recovery. Brain Stimul. 2014, 7, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Goldsworthy, M.; Pitcher, J.; Ridding, M.C. A comparison of two different continuous theta burst stimulation paradigms applied to the human primary motor cortex. Clin. Neurophysiol. 2012, 123, 2256–2263. [Google Scholar] [CrossRef]
- Nyffeler, T.; Cazzoli, D.; Wurtz, P.; Luthi, M.; von Wartburg, R.; Chaves, S.; Deruaz, A.; Hess, C.W.; Muri, R.M. Ne-glect-Like Visual Exploration Behaviour after Theta Burst Transcranial Magnetic Stimulation of the Right Posterior Parietal Cortex. Eur. J. Neurosci. 2008, 27, 1809–1813. [Google Scholar] [CrossRef]
- Jacobs, M.F.; Tsang, P.; Lee, K.G.; Asmussen, M.J.; Zapallow, C.M.; Nelson, A.J. 30 Hz Theta-burst Stimulation Over Primary Somatosensory Cortex Modulates Corticospinal Output to the Hand. Brain Stimul. 2014, 7, 269–274. [Google Scholar] [CrossRef]
- Tsang, P.; Jacobs, M.F.; Lee, K.G.H.; Asmussen, M.J.; Zapallow, C.M.; Nelson, A.J. Continuous Theta-Burst Stimu-lation over Primary Somatosensory Cortex Modulates Short-Latency Afferent Inhibition. Clin. Neurophysiol. 2014, 125, 2253–2259. [Google Scholar] [CrossRef]
- Wu, S.W.; Shahana, N.; Huddleston, D.A.; Gilbert, D.L. Effects of 30hz Theta Burst Transcranial Magnetic Stimu-lation on the Primary Motor Cortex. J. Neurosci. Methods 2012, 208, 161–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedapati, E.V.; Gilbert, D.L.; Horn, P.S.; Huddleston, D.A.; Laue, C.S.; Shahana, N.; Wu, S.W. Effect of 30 Hz theta burst transcranial magnetic stimulation on the primary motor cortex in children and adolescents. Front. Hum. Neurosci. 2015, 9, 91. [Google Scholar] [CrossRef] [Green Version]
- Hamada, M.; Murase, N.; Hasan, A.; Balaratnam, M.; Rothwell, J.C. The Role of Interneuron Networks in Driving Human Motor Cortical Plasticity. Cereb. Cortex 2013, 23, 1593–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borckardt, J.J.; Nahas, Z.; Koola, J.; George, M.S. Estimating Resting Motor Thresholds in Transcranial Magnetic Stimulation Research and Practice: A Computer Simulation Evaluation of Best Methods. J. ECT 2006, 22, 169–175. [Google Scholar] [CrossRef]
- Brown, K.E.; Lohse, K.R.; Mayer, I.M.S.; Strigaro, G.; Desikan, M.; Casula, E.P.; Meunier, S.; Popa, T.; Lamy, J.-C.; Odish, O.; et al. The reliability of commonly used electrophysiology measures. Brain Stimul. 2017, 10, 1102–1111. [Google Scholar] [CrossRef] [PubMed]
- Oberman, L.; Edwards, D.; Eldaief, M.; Pascual-Leone, A. Safety of Theta Burst Transcranial Magnetic Stimulation: A Systematic Review of the Literature. J. Clin. Neurophysiol. 2011, 28, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Perellón-Alfonso, R.; Kralik, M.; Pileckyte, I.; Princic, M.; Bon, J.; Matzhold, C.; Fischer, B.; Šlahorová, P.; Pirtošek, Z.; Rothwell, J.; et al. Similar effect of intermittent theta burst and sham stimulation on corticospinal excitability: A 5-day repeated sessions study. Eur. J. Neurosci. 2018, 48, 1990–2000. [Google Scholar] [CrossRef]
- Nielsen, J.F. Logarithmic Distribution of Amplitudes of Compound Muscle Action Potentials Evoked by Transcranial Magnetic Stimulation. J. Clin. Neurophysiol. 1996, 13, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, M.; Zoghi, M.; Jaberzadeh, S. Biological and anatomical factors influencing interindividual variability to noninvasive brain stimulation of the primary motor cortex: A systematic review and meta-analysis. Rev. Neurosci. 2017, 29, 199–222. [Google Scholar] [CrossRef]
- Hordacre, B.; Goldsworthy, M.; Vallence, A.-M.; Darvishi, S.; Moezzi, B.; Hamada, M.; Rothwell, J.C.; Ridding, M.C. Variability in neural excitability and plasticity induction in the human cortex: A brain stimulation study. Brain Stimul. 2017, 10, 588–595. [Google Scholar] [CrossRef] [Green Version]
- Van Dam, J.; Goldsworthy, M.; Hague, W.; Coat, S.; Pitcher, J. Cortical Plasticity and Interneuron Recruitment in Adolescents Born to Women with Gestational Diabetes Mellitus. Brain Sci. 2021, 11, 388. [Google Scholar] [CrossRef] [PubMed]
- Davidson, T.W.; Bolić, M.; Tremblay, F. Predicting Modulation in Corticomotor Excitability and in Transcallosal Inhibition in Response to Anodal Transcranial Direct Current Stimulation. Front. Hum. Neurosci. 2016, 10, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashihara, M.; Bos, M.A.V.D.; Menon, P.; Kiernan, M.C.; Vucic, S. Interneuronal networks mediate cortical inhibition and facilitation. Clin. Neurophysiol. 2020, 131, 1000–1010. [Google Scholar] [CrossRef] [PubMed]
- Di Lazzaro, V.; Rothwell, J.; Capogna, M. Noninvasive Stimulation of the Human Brain: Activation of Multiple Cortical Circuits. Neuroscientist 2017, 24, 246–260. [Google Scholar] [CrossRef] [PubMed]
- Gillick, B.T.; Rich, T.; Chen, M.; Meekins, G.D. Case report of vasovagal syncope associated with single pulse transcranial magnetic stimulation in a healthy adult participant. BMC Neurol. 2015, 15, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malm, E.; Struckmann, W.; Persson, J.; Bodén, R. Pain trajectories of dorsomedial prefrontal intermittent theta burst stimulation versus sham treatment in depression. BMC Neurol. 2020, 20, 311. [Google Scholar] [CrossRef] [PubMed]
- Wischnewski, M.; Schutter, D.J. Efficacy and time course of paired associative stimulation in cortical plasticity: Implications for neuropsychiatry. Clin. Neurophysiol. 2016, 127, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Goldsworthy, M.R.; Hordacre, B.; Rothwell, J.C.; Ridding, M.C. Effects of Rtms on the Brain: Is There Value in Variability? Cortex 2021, 139, 43–59. [Google Scholar] [CrossRef]
- Guerra, A.; Lopez-Alonso, V.; Cheeran, B.; Suppa, A. Variability in non-invasive brain stimulation studies: Reasons and results. Neurosci. Lett. 2020, 719, 133330. [Google Scholar] [CrossRef] [Green Version]
- McCalley, D.M.; Lench, D.H.; Doolittle, J.D.; Imperatore, J.P.; Hoffman, M.; Hanlon, C.A. Determining the optimal pulse number for theta burst induced change in cortical excitability. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Hirsch, J.C.; Crepel, F. Use-dependent changes in synaptic efficacy in rat prefrontal neurons in vitro. J. Physiol. 1990, 427, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wong, T.P.; Pozza, M.F.; Lingenhoehl, K.; Wang, Y.; Sheng, M.; Auberson, Y.P.; Wang, Y.T. Role of NMDA Receptor Subtypes in Governing the Direction of Hippocampal Synaptic Plasticity. Science 2004, 304, 1021–1024. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, M.; Hong, K.; Mikoshiba, K.; Poo, M.-M.; Kato, K. Calcium stores regulate the polarity and input specificity of synaptic modification. Nat. Cell Biol. 2000, 408, 584–588. [Google Scholar] [CrossRef]
- Shen, K.-Z.; Zhu, Z.-T.; Munhall, A.; Johnson, S.W. Synaptic plasticity in rat subthalamic nucleus induced by high-frequency stimulation. Synapse 2003, 50, 314–319. [Google Scholar] [CrossRef]
- Guerra, A.; Pogosyan, A.; Nowak, M.; Tan, H.; Ferreri, F.; Di Lazzaro, V.; Brown, P. Phase Dependency of the Human Primary Motor Cortex and Cholinergic Inhibition Cancelation During Beta tACS. Cereb. Cortex 2016, 26, 3977–3990. [Google Scholar] [CrossRef] [Green Version]
- Ferreri, F.; Vecchio, F.; Guerra, A.; Miraglia, F.; Ponzo, D.; Vollero, L.; Iannello, G.; Maatta, S.; Mervaala, E.; Rossini, P.M.; et al. Age related differences in functional synchronization of EEG activity as evaluated by means of TMS-EEG coregistrations. Neurosci. Lett. 2017, 647, 141–146. [Google Scholar] [CrossRef]
- Bergmann, T.O.; Mölle, M.; Schmidt, M.A.; Lindner, M.A.; Marshall, L.; Born, J.; Siebner, H.R. EEG-Guided Transcranial Magnetic Stimulation Reveals Rapid Shifts in Motor Cortical Excitability during the Human Sleep Slow Oscillation. J. Neurosci. 2012, 32, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keil, J.; Timm, J.; Sanmiguel, I.; Schulz, H.; Obleser, J.; Schonwiesner, M. Cortical Brain States and Corticospinal Syn-chronization Influence Tms-Evoked Motor Potentials. J. Neurophysiol. 2014, 111, 513–519. [Google Scholar] [CrossRef]
- Rocchi, L.; Ibáñez, J.; Benussi, A.; Hannah, R.; Rawji, V.; Casula, E.; Rothwell, J. Variability and Predictors of Response to Continuous Theta Burst Stimulation: A TMS-EEG Study. Front. Neurosci. 2018, 12, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lazzaro, V.; Pilato, F.; Saturno, E.; Oliviero, A.; Dileone, M.; Mazzone, P.; Insola, A.; Tonali, P.A.; Ranieri, F.; Huang, Y.Z.; et al. Theta-burst repetitive transcranial magnetic stimulation suppresses specific excitatory circuits in the human motor cortex. J. Physiol. 2005, 565, 945–950. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Insola, A.; Mazzone, P.; Tonali, P.; Rothwell, J.; Profice, P.; Oliviero, A. Direct demonstration of interhemispheric inhibition of the human motor cortex produced by transcranial magnetic stimulation. Exp. Brain Res. 1999, 124, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-Z.; Chen, R.-S.; Rothwell, J.; Wen, H.-Y. The after-effect of human theta burst stimulation is NMDA receptor dependent. Clin. Neurophysiol. 2007, 118, 1028–1032. [Google Scholar] [CrossRef] [PubMed]
- Krause, B.; Kadosh, R.C. Not All Brains Are Created Equal: The Relevance of Individual Differences in Re-sponsiveness to Transcranial Electrical Stimulation. Front. Syst. Neurosci. 2014, 8, 25. [Google Scholar] [CrossRef] [PubMed]
- Krause, B.; Márquez-Ruiz, J.; Kadosh, R.C. The effect of transcranial direct current stimulation: A role for cortical excitation/inhibition balance? Front. Hum. Neurosci. 2013, 7, 602. [Google Scholar] [CrossRef] [Green Version]
- Cheeran, B.; Talelli, P.; Mori, F.; Koch, G.; Suppa, A.; Edwards, M.; Houlden, H.; Bhatia, K.; Greenwood, R.; Rothwell, J. A common polymorphism in the brain-derived neurotrophic factor gene (BDNF) modulates human cortical plasticity and the response to rTMS. J. Physiol. 2008, 586, 5717–5725. [Google Scholar] [CrossRef]
- Cheeran, B.; Ritter, C.; Rothwell, J.; Siebner, H. Mapping genetic influences on the corticospinal motor system in humans. Neuroscience 2009, 164, 156–163. [Google Scholar] [CrossRef]
- Antal, A.; Terney, D.; Kühnl, S.; Paulus, W. Anodal Transcranial Direct Current Stimulation of the Motor Cortex Ameliorates Chronic Pain and Reduces Short Intracortical Inhibition. J. Pain Symptom Manag. 2010, 39, 890–903. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, J.; Hughes, J.; Ridding, M.; Thomas, P.Q.; Semmler, J.G. Differential modulation of motor cortex excitability in BDNF Met allele carriers following experimentally induced and use-dependent plasticity. Eur. J. Neurosci. 2012, 36, 2640–2649. [Google Scholar] [CrossRef]
- Lee, M.; Kim, S.E.; Kim, W.S.; Lee, J.; Yoo, H.K.; Park, K.-D.; Choi, K.-G.; Jeong, S.-Y.; Kim, B.G.; Lee, H.W. Interaction of Motor Training and Intermittent Theta Burst Stimulation in Modulating Motor Cortical Plasticity: Influence of Bdnf Val66met Polymor-phism. PLoS ONE 2013, 8, e57690. [Google Scholar] [CrossRef]
- Chang, W.H.; Bang, O.Y.; Shin, Y.-I.; Lee, A.; Pascual-Leone, A.; Kim, Y.-H. Bdnf Polymor-phism and Differential Rtms Effects on Motor Recovery of Stroke Patients. Brain Stimul. 2014, 7, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Di Lazzaro, V.; Pellegrino, G.; Di Pino, G.; Corbetto, M.; Ranieri, F.; Brunelli, N.; Paolucci, M.; Bucossi, S.; Ventriglia, M.C.; Brown, P.; et al. Val66Met BDNF Gene Polymorphism Influences Human Motor Cortex Plasticity in Acute Stroke. Brain Stimul. 2015, 8, 92–96. [Google Scholar] [CrossRef] [Green Version]
- A Kleim, J.; Chan, S.; Pringle, E.; Schallert, K.; Procaccio, V.; Jimenez, R.; Cramer, S.C. BDNF val66met polymorphism is associated with modified experience-dependent plasticity in human motor cortex. Nat. Neurosci. 2006, 9, 735–737. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosel, K.; Tremblay, F. Facilitation of Motor Evoked Potentials in Response to a Modified 30 Hz Intermittent Theta-Burst Stimulation Protocol in Healthy Adults. Brain Sci. 2021, 11, 1640. https://doi.org/10.3390/brainsci11121640
Hosel K, Tremblay F. Facilitation of Motor Evoked Potentials in Response to a Modified 30 Hz Intermittent Theta-Burst Stimulation Protocol in Healthy Adults. Brain Sciences. 2021; 11(12):1640. https://doi.org/10.3390/brainsci11121640
Chicago/Turabian StyleHosel, Katarina, and François Tremblay. 2021. "Facilitation of Motor Evoked Potentials in Response to a Modified 30 Hz Intermittent Theta-Burst Stimulation Protocol in Healthy Adults" Brain Sciences 11, no. 12: 1640. https://doi.org/10.3390/brainsci11121640