
Neuroprotective Effect of Clobenpropit against Lipopolysaccharide-Induced Cognitive Deficits via Attenuating Neuroinflammation and Enhancing Mitochondrial Functions in Mice

 ,
,  , , ,
, , ,
Abstract
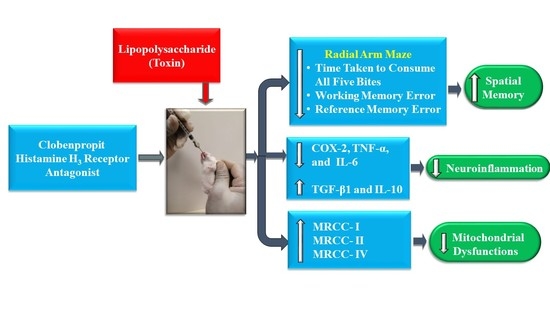
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Drugs and Chemicals
2.2. Animals
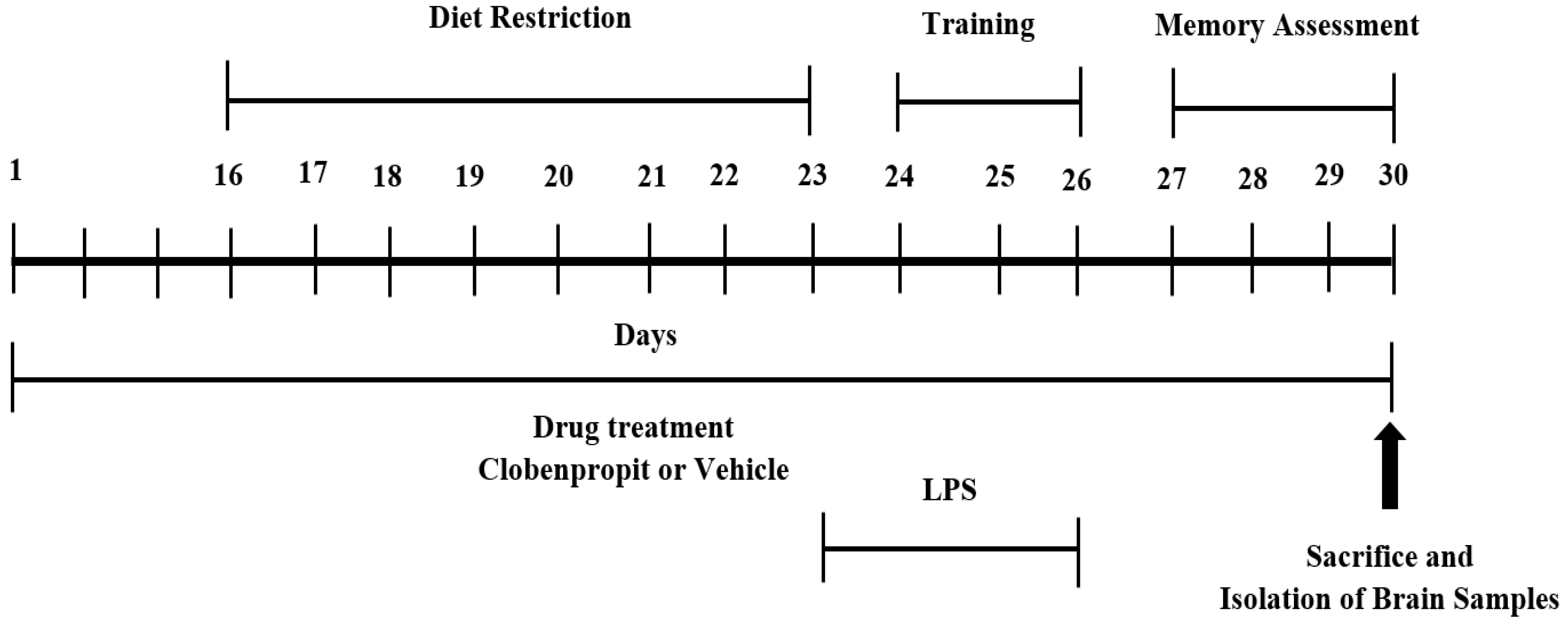
2.3. Experimental Design
2.4. Behavioral Assessment Using the Radial Arm Maze (RAM)
2.5. Collection of Brain Homogenate
2.6. Determination of Cytokines and Cyclooxygenase levels
2.7. Determination of Mitochondrial Respiratory Chain Complexes (MRCC)
2.8. Statistical Analysis
3. Results
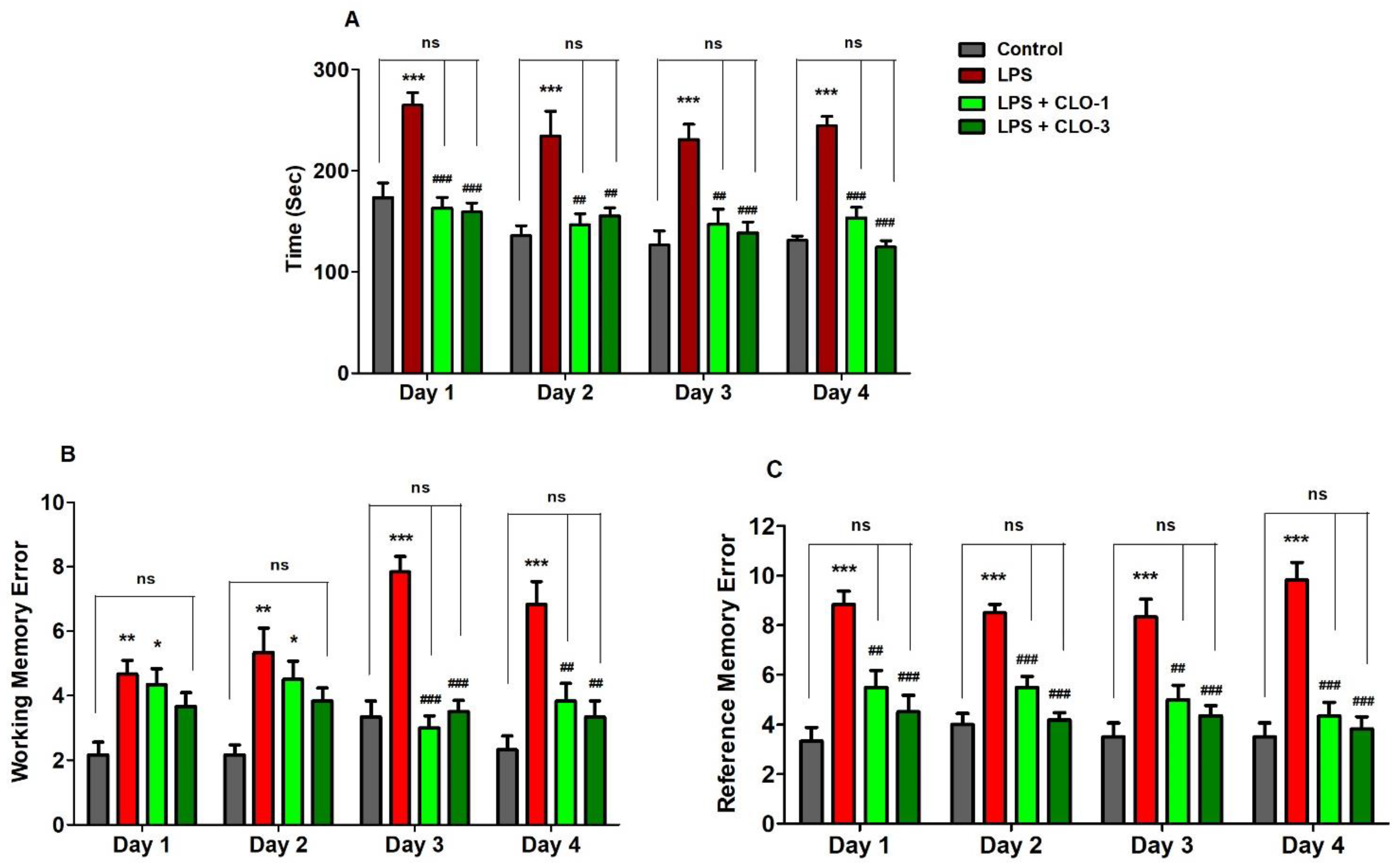
3.1. CLO Improved Memory Functions in LPS-Treated Mice Using the RAM
3.2. CLO Reduced Pro-Inflammatory Cytokine Levels in LPS-Treated Mice
3.3. CLO Improved Anti-Inflammatory Cytokine Levels in LPS-Treated Mice
3.4. CLO Reduced Cyclooxygenase-2 (COX-2) Activities in LPS-Treated Mice
3.5. CLO Improved Mitochondrial Respiratory Chain Complexes (MRCC) Activities in LPS-Treated Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tripathi, A.; Paliwal, P.; Krishnamurthy, S. Piracetam Attenuates LPS-Induced Neuroinflammation and Cognitive Impairment in Rats. Cell Mol. Neurobiol. 2017, 37, 1373–1386. [Google Scholar] [CrossRef]
- Kwon, H.S.; Koh, S.-H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Mani, V.; Jaafar, S.M.; Azahan, N.S.M.; Ramasamy, K.; Lim, S.M.; Ming, L.C.; Majeed, A.B.A. Ciproxifan improves cholinergic transmission, attenuates neuroinflammation and oxidative stress but does not reduce amyloid level in transgenic mice. Life Sci. 2017, 180, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, E.; Musich, P.; Lin, F. Mitochondrial dysfunction in neurodegenerative diseases and the potential countermeasure. CNS Neurosci. Ther. 2019, 25, 816–824. [Google Scholar] [CrossRef] [Green Version]
- Joshi, R.; Garabadu, D.; Teja, G.R.; Krishnamurthy, S. Silibinin ameliorates LPS-induced memory deficits in experimental animals. Neurobiol. Learn. Mem. 2014, 116, 117–131. [Google Scholar] [CrossRef] [PubMed]
- van Horssen, J.; van Schaik, P.; Witte, M. Inflammation and mitochondrial dysfunction: A vicious circle in neurodegenerative disorders? Neurosci. Lett. 2019, 710, 132931. [Google Scholar] [CrossRef]
- Marefati, N.; Beheshti, F.; Memarpour, S.; Bayat, R.; Shafei, M.N.; Sadeghnia, H.R.; Ghazavi, H.; Hosseini, M. The effects of acetyl-11-keto-β-boswellic acid on brain cytokines and memory impairment induced by lipopolysaccharide in rats. Cytokine 2020, 131, 155107. [Google Scholar] [CrossRef]
- Ko, C.-Y.; Chang, L.-H.; Lee, Y.-C.; Sterneck, E.; Cheng, C.-P.; Chen, S.-H.; Huang, A.-M.; Tseng, J.T.; Wang, J.-M. CCAAT/enhancer binding protein delta (CEBPD) elevating PTX3 expression inhibits macrophage-mediated phagocytosis of dying neuron cells. Neurobiol. Aging 2010, 33, 422.e11–422.e25. [Google Scholar] [CrossRef]
- Gemkow, M.J.; Davenport, A.J.; Harich, S.; Ellenbroek, B.; Cesura, A.; Hallett, D. The histamine H3 receptor as a therapeutic drug target for CNS disorders. Drug Discov. Today 2009, 14, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Jansen, F.P.; Mochizuki, T.; Yamamoto, Y.; Timmerman, H.; Yamatodani, A. In vivo modulation of rat hypothalamic histamine release by the histamine H3 receptor ligands, immepip and clobenpropit. Effects of intrahypothalamic and peripheral application. Eur. J. Pharmacol. 1998, 362, 149–155. [Google Scholar] [CrossRef]
- Munzar, P.; Tanda, G.; Justinova, Z.; Goldberg, S.R. Histamine H3 Receptor Antagonists Potentiate Methamphetamine Self-Administration and Methamphetamine-Induced Accumbal Dopamine Release. Neuropsychopharmacology 2004, 29, 705–717. [Google Scholar] [CrossRef]
- Blandina, P.; Giorgetti, M.; Bartolini, L.; Cecchi, M.; Timmerman, H.; Leurs, R.; Pepeu, G.; Giovannini, M. Inhibition of cortical acetylcholine release and cognitive performance by histamine H3 receptor activation in rats. Br. J. Pharmacol. 1996, 119, 1656–1664. [Google Scholar] [CrossRef] [Green Version]
- Aquino-Miranda, G.; Osorio-Espinoza, A.; Escamilla-Sánchez, J.; González-Pantoja, R.; Ortiz, J.; Arias-Montaño, J.-A. Histamine H3 receptors modulate depolarization-evoked [3H]-noradrenaline release from rat olfactory bulb slices. Neuropharmacology 2012, 62, 1127–1133. [Google Scholar] [CrossRef]
- Patnaik, R.; Sharma, A.; Skaper, S.D.; Muresanu, D.F.; Lafuente, J.V.; Castellani, R.J.; Nozari, A.; Sharma, H.S. Histamine H3 Inverse Agonist BF 2649 or Antagonist with Partial H4 Agonist Activity Clobenpropit Reduces Amyloid Beta Peptide-Induced Brain Pathology in Alzheimer’s Disease. Mol. Neurobiol. 2017, 55, 312–321. [Google Scholar] [CrossRef]
- Huang, Y.-W.; Hu, W.-W.; Chen, Z.; Zhang, L.-S.; Shen, H.-Q.; Timmerman, H.; Leurs, R.; Yanai, K. Effect of the histamine H3-antagonist clobenpropit on spatial memory deficits induced by MK-801 as evaluated by radial maze in Sprague–Dawley rats. Behav. Brain Res. 2004, 151, 287–293. [Google Scholar] [CrossRef]
- Delcourt, J.; Miller, N.Y.; Couzin, I.D.; Garnier, S. Methods for the effective study of collective behavior in a radial arm maze. Behav. Res. Methods 2018, 50, 1673–1685. [Google Scholar] [CrossRef]
- Rahim, N.S.; Lim, S.M.; Mani, V.; Hazalin, N.A.M.N.; Majeed, A.B.A.; Ramasamy, K. Virgin Coconut Oil-Induced Neuroprotection in Lipopolysaccharide-Challenged Rats is Mediated, in Part, Through Cholinergic, Anti-Oxidative and Anti-Inflammatory Pathways. J. Diet. Suppl. 2020, 18, 655–681. [Google Scholar] [CrossRef]
- Mohd Azahan, N.S.; Mani, V.; Ramasamy, K.; Lim, S.M.; Johari James, R.M.; Alsharidah, M.; Alhowail, A.; Abdul Majeed, A.B. Mahanimbine-induced neuroprotection via cholinergic system and at-tenuated amyloidogenesis as well as neuroinflammation in lipopolysaccharides-induced mice. Pharmacog. Mag. 2020, 16, 57–63. [Google Scholar]
- Bullón, P.; Román-Malo, L.; Marín-Aguilar, F.; Alvarez-Suarez, J.M.; Giampieri, F.; Battino, M.; Cordero, M.D. Lipophilic antioxidants prevent lipopolysaccharide-induced mitochondrial dysfunction through mitochondrial biogenesis improvement. Pharmacol. Res. 2014, 91, 1–8. [Google Scholar] [CrossRef]
- Abdel-Salam, O.M.E.; Abdel-Rahman, R.F.; Sleem, A.; Farrag, A.R. Modulation of lipopolysaccharide-induced oxidative stress by capsaicin. Inflammopharmacology 2011, 20, 207–217. [Google Scholar] [CrossRef]
- Wang, J.; Liu, B.; Xu, Y.; Luan, H.; Wang, C.; Yang, M.; Zhao, R.; Song, M.; Liu, J.; Sun, L.; et al. Thioperamide attenuates neuroinflammation and cognitive impairments in Alzheimer’s disease via inhibiting gliosis. Exp. Neurol. 2022, 347, 113870. [Google Scholar] [CrossRef]
- Mei, J.; Kohler, J.; Winter, Y.; Spies, C.; Endres, M.; Banneke, S.; Emmrich, J.V. Automated radial 8-arm maze: A voluntary and stress-free behavior test to assess spatial learning and memory in mice. Behav. Brain Res. 2019, 381, 112352. [Google Scholar] [CrossRef] [PubMed]
- Kongsui, R.; Sriraksa, N.; Thongrong, S. The Neuroprotective Effect of Zingiber cassumunar Roxb. Extract on LPS-Induced Neuronal Cell Loss and Astroglial Activation within the Hippocampus. BioMed Res. Int. 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Rubio-Perez, J.M.; Morillas-Ruiz, J.M. A Review: Inflammatory Process in Alzheimer’s Disease, Role of Cytokines. Sci. World J. 2012, 2012, 756357. [Google Scholar] [CrossRef]
- Perry, V. The influence of systemic inflammation on inflammation in the brain: Implications for chronic neurodegenerative disease. Brain Behav. Immun. 2004, 18, 407–413. [Google Scholar] [CrossRef] [PubMed]
- O’Banion, M.K. Cyclooxygenase-2: Molecular Biology, Pharmacology, and Neurobiology. Crit. Rev. Neurobiol. 1999, 13, 45–82. [Google Scholar] [CrossRef]
- Consilvio, C.; Vincent, A.M.; Feldman, E.L. Neuroinflammation, COX-2, and ALS—A dual role? Exp. Neurol. 2004, 187, 1–10. [Google Scholar] [CrossRef]
- Zhu, J.; Li, S.; Zhang, Y.; Ding, G.; Zhu, C.; Huang, S.; Zhang, A.; Jia, Z.; Li, M. COX-2 contributes to LPS-induced Stat3 activation and IL-6 production in microglial cells. Am. J. Transl. Res. 2018, 10, 966–974. [Google Scholar]
- Zhao, D.; Zhang, L.J.; Huang, T.Q.; Kim, J.; Gu, M.-Y.; Yang, H.O. Narciclasine inhibits LPS-induced neuroinflammation by modulating the Akt/IKK/NF-κB and JNK signaling pathways. Phytomedicine 2021, 85, 153540. [Google Scholar] [CrossRef]
- Brochu, M.-E.; Girard, S.; Lavoie, K.; Sébire, G. Developmental regulation of the neuroinflammatory responses to LPS and/or hypoxia-ischemia between preterm and term neonates: An experimental study. J. Neuroinflammation 2011, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Dudchenko, P.A. An overview of the tasks used to test working memory in rodents. Neurosci. Biobehav. Rev. 2004, 28, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Szczepanik, A.M.; Funes, S.; Petko, W.; Ringheim, G.E. IL-4, IL-10 and IL-13 modulate A beta(1--42)-induced cytokine and chemokine production in primary murine microglia and a human monocyte cell line. J. Neuroimmunol. 2001, 113, 49–62. [Google Scholar] [CrossRef]
- Vercellino, I.; Sazanov, L.A. The assembly, regulation and function of the mitochondrial respiratory chain. Nat. Rev. Mol. Cell Biol. 2021, 1–21. [Google Scholar] [CrossRef]
- Pollard, A.K.; Craig, E.L.; Chakrabarti, L. Mitochondrial Complex 1 Activity Measured by Spectrophotometry Is Reduced across All Brain Regions in Ageing and More Specifically in Neurodegeneration. PLoS ONE 2016, 11, e0157405. [Google Scholar] [CrossRef] [Green Version]
- Hoekstra, A.S.; Bayley, J.-P. The role of complex II in disease. Biochim. Biophys. Acta (BBA) Bioenerg. 2013, 1827, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Fontanesi, F.; Soto, I.C.; Horn, D.; Barrientos, A. Assembly of mitochondrial cytochromec-oxidase, a complicated and highly regulated cellular process. Am. J. Physiol. Physiol. 2006, 291, C1129–C1147. [Google Scholar] [CrossRef] [Green Version]
- Karan, K.R.; Trumpff, C.; McGill, M.A.; Thomas, J.E.; Sturm, G.; Lauriola, V.; Sloan, R.P.; Rohleder, N.; Kaufman, B.A.; Marsland, A.L.; et al. Mitochondrial respiratory capacity modulates LPS-induced inflammatory signatures in human blood. Brain Behav. Immun. Health 2020, 5, 100080. [Google Scholar] [CrossRef]
- Dumitru, C.; Kabat, A.M.; Maloy, K.J. Metabolic Adaptations of CD4+ T Cells in Inflammatory Disease. Front. Immunol. 2018, 9, 540. [Google Scholar] [CrossRef]
- Noble, F.; Rubira, E.; Boulanouar, M.; Palmier, B.; Plotkine, M.; Warnet, J.M.; Marchand-Leroux, C.; Massicot, F. Acute systemic inflammation induces central mitochondrial damage and mnesic deficit in adult Swiss mice. Neurosci. Lett. 2007, 424. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mani, V.; Arfeen, M.; Ali, H.M.; Abdel-Moneim, A.-M.H.; Aldubayan, M.; Alhowail, A. Neuroprotective Effect of Clobenpropit against Lipopolysaccharide-Induced Cognitive Deficits via Attenuating Neuroinflammation and Enhancing Mitochondrial Functions in Mice. Brain Sci. 2021, 11, 1617. https://doi.org/10.3390/brainsci11121617
Mani V, Arfeen M, Ali HM, Abdel-Moneim A-MH, Aldubayan M, Alhowail A. Neuroprotective Effect of Clobenpropit against Lipopolysaccharide-Induced Cognitive Deficits via Attenuating Neuroinflammation and Enhancing Mitochondrial Functions in Mice. Brain Sciences. 2021; 11(12):1617. https://doi.org/10.3390/brainsci11121617
Chicago/Turabian StyleMani, Vasudevan, Minhajul Arfeen, Hussein M. Ali, Abdel-Moneim Hafez Abdel-Moneim, Maha Aldubayan, and Ahmad Alhowail. 2021. "Neuroprotective Effect of Clobenpropit against Lipopolysaccharide-Induced Cognitive Deficits via Attenuating Neuroinflammation and Enhancing Mitochondrial Functions in Mice" Brain Sciences 11, no. 12: 1617. https://doi.org/10.3390/brainsci11121617