The Involvement of Hypothalamus–Pituitary–Adrenal (HPA) Axis in Suicide Risk

 , ,
, ,
Abstract
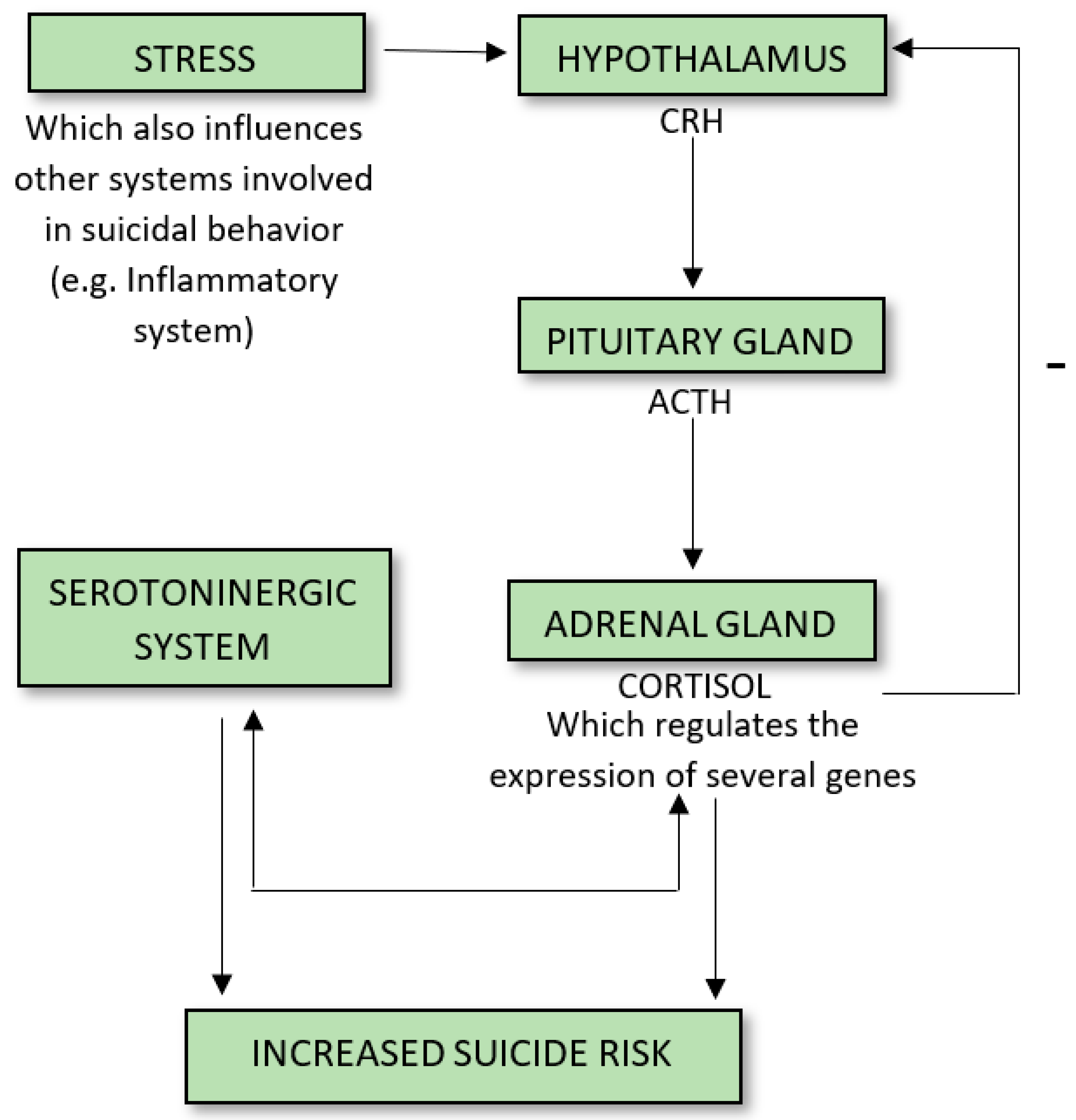
:1. Introduction
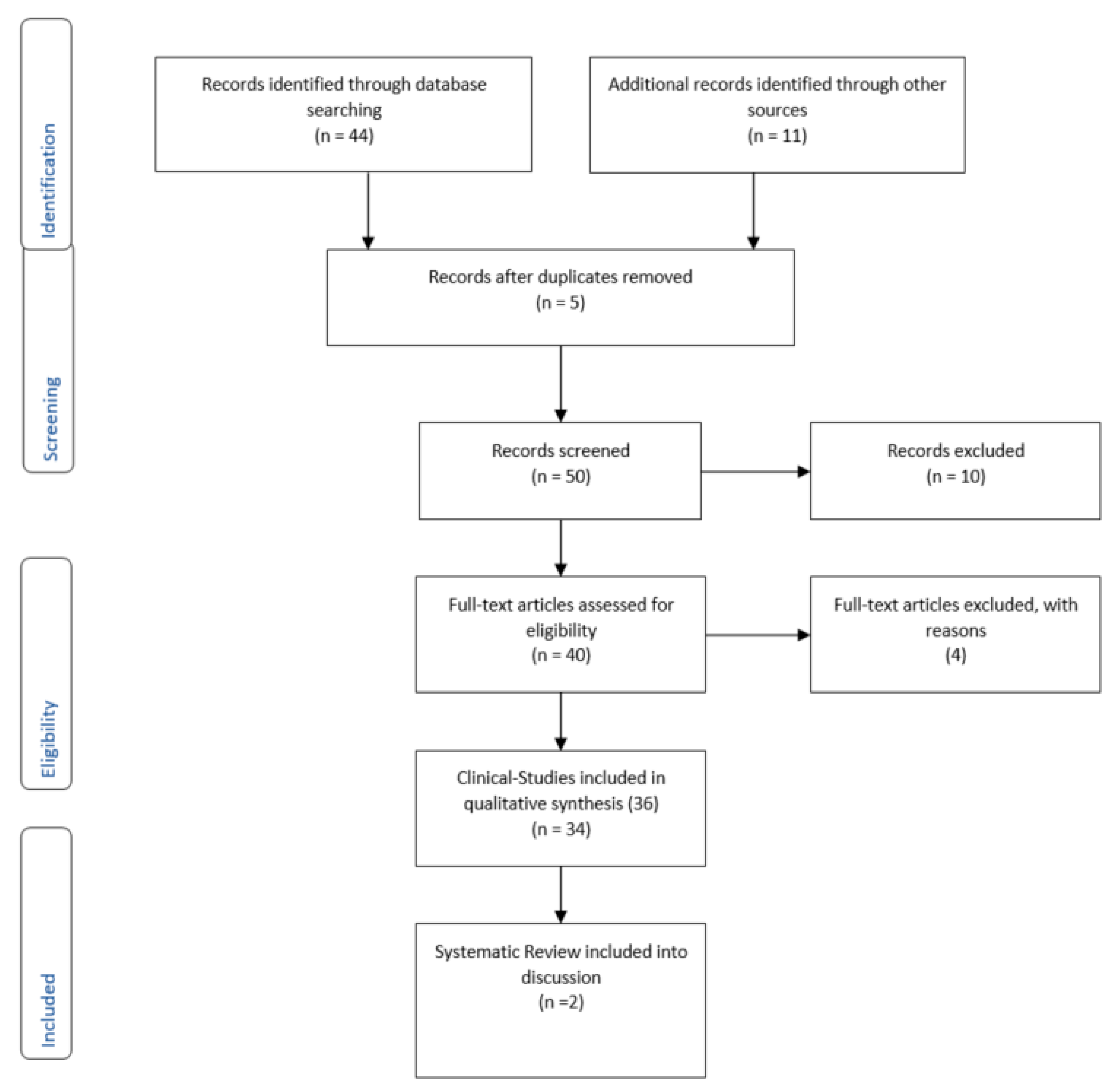
2. Methods
3. Results
3.1. CRH, Cortical Hypertrophy, CRF Binding Sites, and MHPG
3.2. FK506-Binding Protein 51 (FKBP5/FKBP51)
3.3. Tests on the Function of the HPA Axis Dysregulation
3.3.1. Dexamethasone Suppression Test (DST)
3.3.2. The Trier Social Stress Test and (TSSTa) and Maastricht Acute Stress Test (MAST)
3.4. Early Life Stress, HPA-Axis Dysregulation, and Suicide
3.5. Post-Mortem Studies
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Turecki, G.; Brent, D.A. Suicide and suicidal behaviour. Lancet 2016, 387, 1227–1239. [Google Scholar] [CrossRef]
- Pompili, M.; Gibiino, S.; Innamorati, M.; Serafini, G.; Del Casale, A.; De Risio, L.; Palermo, M.; Montebovi, F.; Campi, S.; De Luca, V.; et al. Prolactin and thyroid hormone levels are associated with suicide attempts in psychiatric patients. Psychiatry Res. 2012, 200, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Turecki, G.; Brent, D.A.; Gunnell, D.; O’Connor, R.C.; Oquendo, M.A.; Pirkis, J.; Stanley, B.H. Suicide and suicide risk. Nat. Rev. Dis. Primers 2019, 5, 74. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.C.; Kirtley, O.J. The Integrated Motivational-Volitional Model of Suicidal Behaviour. Philos. Trans. R. Soc. B 2018, 373, 20170268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selby, E.A.; Joiner, T.E., Jr.; Ribeiro, J. Comprehensive Theories of Suicidal Behaviors. In The Oxford Handbook of Suicide and Self-Injury; Matthew, K., Ed.; Oxford University Press: New York, NY, USA, 2014; pp. 286–307. [Google Scholar]
- Mann, J.J.; Waternaux, C.; Haas, G.L.; Malone, K.M. Toward a clinical model of suicidal behavior in psychiatric patients. Am. J. Psychiatry 1999, 156, 181–189. [Google Scholar]
- Hawton, K.; Van Heeringen, K. Suicide. Lancet 2009, 373, 1372–1381. [Google Scholar] [CrossRef]
- Labonte, B.; Turecki, G. The epigenetics of suicide: Explaining the biological effects of early life environmental adversity. Arch. Suicide Res. 2010, 14, 291–310. [Google Scholar] [CrossRef]
- Lutz, P.E.; Mechawar, N.; Turecki, G. Neuropathology of suicide: Recent findings and future directions. Mol. Psychiatry 2017, 22, 1395–1412. [Google Scholar] [CrossRef]
- Pompili, M.; Shrivastava, A.; Serafini, G.; Innamorati, M.; Milelli, M.; Erbuto, D.; Ricci, F.; Lamis, D.A.; Scocco, P.; Amore, M.; et al. Bereavement after the suicide of a significant other. Indian J. Psychiatry 2013, 55, 256–263. [Google Scholar] [CrossRef]
- McEwen, B.S. Physiology and neurobiology of stress and adaptation: Central role of the brain. Physiol. Rev. 2007, 87, 873–904. [Google Scholar] [CrossRef] [Green Version]
- Dedovic, K.; Duchesne, A.; Andrews, J.; Engert, V.; Pruessner, J.C. The brain and the stress axis: The neural correlates of cortisol regulation in response to stress. Neuroimage 2009, 47, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Ventriglio, A.; Gentile, A.; Baldessarini, R.J.; Bellomo, A. Early-life stress and psychiatric disorders: Epidemiology, neurobiology and innovative pharmacological targets. Curr. Pharm. Des. 2015, 21, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Sudol, K.; Mann, J.J. Biomarkers of Suicide Attempt Behavior: Towards a Biological Model of Risk. Curr. Psychiatry Rep. 2017, 19, 31. [Google Scholar] [CrossRef] [PubMed]
- Menke, A. Is the HPA Axis as Target for Depression Outdated, or Is There a New Hope? Front. Psychiatry 2019, 10, 101. [Google Scholar] [CrossRef]
- Merali, Z.; Du, L.; Hrdina, P.; Palkovits, M.; Faludi, G.; Poulter, M.O.; Anisman, H. Dysregulation in the suicide brain: mRNA expression of corticotropin-releasing hormone receptors and GABA (A) receptor subunits in frontal cortical brain region. J. Neurosci. 2004, 24, 1478–1485. [Google Scholar] [CrossRef] [Green Version]
- Austin, M.C.; Janosky, J.E.; Murphy, H.A. Increased corticotropin-releasing hormone immunoreactivity in monoamine-containing pontine nuclei of depressed suicide men. Mol. Psychiatry 2003, 8, 324–332. [Google Scholar] [CrossRef] [Green Version]
- Arató, M.; Bánki, C.M.; Bissette, G.; Nemeroff, C.B. Elevated CSF CRF in suicide victims. Biol. Psychiatry 1989, 25, 355–359. [Google Scholar]
- Dumser, T.; Barocka, A.; Schubert, E. Weight of adrenal glands may be increased in persons who commit suicide. Am. J. Forensic Med. Pathol. 1998, 19, 72–76. [Google Scholar] [CrossRef]
- Szigethy, E.; Conwell, Y.; Forbes, N.T.; Cox, C.; Caine, E.D. Adrenal weight and morphology in victims of completed suicide. Biol. Psychiatry 1994, 36, 374–380. [Google Scholar] [CrossRef]
- Stein, E.; McCrank, E.; Schaefer, B.; Goyer, R. Adrenal gland weight and suicide. Can. J. Psychiatry 1993, 38, 563–566. [Google Scholar] [CrossRef]
- Nemeroff, C.B.; Owens, M.J.; Bissette, G.; Andorn, A.C.; Stanley, M. Reduced corticotropin releasing factor binding sites in the frontal cortex of suicide victims. Arch. Gen. Psychiatry 1988, 45, 577–579. [Google Scholar] [CrossRef] [PubMed]
- Merali, Z.; Kent, P.; Du, L.; Hrdina, P.; Palkovits, M.; Faludi, G.; Poulter, M.O.; Bédard, T.; Anisman, H. Corticotropin-releasing hormone, arginine vasopressin, gastrin-releasing peptide, and neuromedin B alterations in stress-relevant brain regions of suicides and control subjects. Biol. Psychiatry 2006, 59, 594–602. [Google Scholar] [CrossRef] [PubMed]
- López, J.F.; Palkovits, M.; Arató, M.; Mansour, A.; Akil., H.; Watson, S.J. Localization and quantification of pro-opiomelanocortin mRNA and glucocorticoid receptor mRNA in pituitaries of suicide victims. Neuroendocrinology 1992, 56, 491–501. [Google Scholar]
- Jokinen, J.; Ouda, J.; Nordström, P. Noradrenergic function and HPA axis dysregulation in suicidal behaviour. Psychoneuroendocrinology 2010, 35, 1536–1542. [Google Scholar] [CrossRef]
- Jokinen, J.; Boström, A.E.; Dadfar, A.; Ciuculete, D.M.; Chatzittofis, A.; Asberg, M.; Schioth, H.B. Epigenetic Changes in the CRH Gene are Related to Severity of Suicide Attempt and a General Psychiatric Risk Score in Adolescents. EBioMedicine 2018, 27, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Cioffi, D.L.; Hubler, T.R.; Scammell, J.G. Organization and function of the FKBP52 and FKBP51 genes. Curr. Opin. Pharmacol. 2011, 11, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Fudalej, S.; Kopera, M.; Wołyńczyk-Gmaj, D.; Fudalej, M.; Krajewski, P.; Wasilewska, K.; Szymański, K.; Chojnicka, I. Association between FKBP5 Functional Polymorphisms and Completed Suicide. Neuropsychobiology 2015, 72, 126–131. [Google Scholar] [CrossRef]
- Supriyanto, I.; Sasada, T.; Fukutake, M.; Asano, M.; Ueno, Y.; Nagasaki, Y.; Shirakawa, O.; Hishimoto, A. Association of FKBP5 gene haplotypes with completed suicide in the Japanese population. Prog. Neuropsychopharm. Biol. Psychiatry 2011, 35, 252–256. [Google Scholar] [CrossRef]
- Giletta, M.; Calhoun, C.D.; Hastings, P.D.; Rudolph, K.D.; Nock, M.K.; Prinstein, M.J. Multi-Level Risk Factors for Suicidal Ideation Among at-Risk Adolescent Females: The Role of Hypothalamic-Pituitary-Adrenal Axis Responses to Stress. J. Abnorm. Child Psychol. 2015, 43, 807–820. [Google Scholar] [CrossRef]
- Keilp, J.G.; Stanley, B.H.; Burke, A.K.; Melhem, N.M.; Oquendo, M.A.; Brent, D.A.; Mann, J.J. Further evidence of low baseline cortisol levels in suicide attempters. J. Affect. Disord. 2016, 190, 187–192. [Google Scholar] [CrossRef]
- Yerevanian, B.I.; Feusner, J.D.; Koek, R.J.; Mintz, J. The dexamethasone suppression test as a predictor of suicidal behavior in unipolar depression. J. Affect. Disord. 2004, 83, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Jokinen, J.; Carlborg, A.; Martensson, B.; Forslund, K.; Nordström, A.L.; Nordström, P. DST non-suppression predicts suicide after attempted suicide. Psychiatry Res. 2007, 150, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Jokinen, J.; Nordström, P. HPA axis hyperactivity as suicide predictor in elderly mood disorder inpatients. Psychoneuroendocrinology 2008, 33, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Jokinen, J.; Nordström, P. HPA axis hyperactivity and attempted suicide in young adult mood disorder inpatients. J. Affect. Disord. 2009, 116, 117–120. [Google Scholar] [CrossRef]
- Coryell, W.; Schlesser, M. The dexamethasone suppression test and suicide prediction. Am. J. Psychiatry 2001, 158, 748–753. [Google Scholar] [CrossRef]
- Beauchaine, T.P.; Crowell, S.E.; Hsiao, R.C. Post-dexamethasone cortisol, self-inflicted injury, and suicidal ideation among depressed adolescent girls. J. Abnorm. Child Psychol. 2015, 43, 619–632. [Google Scholar] [CrossRef] [Green Version]
- Ambrus, L.; Westling, S. Leptin, Anxiety Symptoms, and Hypothalamic-Pituitary-Adrenal Axis Activity among Drug-Free, Female Suicide Attempters. Neuropsychobiology 2019, 78, 145–152. [Google Scholar] [CrossRef]
- Melhem, N.M.; Keilp, J.G.; Porta, G.; Oquendo, M.A.; Burke, A.; Stanley, B.; Cooper, T.B.; Mann, J.J.; Brent, D.A. Blunted HPA Axis Activity in Suicide Attempters Compared to those at High Risk for Suicidal Behavior. Neuropsychopharmacology 2016, 41, 1447–1456. [Google Scholar] [CrossRef] [Green Version]
- Melhem, N.M.; Munroe, S.; Marsland, A.; Gray, K.; Brent, D.; Porta, G.; Douaihy, A.; Laudenslager, M.L.; DePietro, F.; Diler, R.; et al. Blunted HPA axis activity prior to suicide attempt and increased inflammation in attempters. Psychoneuroendocrinology 2017, 77, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Eisenlohr-Moul, T.A.; Miller, A.B.; Giletta, M.; Hastings, P.D.; Rudolph, K.D.; Nock, M.K.; Prinstein, M.J. HPA axis response and psychosocial stress as interactive predictors of suicidal ideation and behavior in adolescent females: A multilevel diathesis-stress framework. Neuropsychopharmacology 2018, 43, 2564–2571. [Google Scholar] [CrossRef] [Green Version]
- Stanley, B.; Michel, C.A.; Galfalvy, H.C.; Keilp, J.G.; Rizk, M.M.; Richardson-Vejlgaard, R.; Oquendo, M.A.; Mann, J.J. Suicidal subtypes, stress responsivity and impulsive aggression. Psychiatry Res. 2019, 280, 112486. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.M.; Galfalvy, H.; Singh, T.; Keilp, J.G.; Sublette, M.E.; Oquendo, M.A.; Mann, J.J.; Stanley, B. Toward subtyping of suicidality: Brief suicidal ideation is associated with greater stress response. J. Affect. Disord. 2018, 230, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Shalev, A.; Porta, G.; Biernesser, C.; Zelazny, J.; Walker-Payne, M.; Melhem, N.; Brent, D. Cortisol response to stress as a predictor for suicidal ideation in youth. J. Affect. Disord. 2019, 257, 10–16. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, D.B.; Green, J.A.; Ferguson, E.; O’Carroll, R.E.; O’Connor, R.C. Cortisol reactivity and suicidal behavior: Investigating the role of hypothalamic-pituitary-adrenal axis responses to stress in suicide attempters and ideators. Psychoneuroendocrinology 2017, 75, 183–191. [Google Scholar] [CrossRef] [Green Version]
- McCrory, E.; De Brito, S.A.; Viding, E. The link between child abuse and psychopathology: A review of neurobiological and genetic research. J. R. Soc. Med. 2012, 105, 151–156. [Google Scholar] [CrossRef]
- Jaffee, S.R. Child Maltreatment and Risk for Psychopathology in Childhood and Adulthood. Annu. Rev. Clin. Psychol. 2017, 13, 525–551. [Google Scholar] [CrossRef]
- Green, J.G.; Mclaughlin, K.A.; Berglund, P.A.; Gruber, M.J.; Sampson, N.A.; Zaslavsky, A.M.; Kessler, R.C. Childhood Adversities and Adult Psychiatric Disorders in the National Comorbidity Survey Replication I. Arch. Gen. Psychiatry 2010, 67, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Heim, C.; Shugart, M.; Shugart, M.; Craighead, W.E.; Nemeroff, C.B. Neurobiological and psychiatric consequences of child abuse and neglect. Dev. Psychobiol. 2010, 52, 671–690. [Google Scholar] [CrossRef]
- Heim, C.; Newport, D.J.; Mletzko, T.M.; Bonsall, R.; Miller, A.H.; Nemeroff, C.B. The link between childhood trauma and depression: Insights from HPA axis studies in humans. Psychoneuroendocrinology 2008, 33, 693–710. [Google Scholar] [CrossRef]
- McGowan, P.O.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; Labonté, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, D.B.; Branley-Bell, D.; Green, J.A.; Ferguson, E.; O’Carroll, R.E.; O’Connor, R.C. Effects of childhood trauma, daily stress, and emotions on daily cortisol levels in individuals vulnerable to suicide. J. Abnorm. Psychol. 2020, 129, 92–107. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Hodgkinson, C.A.; Deluca, V.; Goldman, D.; Enoch, M.A. Two HPA axis genes, CRHBP and FKBP5, interact with childhood trauma to increase the risk for suicidal behavior. J. Psychiatr. Res. 2012, 46, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, D.B.; Green, J.A.; Ferguson, E.; O’Carroll, R.E.; O’Connor, R.C. Effects of childhood trauma on cortisol levels in suicide attempters and ideators. Psychoneuroendocrinology 2018, 88, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanabrais-Jiménez, M.A.; Sotelo-Ramirez, C.E.; Ordoñez-Martinez, B.; Jiménez-Pavón, J.; Ahumada-Curiel, G.; Piana-Diaz, S.; Flores-Flores, G.; Flores-Ramos, M.; Jiménez-Anguiano, A.; Camarena, B. Effect of CRHR1 and CRHR2 gene polymorphisms and childhood trauma in suicide attempt. J. Neural. Transm. 2019, 126, 637–644. [Google Scholar] [CrossRef]
- Pandey, G.N.; Rizavi, H.S.; Ren, X.; Dwivedi, Y.; Palkovits, M. Region-specific alterations in glucocorticoid receptor expression in the postmortem brain of teenage suicide victims. Psychoneuroendocrinology 2013, 38, 2628–2639. [Google Scholar] [CrossRef] [Green Version]
- Labonte, B.; Yerko, V.; Gross, J.; Mechawar, N.; Meaney, M.J.; Szyf, M.; Turecki, G. Differential glucocorticoid receptor exon 1(B), 1(C), and 1(H) expression and methylation in suicide completers with a history of childhood abuse. Biol. Psychiatry 2012, 72, 41–48. [Google Scholar] [CrossRef]
- Janiri, D.; De Rossi, P.; Kotzalidis, G.D.; Girardi, P.; Koukopoulos, A.E.; Reginaldi, D.; Dotto, F.; Manfredi, G.; Jollant, F.; Gorwood, P.; et al. Psychopathological characteristics and adverse childhood events are differentially associated with suicidal ideation and suicidal acts in mood disorders. Eur. Psychiatry 2018, 53, 31–36. [Google Scholar] [CrossRef]
- Pompili, M.; Serafini, G.; Innamorati, M.; Möller-Leimkühler, A.M.; Giupponi, G.; Girardi, P.; Tatarelli, R.; Lester, D. The hypothalamic-pituitary-adrenal axis and serotonin abnormalities: A selective overview for the implications of suicide prevention. Eur. Arch. Psychiatry Clin. Neurosci. 2010, 260, 583–600. [Google Scholar] [CrossRef]
- Heim, C.; Nemeroff, C.B. The role of childhood trauma in the neurobiology of mood and anxiety disorders: Preclinical and clinical studies. Biol. Psychiatry 2001, 49, 1023–1039. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Author, Year | Sample | Method | Results |
|---|---|---|---|
| Giletta et al. 2015 [30] | 138 adolescent females at risk for SB | PST | The hyperresponsive group was more likely to report a lifetime history of SI |
| Jokinen et al. 2007 [33] | 382 psychiatric inpatients with mood disorder | DST | Non-suppressor status was significantly associated with suicide |
| Jokinen et al. 2008 [34] | 99 depressed elderly inpatients | DST | DST non-suppression distinguished between suicides and survivors in this sample |
| Jokinen et al. 2009 [35] | 36 patients with mood disorder, with (n = 18) and without suicide attempt | DST | DST non-suppressor rate was 39% in those with a suicide attempt history and 11% in those with a non-attempt history |
| Coryell and Schlesser 2001 [36] | 78 patients with mood disorder | DST | Estimated risk for suicide in the group with abnormal DST results was 26.8%, compared to only 2.9% for the other group |
| Beauchaine et al. 2015 [37] | 57 adolescent females (13–17 years) | DST | Lower post-DST cortisol was associated with suicidal ideation and self-injury |
| Author, Year | Sample | Method | Results |
|---|---|---|---|
| Melhem et al. 2016 [40] | 208 offspring of parents with mood disorder | TSST | Offspring with a suicide attempt history showed lower total cortisol output |
| Eisenlohr-Moul et al. 2018 [41] | 220 adolescent females | TSST | Stress triggers suicidal behavior among those with blunted cortisol reactivity |
| Stanley et al. 2019 [42] | 72 patients with mood disorder (SA n = 35, non-SA n = 37) | TSST | Stress response differed based on impulsivity/aggression levels in those who had attempted suicide (with more pronounced cortisol response) |
| Rizk et al. 2018 [43] | 35 patients with MDD, 23 healthy controls | TSST | Patients with brief suicidal ideation presented greater cortisol response |
| Shalev et al. 2019 [44] | 114 youth bereaved by sudden parental death, 109 non-bereaved controls | TSST | Cortisol reactivity to stress was higher in those with high SI trajectory |
| O’Connor et al. 2017 [45] | 145 patients (47 attempter, 53 with SI, 45 control group) | MAST | HPA axis activity is markedly lower in suicide attempters compared to controls, but not ideators |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berardelli, I.; Serafini, G.; Cortese, N.; Fiaschè, F.; O’Connor, R.C.; Pompili, M. The Involvement of Hypothalamus–Pituitary–Adrenal (HPA) Axis in Suicide Risk. Brain Sci. 2020, 10, 653. https://doi.org/10.3390/brainsci10090653
Berardelli I, Serafini G, Cortese N, Fiaschè F, O’Connor RC, Pompili M. The Involvement of Hypothalamus–Pituitary–Adrenal (HPA) Axis in Suicide Risk. Brain Sciences. 2020; 10(9):653. https://doi.org/10.3390/brainsci10090653
Chicago/Turabian StyleBerardelli, Isabella, Gianluca Serafini, Natalia Cortese, Federica Fiaschè, Rory C O’Connor, and Maurizio Pompili. 2020. "The Involvement of Hypothalamus–Pituitary–Adrenal (HPA) Axis in Suicide Risk" Brain Sciences 10, no. 9: 653. https://doi.org/10.3390/brainsci10090653