Dream Recall upon Awakening from Non-Rapid Eye Movement Sleep in Older Adults: Electrophysiological Pattern and Qualitative Features

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Study Design
2.2.1. Polysomnography
2.2.2. Dream Collection
2.3. Data Analysis
2.3.1. Sleep Architecture
2.3.2. Quantitative EEG Analysis
2.3.3. Dream Reports
2.3.4. Statistical Analysis
3. Results
3.1. Sleep Architecture
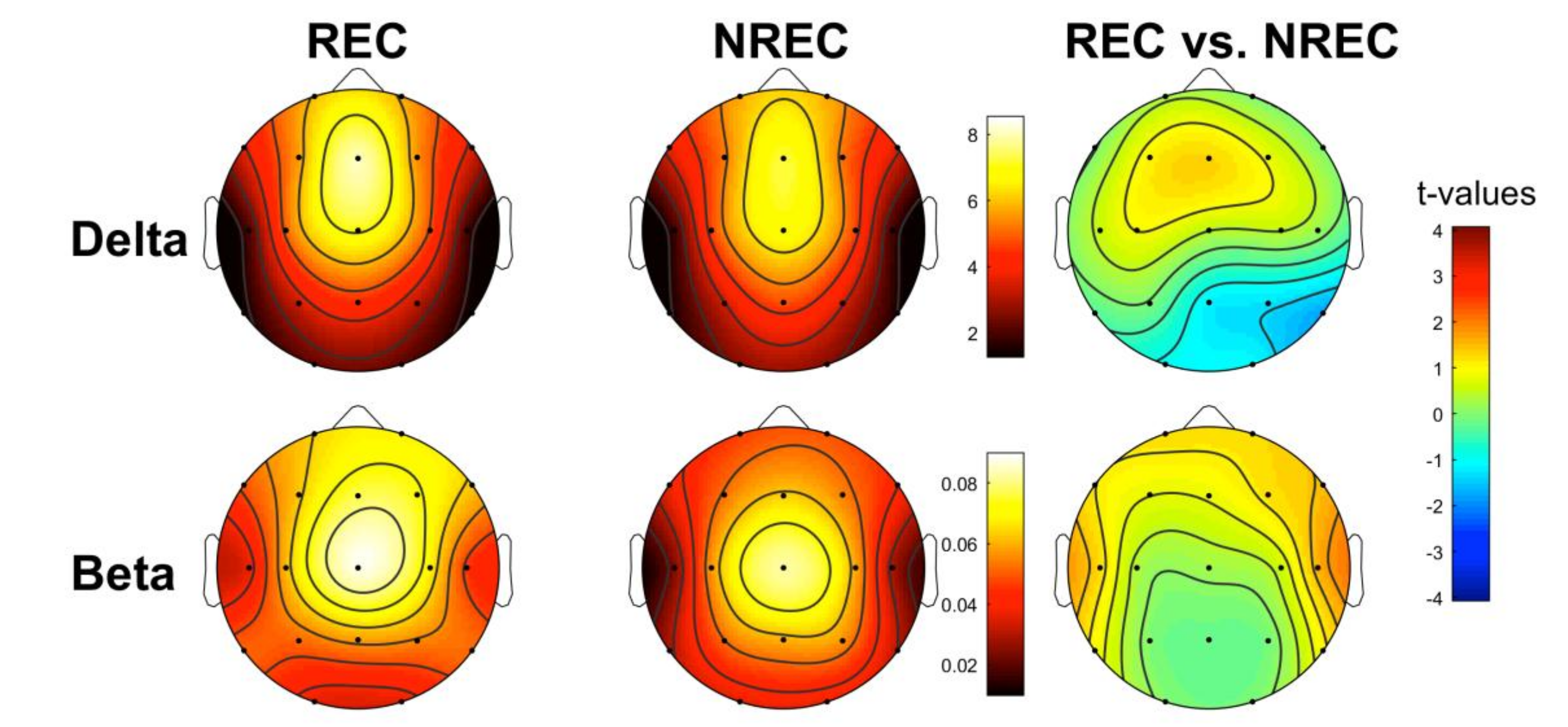
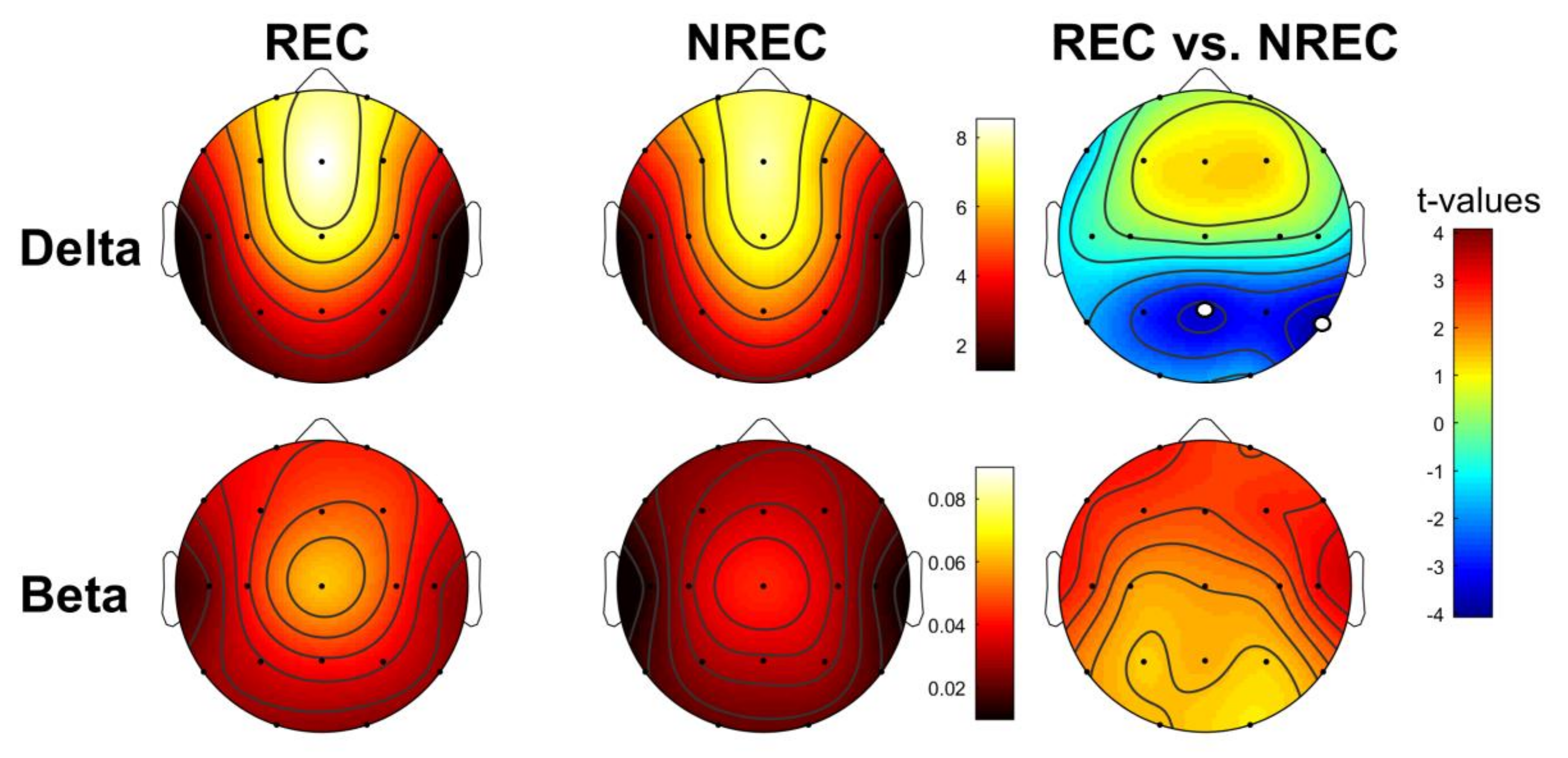
3.2. EEG Pattern of Dream Recall: Last 5 min of NREM Sleep
3.3. EEG Pattern of Dream Recall: Total NREM Sleep
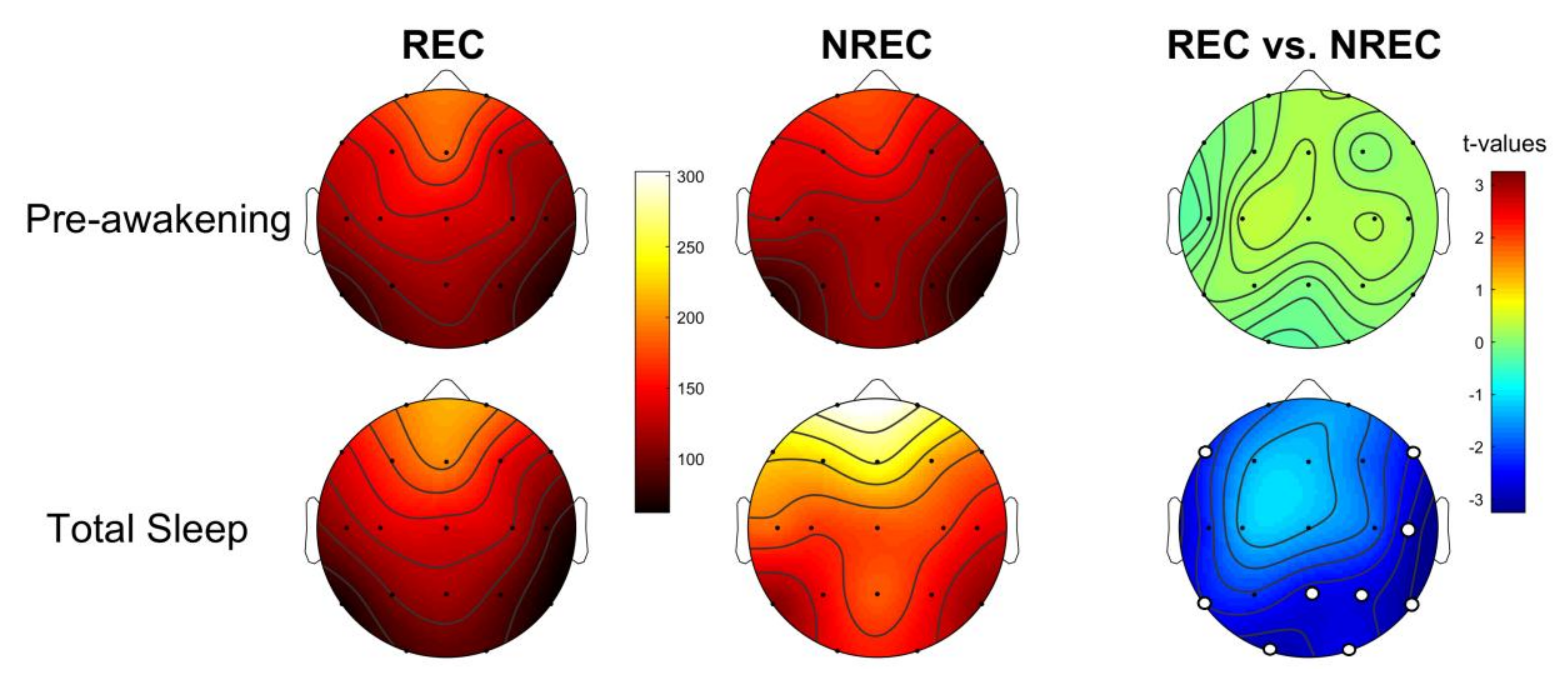
3.4. Activation Index as Predictor of Dream Recall
3.5. Sleep Pattern and Dream Features
4. Discussion
5. Limitations
- (a)
- the absence of the adaptation night that could have affected the continuity of sleep during the night and does not allow us to rule out the so-called first night effect;
- (b)
- the lack of intraindividual repeated measures and a relatively small sample size, not allowing us to disentangle the state-/trait-like issue [2].
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fagioli, I. Mental activity during sleep. Sleep Med. Rev. 2002, 6, 307–320. [Google Scholar] [CrossRef]
- Scarpelli, S.; D’Atri, A.; Gorgoni, M.; Ferrara, M.; De Gennaro, L.; D’Atri, A. EEG oscillations during sleep and dream recall: State or trait-like individual differences? Front. Psychol. 2015, 6. [Google Scholar] [CrossRef]
- De Gennaro, L.; Marzano, C.; Cipolli, C.; Ferrara, M. How we remember the stuff that dreams are made of: Neurobiological approaches to the brain mechanisms of dream recall. Behav. Brain Res. 2012, 226, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Foulkes, W.D. Dream reports from different stages of sleep. J. Abnorm. Soc. Psychol. 1962, 65, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Cipolli, C.; Ferrara, M.; De Gennaro, L.; Plazzi, G. Beyond the neuropsychology of dreaming: Insights into the neural basis of dreaming with new techniques of sleep recording and analysis. Sleep Med. Rev. 2017, 35, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Ogilvie, R.D.; Murphy, T.I.; Ferrelli, A.V. EEG activities during elicited sleep onset REM and NREM periods reflect different mechanisms of dream generation. Chin. Neurophysiol. 2003, 114, 210–220. [Google Scholar] [CrossRef]
- Esposito, M.J.; Nielsen, T.; Paquette, T. Reduced Alpha power associated with the recall of mentation from Stage 2 and Stage REM sleep. Psychophysiology 2004, 41, 288–297. [Google Scholar] [CrossRef]
- Marzano, C.; Ferrara, M.; Mauro, F.; Moroni, F.; Gorgoni, M.; Tempesta, D.; Cipolli, C.; De Gennaro, L. Recalling and Forgetting Dreams: Theta and Alpha Oscillations during Sleep Predict Subsequent Dream Recall. J. Neurosci. 2011, 31, 6674–6683. [Google Scholar] [CrossRef] [Green Version]
- Scarpelli, S.; D’Atri, A.; Mangiaruga, A.; Marzano, C.; Gorgoni, M.; Schiappa, C.; Ferrara, M.; De Gennaro, L. Predicting Dream Recall: EEG Activation During NREM Sleep or Shared Mechanisms with Wakefulness? Brain Topogr. 2017, 30, 629–638. [Google Scholar] [CrossRef]
- Siclari, F.; Baird, B.; Perogamvros, L.; Bernardi, G.; Larocque, J.J.; Riedner, B.; Boly, M.; Postle, B.R.; Tononi, G. The neural correlates of dreaming. Nat. Neurosci. 2017, 20, 872–878. [Google Scholar] [CrossRef] [Green Version]
- Siclari, F.; Bernardi, G.; Cataldi, J.; Tononi, G. Dreaming in NREM Sleep: A High-Density EEG Study of Slow Waves and Spindles. J. Neurosci. 2018, 38, 9175–9185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wamsley, E.J. EEG predictors of dreaming outside of REM sleep. Psychophysiology 2019, 56, e13368. [Google Scholar] [CrossRef] [PubMed]
- D’Atri, A.; Scarpelli, S.; Schiappa, C.; Pizza, F.; Vandi, S.; Ferrara, M.; Cipolli, C.; Plazzi, G.; De Gennaro, L. Cortical activation during sleep predicts dream experience in narcolepsy. Ann. Clin. Transl. Neurol. 2019, 6, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Antrobus, J. Cortical hemisphere asymmetry and sleep mentation. Psychol. Rev. 1987, 94, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Schredl, M. Dreams in patients with sleep disorders. Sleep Med. Rev. 2009, 13, 215–221. [Google Scholar] [CrossRef]
- Chellappa, S.L.; Frey, S.; Knoblauch, V.; Cajochen, C. Cortical activation patterns herald successful dream recall after NREM and REM sleep. Boil. Psychol. 2011, 87, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Mangiaruga, A.; Scarpelli, S.; Bartolacci, C.; De Gennaro, L. Spotlight on dream recall: The ages of dreams. Nat. Sci. Sleep 2018, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mander, B.A.; Winer, J.R.; Walker, M.P. Sleep and Human Aging. Neuron 2017, 94, 19–36. [Google Scholar] [CrossRef] [Green Version]
- Scarpelli, S.; Bartolacci, C.; D’Atri, A.; Gorgoni, M.; De Gennaro, L. Mental Sleep Activity and Disturbing Dreams in the Lifespan. Int. J. Environ. Res. Public Health 2019, 16, 3658. [Google Scholar] [CrossRef] [Green Version]
- Ancoli-Israel, S.; Alessi, C. Sleep and Aging. Am. J. Geriatr. Psychiatry 2005, 13, 341–343. [Google Scholar] [CrossRef]
- Giambra, L.M.; Jung, R.E.; Grodsky, A. Age changes in dream recall in adulthood. Dreaming 1996, 6, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, T. Variations in Dream Recall Frequency and Dream Theme Diversity by Age and Sex. Front. Neurol. 2012, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, D.B.; MacNeilage, P.F. A test of the salience hypothesis of dream recall. J. Consult. Clin. Psychol. 1974, 42, 699–703. [Google Scholar] [CrossRef] [PubMed]
- Cappeliez, P. An explanation of the reminiscence bump in the dreams of older adults in terms of life goals and identity. Self Identity 2008, 7, 25–33. [Google Scholar] [CrossRef]
- Funkhouser, A.; Hirsbrunner, H.-P.; Cornu, C.; Bahro, M. Dreams and dreaming among the elderly: An overview. Aging Ment. Health 1999, 3, 10–20. [Google Scholar] [CrossRef]
- Waterman, D. Aging and Memory for Dreams. Percept. Mot. Ski. 1991, 73, 355–365. [Google Scholar] [CrossRef]
- Kahn, E.; Fisher, C.; Lieberman, L. Dream Recall In The Normal Aged. J. Am. Geriatr. Soc. 1969, 17, 1121–1126. [Google Scholar] [CrossRef]
- Fein, G.; Feinberg, I.; Insel, T.R.; Antrobus, J.S.; Price, L.J.; Floyd, T.C.; Nelson, M.A. Sleep Mentation in the Elderly. Psychophysiology 1985, 22, 218–225. [Google Scholar] [CrossRef]
- De Padova, V.; Aiello, P.; Cocozza, G.; Ficca, G. Dream Recall in Elderly Subjects after Spontaneous awakening in Laboratory. In Proceedings of the World Association of Sleep Medicine, 1st Congress, Berlin, Germany, 7 July 2005; pp. 35–40. [Google Scholar]
- Cavallero, C.; Natale, V.; Occhionero, M.; Zito, A.; Cicogna, P. Slow Wave Sleep Dreaming. Sleep 1992, 15, 562–566. [Google Scholar] [CrossRef] [Green Version]
- Chellappa, S.L.; Münch, M.; Knoblauch, V.; Cajochen, C. Age effects on spectral electroencephalogram activity prior to dream recall. J. Sleep Res. 2011, 21, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Scarpelli, S.; D’Atri, A.; Bartolacci, C.; Mangiaruga, A.; Gorgoni, M.; De Gennaro, L. Oscillatory EEG Activity During REM Sleep in Elderly People Predicts Subsequent Dream Recall After Awakenings. Front. Neurol. 2019, 10, 985. [Google Scholar] [CrossRef] [PubMed]
- De Gennaro, L.; Cipolli, C.; Cherubini, A.; Assogna, F.; Cacciari, C.; Marzano, C.; Curcio, G.; Ferrara, M.; Caltagirone, C.; Spalletta, G. Amygdala and hippocampus volumetry and diffusivity in relation to dreaming. Hum. Brain Mapp. 2010, 32, 1458–1470. [Google Scholar] [CrossRef]
- De Gennaro, L.; Lanteri, O.; Piras, F.; Scarpelli, S.; Assogna, F.; Ferrara, M.; Caltagirone, C.; Spalletta, G. Dopaminergic system and dream recall: An MRI study in Parkinson’s disease patients. Hum. Brain Mapp. 2015, 37, 1136–1147. [Google Scholar] [CrossRef] [PubMed]
- Polini, F.; Principe, R.; Scarpelli, S.; Clementi, F.; De Gennaro, L. Use of varenicline in smokeless tobacco cessation influences sleep quality and dream recall frequency but not dream affect. Sleep Med. 2017, 30, 1–6. [Google Scholar] [CrossRef]
- Rechtschaffen, A.; Kales, A. A Manual of Standardized Terminology, Techniques and Scoring System for Sleep Stages of Human Subjects; UCLA Brain Information Service: Los Angeles, CA, USA, 1978. [Google Scholar]
- Perneger, T.V. What’s wrong with Bonferroni adjustments. BMJ 1998, 316, 1236–1238. [Google Scholar] [CrossRef] [PubMed]
- Sankoh, A.J.; Huque, M.F.; Dubey, S.D. Some comments on frequently used multiple endpoint adjustment methods in clinical trials. Stat. Med. 1997, 16, 2529–2542. [Google Scholar] [CrossRef]
- Koch, C.; Massimini, M.; Boly, M.; Tononi, G. Neural correlates of consciousness: Progress and problems. Nat. Rev. Neurosci. 2016, 17, 307–321. [Google Scholar] [CrossRef]
- Whitlock, J.R. Posterior parietal cortex. Curr. Boil. 2017, 27, R691–R695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, J.J.; Marois, R. Posterior parietal cortex activity predicts individual differences in visual short-term memory capacity. Cogn. Affect. Behav. Neurosci. 2005, 5, 144–155. [Google Scholar] [CrossRef] [Green Version]
- Murri, L.; Massetani, R.; Siciliano, G.; Giovanditti, L.; Arena, R. Dream Recall After Sleep Interruption in Brain-Injured Patients. Sleep 1985, 8, 356–362. [Google Scholar] [CrossRef] [Green Version]
- Solms, M. The Neuropsychology of Dreams: A Clinico-Anatomical Study; Erlbaum: Hillsdale, MI, USA, 1997. [Google Scholar]
- Bischof, M.; Bassetti, C.L. Total dream loss: A distinct neuropsychological dysfunction after bilateral PCA stroke. Ann. Neurol. 2004, 56, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Eichenlaub, J.-B.; Nicolas, A.; Daltrozzo, J.; Redouté, J.; Costes, N.; Ruby, P. Resting Brain Activity Varies with Dream Recall Frequency Between Subjects. Neuropsychopharmacology 2014, 39, 1594–1602. [Google Scholar] [CrossRef] [PubMed]
- Koulack, D.; Goodenough, D.R. Dream recall and dream recall failure: An arousal-retrieval model. Psychol. Bull. 1976, 83, 975. [Google Scholar] [CrossRef]
- Eichenlaub, J.-B.; Bertrand, O.; Morlet, D.; Ruby, P. Brain Reactivity Differentiates Subjects with High and Low Dream Recall Frequencies during Both Sleep and Wakefulness. Cereb. Cortex 2013, 24, 1206–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gennaro, L.; Marzano, C.; Moroni, F.; Curcio, G.; Ferrara, M.; Cipolli, C. Recovery sleep after sleep deprivation almost completely abolishes dream recall. Behav. Brain Res. 2010, 206, 293–298. [Google Scholar] [CrossRef]
- Carrasco, E.; Santamaria, J.; Iranzo, A.; Pintor, L.; De Pablo, J.; Solanas, A.; Kumru, H.; Boget, T.; Rodriguez, J.E.M. Changes in dreaming induced by CPAP in severe obstructive sleep apnea syndrome patients. J. Sleep Res. 2006, 15, 430–436. [Google Scholar] [CrossRef]
- Chabani, E.; Vionnet, M.C.; Beauté, R.; Leu-Semenescu, S.; Dodet, P.; Arnulf, I. Blackout of my nights: Contentless, timeless and selfless report from the night in patients with central hypersomnias. Conscious. Cogn. 2020, 81, 102931. [Google Scholar] [CrossRef]
- Krystal, A.D. Non-REM sleep EEG spectral analysis in insomnia. Psychiatr. Ann. 2008, 38, 615–620. [Google Scholar] [CrossRef]
- Ferrara, M.; Curcio, G.; Fratello, F.; Moroni, F.; Marzano, C.; Pellicciari, M.C.; Gennaro, L.; De Gennaro, L. The electroencephalographic substratum of the awakening. Behav. Brain Res. 2006, 167, 237–244. [Google Scholar] [CrossRef]
- Marzano, C.; Ferrara, M.; Moroni, F.; Gentile, B.; De Gennaro, L. P24.4 Electroencephalographic sleep inertia of the awakening brain. Clin. Neurophysiol. 2011, 122, S170. [Google Scholar] [CrossRef]
- Gorgoni, M.; Ferrara, M.; D’Atri, A.; Lauri, G.; Scarpelli, S.; Truglia, I.; De Gennaro, L. EEG topography during sleep inertia upon awakening after a period of increased homeostatic sleep pressure. Sleep Med. 2015, 16, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Voss, U.; Holzmann, R.; Hobson, A.; Paulus, W.; Koppehele-Gossel, J.; Klimke, A.; A Nitsche, M. Induction of self awareness in dreams through frontal low current stimulation of gamma activity. Nat. Neurosci. 2014, 17, 810–812. [Google Scholar] [CrossRef] [PubMed]
- Dodet, P.; Chávez, M.; Leu-Semenescu, S.; Golmard, J.-L.; Arnulf, I. Lucid Dreaming in Narcolepsy. Sleep 2015, 38, 487–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaskovich, B.; Reichardt, R.; Gombos, F.; I Spoormaker, V.; Simor, P. Cortical hyperarousal in NREM sleep normalizes from pre- to post- REM periods in individuals with frequent nightmares. Sleep 2019, 43. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| REC | NREC | t-Values | p-Values | |||
|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | |||
| Stage 1 latency (min) | 47.41 | 15.21 | 28.21 | 9.87 | 1.10 | 0.29 |
| Stage 2 latency (min) | 20.25 | 8.29 | 17.12 | 4.30 | 0.35 | 0.73 |
| Stage 3 latency (min) | 58.06 | 15.35 | 57.48 | 10.15 | 0.03 | 0.98 |
| REM sleep latency (min) | 132 | 26.14 | 88.72 | 13.18 | 1.56 | 0.14 |
| Stage 1 (min) | 14.45 | 3.29 | 14.36 | 2.86 | 0.02 | 0.99 |
| Stage 2 (min) | 235.11 | 11.38 | 244.21 | 11.14 | −0.57 | 0.58 |
| SWS (min) | 1.33 | 0.66 | 3.73 | 1.80 | −1.15 | 0.27 |
| REM (min) | 45.33 | 7.04 | 51.73 | 4.72 | −0.78 | 0.26 |
| WASO (min) | 72.25 | 16.58 | 53.75 | 9.54 | 1.01 | 0.33 |
| Awakenings (#) | 19.44 | 2.82 | 14.18 | 2.32 | 1.45 | 0.16 |
| TST (min) | 282.40 | 24.03 | 305.62 | 9.28 | −0.97 | 0.34 |
| TBT (min) | 388.70 | 13.94 | 374.18 | 10.87 | 0.83 | 0.42 |
| SEI% (TST/TBT) | 76.5 | 0.04 | 81.4 | 0.03 | −1.01 | 0.33 |
| Dream features | Mean | SE | |
| # DR | 1.67 | 0.24 | |
| Sr_EL | 3 | 0.71 | |
| Sr_VV | 3.33 | 0.76 | |
| Sr_B | 2.56 | 0.69 | |
| Sr_L | 2.11 | 0.59 | |
| EL | 1.89 | 0.39 | |
| VV | 4.22 | 0.32 | |
| B | 2.67 | 0.60 | |
| TWC | 83.78 | 41.48 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scarpelli, S.; D’Atri, A.; Bartolacci, C.; Gorgoni, M.; Mangiaruga, A.; Ferrara, M.; De Gennaro, L. Dream Recall upon Awakening from Non-Rapid Eye Movement Sleep in Older Adults: Electrophysiological Pattern and Qualitative Features. Brain Sci. 2020, 10, 343. https://doi.org/10.3390/brainsci10060343
Scarpelli S, D’Atri A, Bartolacci C, Gorgoni M, Mangiaruga A, Ferrara M, De Gennaro L. Dream Recall upon Awakening from Non-Rapid Eye Movement Sleep in Older Adults: Electrophysiological Pattern and Qualitative Features. Brain Sciences. 2020; 10(6):343. https://doi.org/10.3390/brainsci10060343
Chicago/Turabian StyleScarpelli, Serena, Aurora D’Atri, Chiara Bartolacci, Maurizio Gorgoni, Anastasia Mangiaruga, Michele Ferrara, and Luigi De Gennaro. 2020. "Dream Recall upon Awakening from Non-Rapid Eye Movement Sleep in Older Adults: Electrophysiological Pattern and Qualitative Features" Brain Sciences 10, no. 6: 343. https://doi.org/10.3390/brainsci10060343