P-cresol Alters Brain Dopamine Metabolism and Exacerbates Autism-Like Behaviors in the BTBR Mouse

 ,
,
Abstract
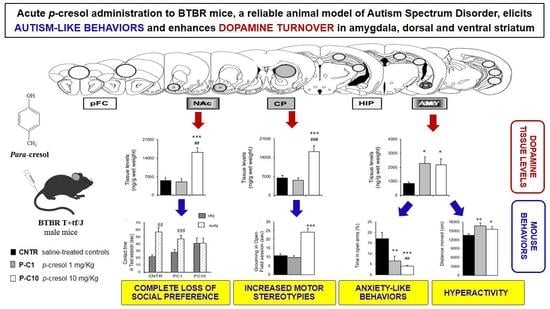
:
1. Background
2. Methods
2.1. Animals
2.2. P-cresol Treatment
2.3. Elevated Plus Maze
2.4. Open Field Test
2.5. Object Recognition Test
2.6. Three-chamber Social Interaction Test
2.7. Biochemical Assay
2.8. Statistical Analysis
3. Results
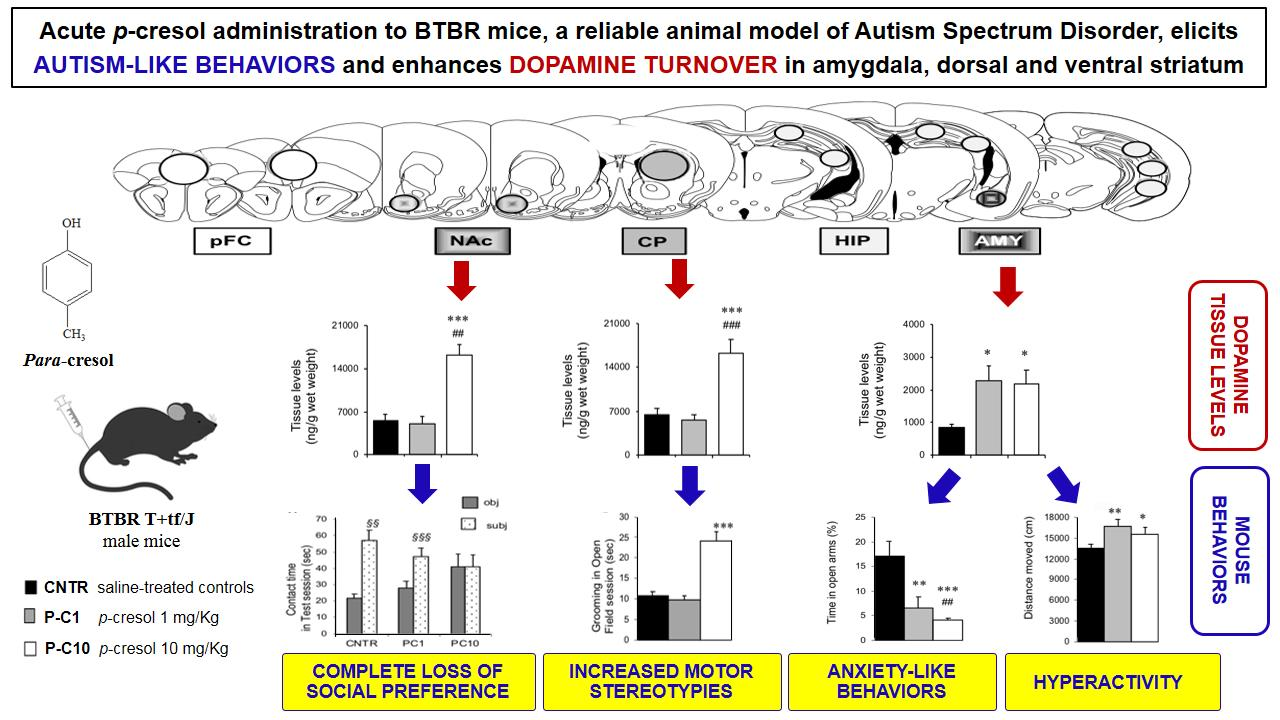
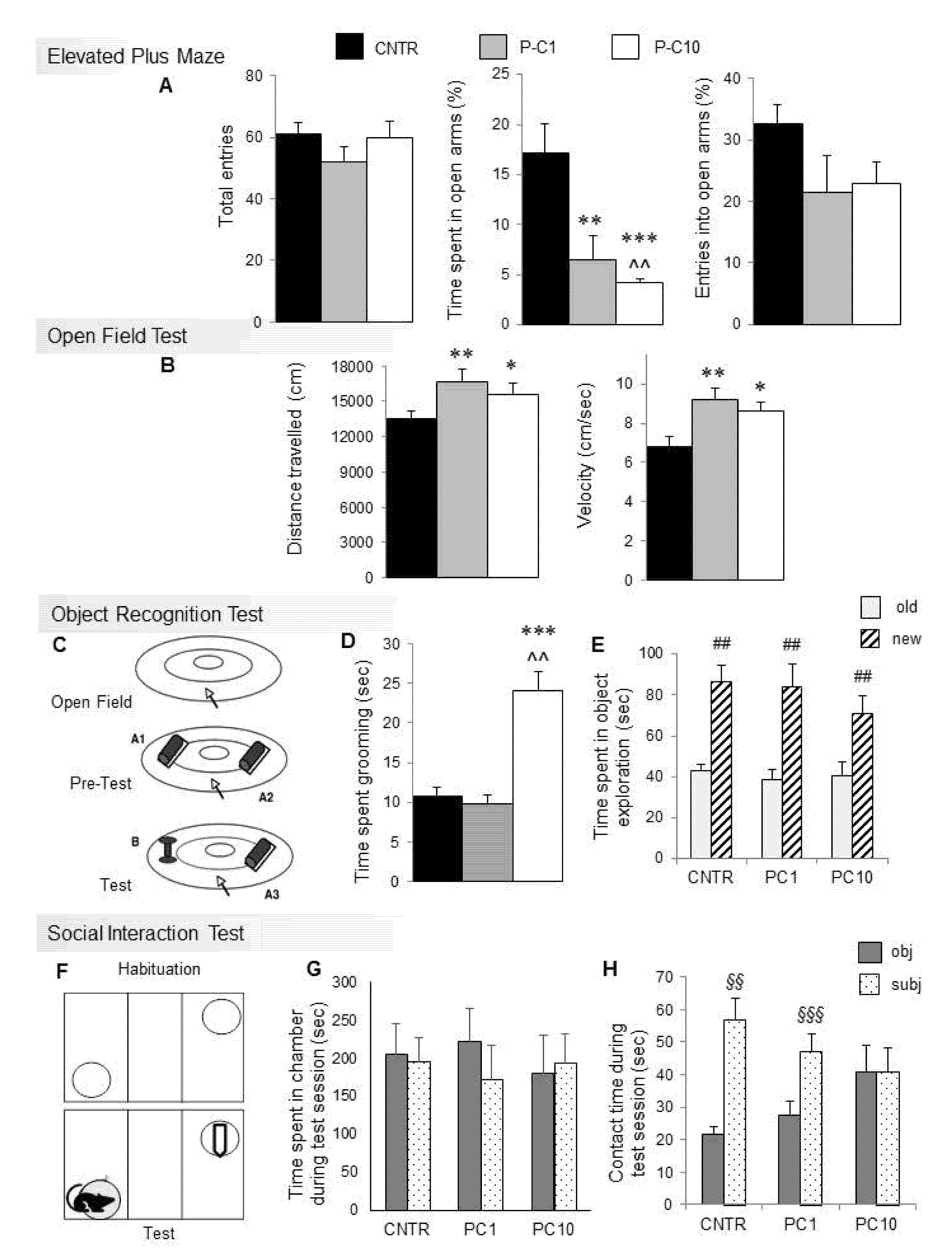
3.1. P-cresol Enhances Anxiety-like Behaviors in BTBR Mice
3.2. Locomotor Activity is Enhanced by p-cresol in the Open Field Test
3.3. P-cresol Enhances Motor Stereotypies without Modifying Object Recognition and Discrimination Behaviors
3.4. High Dose p-cresol Thwarts Preference for Social Interaction
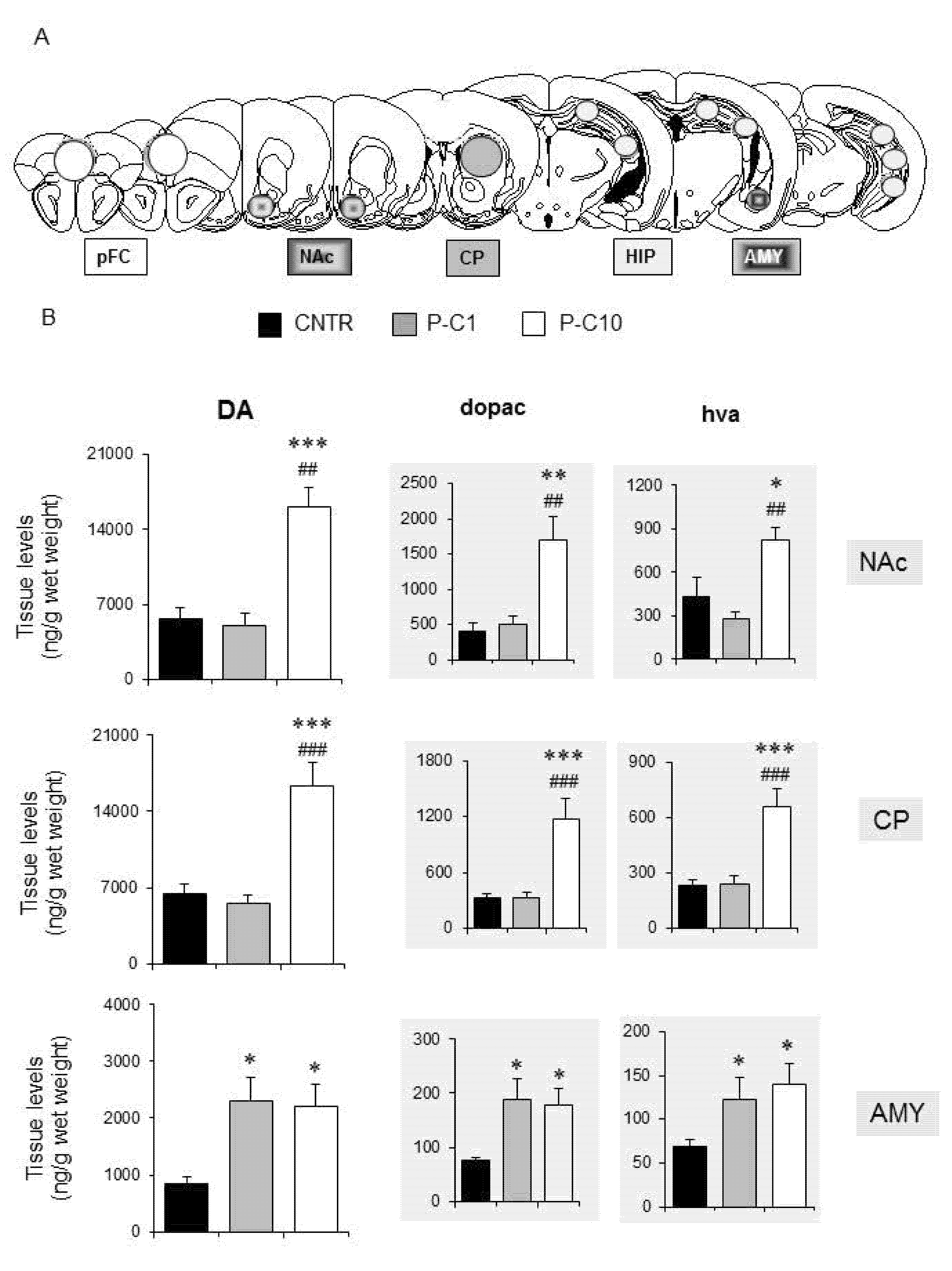
3.5. P-cresol Enhances Dopamine Metabolism in NAc, CP and AMY
4. Discussion
5. Limitations
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMY | Amygdala |
| ASD | Autism Spectrum Disorder; |
| CARS | Childhood Autism Rating Scale; |
| CNT | Control; |
| CP | Caudate Putamen; |
| DA | dopamine; |
| DOPAC | 3,4-Dihydroxyphenylacetic acid; |
| HIP | Hippocampus; |
| HVA | Homovanillic acid; |
| pFC | preFrontal Cortex; |
| NAc | Nucleus Accumbens; |
| NE | Norepinephrine; |
| 5-HIAA | 5-hydroxyindoleacetic acid; |
| 5-HT | Serotonin, |
| WT | Wild-type |
References
- Diagnostic and Statistical Manual of Mental Disorders: DSM-5; American Psychiatric Association: Arlington, VA, USA, 2013.
- Baio, J.; Wiggins, L.; Christensen, D.L.; Maenner, M.J.; Daniels, J.; Warren, Z.; Kurzius-Spencer, M.; Zahorodny, W.; Robinson Rosenberg, C.; White, T.; et al. Prevalence and Characteristics of Autism Spectrum Disorder Among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2012. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 1279. [Google Scholar]
- Baron-Cohen, S.; Scott, F.J.; Allison, C.; Williams, J.; Bolton, P.; Matthews, F.E.; Brayne, C. Prevalence of Autism-Spectrum Conditions: UK School-Based Population Study. Br. J. Psychiatry 2009, 194, 500–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fombonne, E. Epidemiology of Pervasive Developmental Disorders. Pediatric Res. 2009, 65, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Bourgeron, T. From the Genetic Architecture to Synaptic Plasticity in Autism Spectrum Disorder. Nat. Rev. Neurosci. 2015, 16, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Mandy, W.; Lai, M.-C. Annual Research Review: The Role of the Environment in the Developmental Psychopathology of Autism Spectrum Condition. J. Child Psychol. Psychiatry 2016, 57, 271–292. [Google Scholar] [CrossRef]
- Hertz-Picciotto, I.; Schmidt, R.J.; Krakowiak, P. Understanding Environmental Contributions to Autism: Causal Concepts and the State of Science. Autism Res. 2018, 11, 554–586. [Google Scholar] [CrossRef]
- Gottesman, I.I.; Shields, J. Genetic Theorizing and Schizophrenia. Br. J. Psychiatry 1973, 122, 15–30. [Google Scholar] [CrossRef]
- Persico, A.M.; Sacco, R. Endophenotypes in Autism Spectrum Disorders. In The Comprehensive Guide to Autism; Patel, V.B., Preedy, V.R., Martin, C.R., Eds.; Springer Science+Business Media: New York, NY, USA, 2014; pp. 77–96. [Google Scholar]
- Emond, P.; Mavel, S.; Aïdoud, N.; Nadal-Desbarats, L.; Montigny, F.; Bonnet-Brilhault, F.; Barthélémy, C.; Merten, M.; Sarda, P.; Laumonnier, F.; et al. GC-MS-Based Urine Metabolic Profiling of Autism Spectrum Disorders. Anal. Bioanal. Chem. 2013, 405, 5291–5300. [Google Scholar] [CrossRef] [Green Version]
- Gevi, F.; Zolla, L.; Gabriele, S.; Persico, A.M. Urinary Metabolomics of Young Italian Autistic Children Supports Abnormal Tryptophan and Purine Metabolism. Mol. Autism 2016, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Altieri, L.; Neri, C.; Sacco, R.; Curatolo, P.; Benvenuto, A.; Muratori, F.; Santocchi, E.; Bravaccio, C.; Lenti, C.; Saccani, M.; et al. Urinary p-Cresol Is Elevated in Small Children with Severe Autism Spectrum Disorder. Biomarkers 2011, 16, 252–260. [Google Scholar] [CrossRef]
- Gabriele, S.; Sacco, R.; Cerullo, S.; Neri, C.; Urbani, A.; Tripi, G.; Malvy, J.; Barthelemy, C.; Bonnet-Brihault, F.; Persico, A.M. Urinary p-Cresol Is Elevated in Young French Children with Autism Spectrum Disorder: A Replication Study. Biomarkers 2014, 19, 463–470. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Piccolo, M.; Vannini, L.; Siragusa, S.; Giacomo, A.D.; Serrazzanetti, D.I.; Cristofori, F.; Guerzoni, M.E.; Gobbetti, M.; Francavilla, R. Fecal Microbiota and Metabolome of Children with Autism and Pervasive Developmental Disorder Not Otherwise Specified. PLoS ONE 2013, 8, e76993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, D.-W.; Ilhan, Z.E.; Isern, N.G.; Hoyt, D.W.; Howsmon, D.P.; Shaffer, M.; Lozupone, C.A.; Hahn, J.; Adams, J.B.; Krajmalnik-Brown, R. Differences in Fecal Microbial Metabolites and Microbiota of Children with Autism Spectrum Disorders. Anaerobe 2018, 49, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Gabriele, S.; Sacco, R.; Altieri, L.; Neri, C.; Urbani, A.; Bravaccio, C.; Riccio, M.P.; Iovene, M.R.; Bombace, F.; Magistris, L.D.; et al. Slow Intestinal Transit Contributes to Elevate Urinary p-Cresol Level in Italian Autistic Children. Autism Res. 2016, 9, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Persico, A.M.; Napolioni, V. Urinary p-Cresol in Autism Spectrum Disorder. Neurotoxicol. Teratol. 2013, 36, 82–90. [Google Scholar] [CrossRef]
- Yehuda, S.; Carasso, R.L.; Mostofsky, D.I. Essential Fatty Acid Preparation (SR-3) Raises the Seizure Threshold in Rats. Eur. J. Pharmacol. 1994, 254, 193–198. [Google Scholar] [CrossRef]
- Calderón-Guzmán, D.; Hernández-Islas, J.L.; Vázquez, I.R.E.; Barragán-Mejía, G.; Hernández-García, E.; Angel, D.S.D.; Juárez-Olguín, H. Effect of Toluene and Cresols on Na, K-ATPase, and Serotonin in Rat Brain. Regul. Toxicol. Pharmacol. 2005, 41, 1–5. [Google Scholar] [CrossRef]
- Goodhart, P.J.; Dewolf, W.E.; Kruse, L.I. Mechanism-Based Inactivation of Dopamine. Beta.-Hydroxylase by p-Cresol and Related Alkylphenols. Biochemistry 1987, 26, 2576–2583. [Google Scholar] [CrossRef]
- Patterson, P.H. Modeling Autistic Features in Animals. Pediatric Res. 2011, 69 Pt 2, 34R–40R. [Google Scholar] [CrossRef]
- Crawley, J.N. Translational animal models of autism and neurodevelopmental disorders. Dialogues. Clin. Neurosci. 2012, 14, 293–305. [Google Scholar]
- Silverman, J.L.; Yang, M.; Lord, C.; Crawley, J.N. Behavioural Phenotyping Assays for Mouse Models of Autism. Nat. Rev. Neurosci. 2010, 11, 490–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courchesne, E.; Pramparo, T.; Gazestani, V.H.; Lombardo, M.V.; Pierce, K.; Lewis, N.E. The ASD Living Biology: From Cell Proliferation to Clinical Phenotype. Mol. Psychiatry 2019, 24, 88–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaral, D.G.; Schumann, C.M.; Nordahl, C.W. Neuroanatomy of Autism. Trends Neurosci. 2008, 31, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Schubert, D.; Martens, G.J.M.; Kolk, S.M. Molecular Underpinnings of Prefrontal Cortex Development in Rodents Provide Insights into the Etiology of Neurodevelopmental Disorders. Mol. Psychiatry 2015, 20, 795–809. [Google Scholar] [CrossRef] [Green Version]
- Meyza, K.Z.; Defensor, E.B.; Jensen, A.L.; Corley, M.J.; Pearson, B.L.; Pobbe, R.L.; Bolivar, V.J.; Blanchard, D.C.; Blanchard, R.J. The BTBR T Tf/J Mouse Model for Autism Spectrum Disorders–in Search of Biomarkers. Behav. Brain Res. 2013, 251, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Ellegood, J.; Crawley, J.N. Behavioral and Neuroanatomical Phenotypes in Mouse Models of Autism. Neurotherapeutics 2015, 12, 521–533. [Google Scholar] [CrossRef]
- Pascucci, T.; Giacovazzo, G.; Andolina, D.; Accoto, A.; Fiori, E.; Ventura, R.; Orsini, C.; Conversi, D.; Carducci, C.; Leuzzi, V.; et al. Behavioral and Neurochemical Characterization of New Mouse Model of Hyperphenylalaninemia. PLoS ONE 2013, 8, e84697. [Google Scholar] [CrossRef] [Green Version]
- Nadler, J.J.; Moy, S.S.; Dold, G.; Simmons, N.; Perez, A.; Young, N.B.; Barbaro, R.P.; Piven, J.; Magnuson, T.R.; Crawley, J.N. Automated Apparatus for Quantitation of Social Approach Behaviors in Mice. GenesBrain Behav. 2004, 3, 303–314. [Google Scholar] [CrossRef]
- Fiori, E.; Oddi, D.; Ventura, R.; Colamartino, M.; Valzania, A.; D’Amato, F.R.; Bruinenberg, V.; Zee, E.V.D.; Puglisi-Allegra, S.; Pascucci, T. Early-Onset Behavioral and Neurochemical Deficits in the Genetic Mouse Model of Phenylketonuria. PLoS ONE 2017, 12, e0183430. [Google Scholar] [CrossRef] [Green Version]
- Pascucci, T.; Rossi, L.; Colamartino, M.; Gabucci, C.; Carducci, C.; Valzania, A.; Sasso, V.; Bigini, N.; Pierigè, F.; Viscomi, M.T.; et al. A New Therapy Prevents Intellectual Disability in Mouse with Phenylketonuria. Mol. Genet. Metab. 2018, 124, 39–49. [Google Scholar] [CrossRef]
- Puglisi-Allegra, S.; Cabib, S.; Pascucci, T.; Ventura, R.; Cali, F.; Romano, V. Dramatic Brain Aminergic Deficit in a Genetic Mouse Model of Phenylketonuria. NeuroReport 2000, 11, 1361–1364. [Google Scholar] [CrossRef] [PubMed]
- Moy, S.; Nadler, J.; Young, N.; Perez, A.; Holloway, L.; Barbaro, R.; Barbaro, J.; Wilson, L.; Threadgill, D.; Lauder, J. Mouse Behavioral Tasks Relevant to Autism: Phenotypes of 10 Inbred Strains. Behav. Brain Res. 2007, 176, 4–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berenguer-Forner, C.; Miranda-Casas, A.; Pastor-Cerezuela, G.; Roselló-Miranda, R. Comorbidity of autism spectrum disorder and attention deficit with hyperactivity. A review study. Rev. De Neurol. 2015, 60 (Suppl. 1), S37–S43. [Google Scholar]
- Van Steensel, F.J.A.; Bögels, S.M.; Perrin, S. Anxiety Disorders in Children and Adolescents with Autistic Spectrum Disorders: A Meta-Analysis. Clin. Child Fam. Psychol. Rev. 2011, 14, 302–317. [Google Scholar] [CrossRef] [Green Version]
- Bales, K.L.; Solomon, M.; Jacob, S.; Crawley, J.N.; Silverman, J.L.; Larke, R.H.; Sahagun, E.; Puhger, K.R.; Pride, M.C.; Mendoza, S.P. Long-Term Exposure to Intranasal Oxytocin in a Mouse Autism Model. Transl. Psychiatry 2014, 4, e480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segal-Gavish, H.; Karvat, G.; Barak, N.; Barzilay, R.; Ganz, J.; Edry, L.; Aharony, I.; Offen, D.; Kimchi, T. Mesenchymal Stem Cell Transplantation Promotes Neurogenesis and Ameliorates Autism Related Behaviors in BTBR Mice. Autism Res. 2016, 9, 17–32. [Google Scholar] [CrossRef]
- Yang, M.; Abrams, D.N.; Zhang, J.Y.; Weber, M.D.; Katz, A.M.; Clarke, A.M.; Silverman, J.L.; Crawley, J.N. Low Sociability in BTBR T Tf/J Mice Is Independent of Partner Strain. Physiol. Behav. 2012, 107, 649–662. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, H.G.; Kusek, G.K.; Yang, M.; Phoenix, J.L.; Bolivar, V.J.; Crawley, J.N. Autism-like Behavioral Phenotypes in BTBR T Tf/J Mice. GenesBrain Behav. 2008, 7, 152–163. [Google Scholar] [CrossRef]
- Silverman, J.; Oliver, C.; Karras, M.; Gastrell, P.; Crawley, J. AMPAKINE Enhancement of Social Interaction in the BTBR Mouse Model of Autism. Neuropharmacology 2013, 64, 268–282. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.Q.; Smolik, C.M.; Barba-Escobedo, P.A.; Gamez, M.; Sanchez, J.J.; Javors, M.A.; Daws, L.C.; Gould, G.G. Acute Dietary Tryptophan Manipulation Differentially Alters Social Behavior, Brain Serotonin and Plasma Corticosterone in Three Inbred Mouse Strains. Neuropharmacology 2015, 90, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Cabib, S.; Pascucci, T.; Ventura, R.; Romano, V.; Puglisi-Allegra, S. The behavioral profile of severe mental retardation in a genetic mouse model of Phenylketonuria, Behav. Genet. 2003, 33, 301–310. [Google Scholar]
- Andolina, D.; Conversi, D.; Cabib, S.; Trabalza, A.; Ventura, R.; Puglisi-Allegra, S.; Pascucci, T. 5-Hydroxytryptophan during postnatal period improves cognitive performances and promotes dendritic spine maturation in genetic mouse model of Phenylketonuria. Int. J. Neuropsychopharmacol. 2010, 14, 479–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pobbe, R.L.H.; Defensor, E.B.; Pearson, B.L.; Blanchard, V.J.C.; Blancharda, R.J. General and social anxiety in the BTBR T+ tf/J mouse strain Behav. Brain Res. 2011, 216, 446–451. [Google Scholar]
- Bruinenberg, V.M.; van der Goot, E.; van Vliet, D.; de Groot, M.J.; Mazzola, P.N.; Heiner-Fokkema, M.R.; van Faassen, M.; van Spronsen, F.J.; van der Zee, E.A. The behavioral consequence of Phenylketonuria in mice depends on the genetic background. Front. Behav. Neurosci. 2016, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Zilkha, N.; Kuperman, Y.; Kimchi, T. High-fat diet exacerbates cognitive rigidity and social deficiency in the BTBR mouse model of autism. Neuroscience 2017, 345, 142–154. [Google Scholar] [CrossRef]
- Schindler, C.; Persico, A.; Uhl, G.; Goldberg, S. Behavioral Assessment of High-Dose Amphetamine Withdrawal: Importance of Training and Testing Conditions. Pharmacol. Biochem. Behav. 1994, 49, 41–46. [Google Scholar] [CrossRef]
- Graybiel, A.M. Habits, Rituals, and the Evaluative Brain. Annu. Rev. Neurosci. 2008, 31, 359–387. [Google Scholar] [CrossRef] [Green Version]
- Campbell, N.G.; Shekar, A.; Aguilar, J.I.; Peng, D.; Navratna, V.; Yang, D.; Morley, A.N.; Duran, A.M.; Galli, G.; O’Grady, B.; et al. Structural, functional, and behavioral insights of dopamine dysfunction revealed by a deletion in SLC6A3. Proc. Natl. Acad. Sci. USA. 2019, 116, 3853–3862. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Sato, A.; Kasai, S.; Hagino, Y.; Kotajima-Murakami, H.; Kashii, H.; Takamatsu, Y.; Nishito, Y.; Inagaki, M.; Mizuguchi, M.; et al. Brain hyperserotonemia causes autism-relevant social deficits in mice. Mol. Autism 2018, 9, 60. [Google Scholar] [CrossRef]
- Pavăl, D. A Dopamine Hypothesis of Autism Spectrum Disorder. Dev. Neurosci. 2017, 39, 355–360. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, H.; Kim, J.-E.; Park, J.-Y.; Choi, J.; Lee, J.-E.; Lee, E.-H.; Han, P.-L. Excessive D1 Dopamine Receptor Activation in the Dorsal Striatum Promotes Autistic-Like Behaviors. Mol. Neurobiol. 2018, 55, 5658–5671. [Google Scholar] [CrossRef] [PubMed]
- Squillace, M.; Dodero, L.; Federici, M.; Migliarini, S.; Errico, F.; Napolitano, F.; Krashia, P.; Di Maio, A.; Galbusera, A.; Bifone, A.; et al. Dysfunctional dopaminergic neurotransmission in asocial BTBR mice. Transl. Psychiatry 2014, 4, e427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohls, G.; Schulte-Rüther, M.; Nehrkorn, B.; Müller, K.; Fink, G.R.; Kamp-Becker, I.; Herpertz-Dahlmann, B.; Schultz, R.T.; Konrad, K. Reward system dysfunction in autism spectrum disorders. Soc. Cogn. Affect. Neurosci. 2013, 8, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, L.M.; Rudie, J.D.; Green, S.A.; Bookheimer, S.; Dapretto, M. Neural Signatures of Autism Spectrum Disorders: Insights into Brain Network Dynamics. Neuropsychopharmacology 2015, 40, 171–189. [Google Scholar] [CrossRef] [Green Version]
- Dichter, G.S.; Felder, J.N.; Green, S.R.; Rittenberg, A.M.; Sasson, N.J.; Bodfish, J.W. Reward Circuitry Function in Autism Spectrum Disorders. Soc. Cogn. Affect. Neurosci. 2012, 7, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Sasson, N.J.; Dichter, G.S.; Bodfish, J.W. Affective Responses by Adults with Autism Are Reduced to Social Images but Elevated to Images Related to Circumscribed Interests. PLoS ONE 2012, 7, e42457. [Google Scholar] [CrossRef] [Green Version]
- Stavropoulos, K.K.; Carver, L.J. Reward Anticipation and Processing of Social versus Nonsocial Stimuli in Children with and without Autism Spectrum Disorders. J. Child Psychol. Psychiatry 2014, 55, 1398–1408. [Google Scholar] [CrossRef] [Green Version]
- Russo, S.J.; Dietz, D.M.; Dumitriu, D.; Morrison, J.H.; Malenka, R.C.; Nestler, E.J. The Addicted Synapse: Mechanisms of Synaptic and Structural Plasticity in Nucleus Accumbens. Trends Neurosci. 2010, 33, 267–276. [Google Scholar] [CrossRef] [Green Version]
- MacFabe, D.F. Short-Chain Fatty Acid Fermentation Products of the Gut Microbiome: Implications in Autism Spectrum Disorders. Microb. Ecol. Health Dis. 2012, 23, 19260. [Google Scholar] [CrossRef]
- Rubenstein, J.L.R.; Merzenich, M.M. Model of Autism: Increased Ratio of Excitation/Inhibition in Key Neural Systems. Genes Brain Behav. 2003, 2, 255–267. [Google Scholar] [CrossRef]
- Gabriele, S.; Sacco, R.; Persico, A.M. Blood Serotonin Levels in Autism Spectrum Disorder: A Systematic Review and Meta-Analysis. Eur. Neuropsychopharmacol. 2014, 24, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Pagan, C.; Delorme, R.; Callebert, J.; Goubran-Botros, H.; Amsellem, F.; Drouot, X.; Boudebesse, C.; Dudal, K.L.; Ngo-Nguyen, N.; Laouamri, H.; et al. The Serotonin-N-Acetylserotonin–Melatonin Pathway as a Biomarker for Autism Spectrum Disorders. Transl. Psychiatry 2014, 4, e479. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.W.; Adams, J.B.; Coleman, D.M.; Pollard, E.L.; Maldonado, J.; Mcdonough-Means, S.; Caporaso, J.G.; Krajmalnik-Brown, R. Long-Term Benefit of Microbiota Transfer Therapy on Autism Symptoms and Gut Microbiota. Sci. Rep. 2019, 9, 5821. [Google Scholar] [CrossRef]
- Dölen, G. Autism: Oxytocin, Serotonin, and Social Reward. Soc. Neurosci. 2015, 10, 450–465. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| DA | DOPAC | HVA | NE | 5HT | HIAA | ||
|---|---|---|---|---|---|---|---|
| pFC | CNTR | 342.95 ± 99.78 | 44.96 ± 9.21 | 43.95 ± 7.91 | 179.12 ± 26.24 | 826.28 ± 116.29 | 266.67 ± 51.36 |
| P-C1 | 305.53 ± 57.11 | 43.79 ± 11.19 | 37.07 ± 6.91 | 132.30 ± 25.35 | 607.64 ± 74.63 | 152.62 ± 15.13 | |
| P-C10 | 423.87 ± 138.8 | 76.9 ± 24.84 | 42.91 ± 14.9 | 139.42 ± 36.39 | 881.03 ± 207.31 | 171.06 ± 39.93 | |
| HIPP | CNTR | 155.18 ± 15.24 | 41.73 ± 15.71 | 29.98 ± 2.79 | n.d. | 641.22 ± 173.86 | 378.98 ± 59.82 |
| P-C1 | 113.92 ± 26.27 | 21.54 ± 7.92 | 22.77 ± 3.72 | n.d. | 425.17 ± 92.36 | 327.61 ± 102.74 | |
| P-C10 | 119.47 ± 40.91 | 37.29 ± 12.97 | 30.65 ± 7.65 | n.d. | 365.6 ± 145.62 | 314.2 ± 99.42 | |
| AMY | CNTR | 858.36 ± 112.78 | 68.98 ± 6.62 | 57.41 ± 11.05 | 274.34 ± 103.69 | 193.46 ± 42.02 | 149.84 ± 55.3 |
| P-C1 | 2292.71 ± 526.75 * | 187.95 ± 38.11 * | 122.05 ± 26.41 * | 314.18 ± 89.95 | 356.62 ± 109.27 | 212.38 ± 88.17 | |
| P-C10 | 2197.45 ± 992.31 * | 179.38 ± 30.12 * | 140.21 ± 23.75 * | 283.15 ± 48.49 | 330.85 ± 59.27 | 226.03 ± 36.56 | |
| CP | CNTR | 5284.18 ± 1015.8 | 584.97 ± 186.65 | 251.81 ± 33.64 | 35.46 ± 8.61 | 159.61 ± 31.77 | 133.99 ± 28.56 |
| P-C1 | 5499.14 ± 842.38 | 327.32 ± 56.27 | 240.48 ± 43.35 | 46.41 ± 8.61 | 183.88 ± 31.09 | 130.22 ± 38.39 | |
| P-C10 | 16270.59 ± 2153.37 ***,### | 1176.26 ± 223.34 ***,### | 658.32 ± 97.93 ***,### | 63.11 ± 12.83 | 341.1 ± 34.82 **,## | 242.73 ± 49.04 | |
| NAc | CNTR | 5623.89 ± 1050.64 | 506.55 ± 91.15 | 412.55 ± 87.32 | 1214.04 ± 390.84 | 1191.69 ± 355.05 | 459.72 ± 126.19 |
| P-C1 | 5035.05 ± 1134.2 | 504.3 ± 119.73 | 276.01 ± 49.36 | 1359.63 ± 200.81 | 1232.82 ± 357.02 | 305.25 ± 63.12 | |
| P-C10 | 16156.11 ± 1812.97 **,## | 1698.82 ± 325.04 **,## | 817.35 ± 91.08 *,## | 1883.69 ± 626.27 | 1360.47 ± 512.3 | 480.62 ± 189.73 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pascucci, T.; Colamartino, M.; Fiori, E.; Sacco, R.; Coviello, A.; Ventura, R.; Puglisi-Allegra, S.; Turriziani, L.; Persico, A.M. P-cresol Alters Brain Dopamine Metabolism and Exacerbates Autism-Like Behaviors in the BTBR Mouse. Brain Sci. 2020, 10, 233. https://doi.org/10.3390/brainsci10040233
Pascucci T, Colamartino M, Fiori E, Sacco R, Coviello A, Ventura R, Puglisi-Allegra S, Turriziani L, Persico AM. P-cresol Alters Brain Dopamine Metabolism and Exacerbates Autism-Like Behaviors in the BTBR Mouse. Brain Sciences. 2020; 10(4):233. https://doi.org/10.3390/brainsci10040233
Chicago/Turabian StylePascucci, Tiziana, Marco Colamartino, Elena Fiori, Roberto Sacco, Annalisa Coviello, Rossella Ventura, Stefano Puglisi-Allegra, Laura Turriziani, and Antonio M. Persico. 2020. "P-cresol Alters Brain Dopamine Metabolism and Exacerbates Autism-Like Behaviors in the BTBR Mouse" Brain Sciences 10, no. 4: 233. https://doi.org/10.3390/brainsci10040233