Post-Encoding Stress Does Not Enhance Memory Consolidation: The Role of Cortisol and Testosterone Reactivity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedure
2.2.1. Session 1: Acquisition Session
2.2.2. Session 2: Memory Testing Session
2.3. Anxiety
2.4. Hormonal Determination
2.5. Statistical Analyses
3. Results
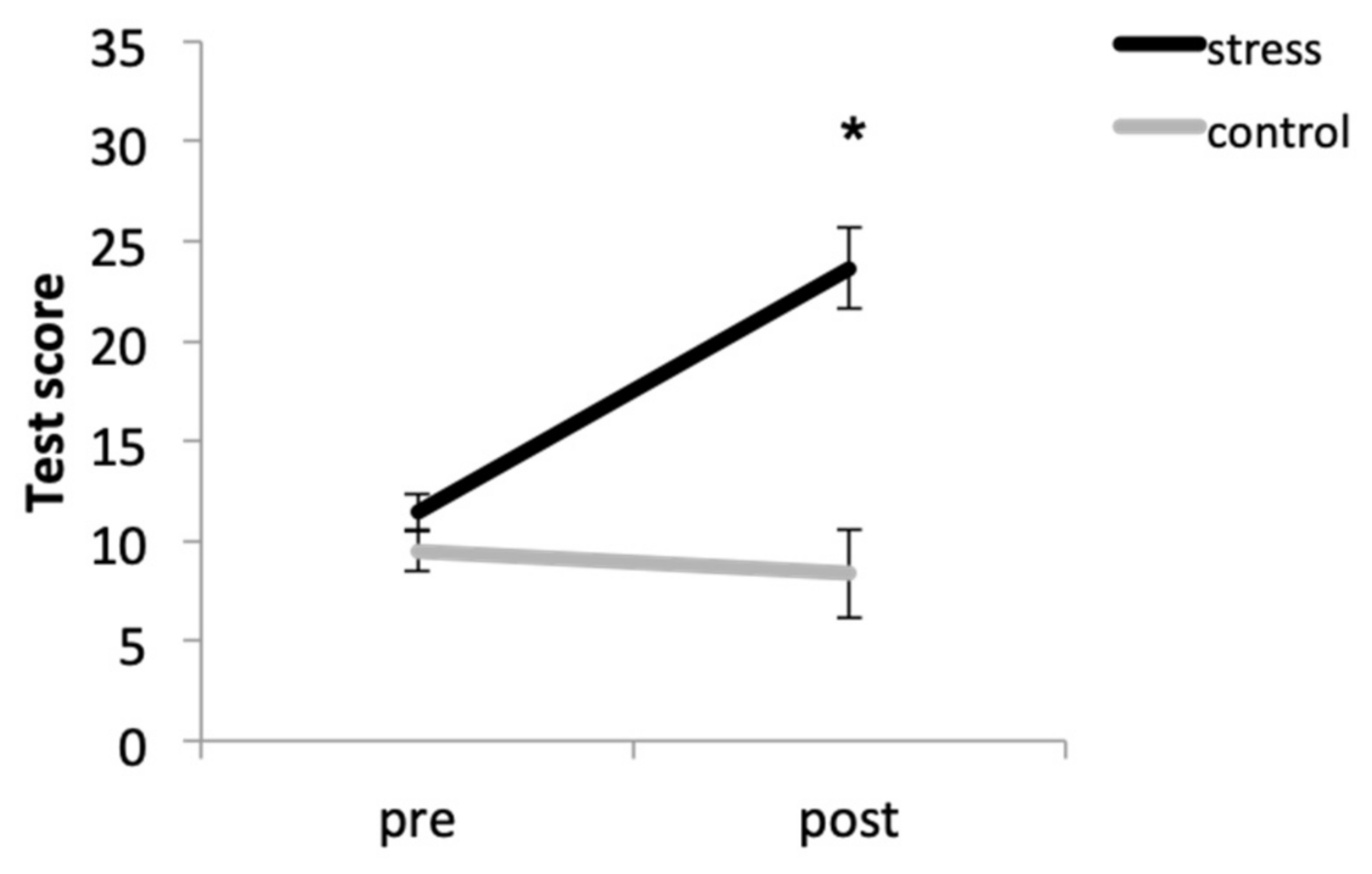
3.1. Anxiety Self-Reported
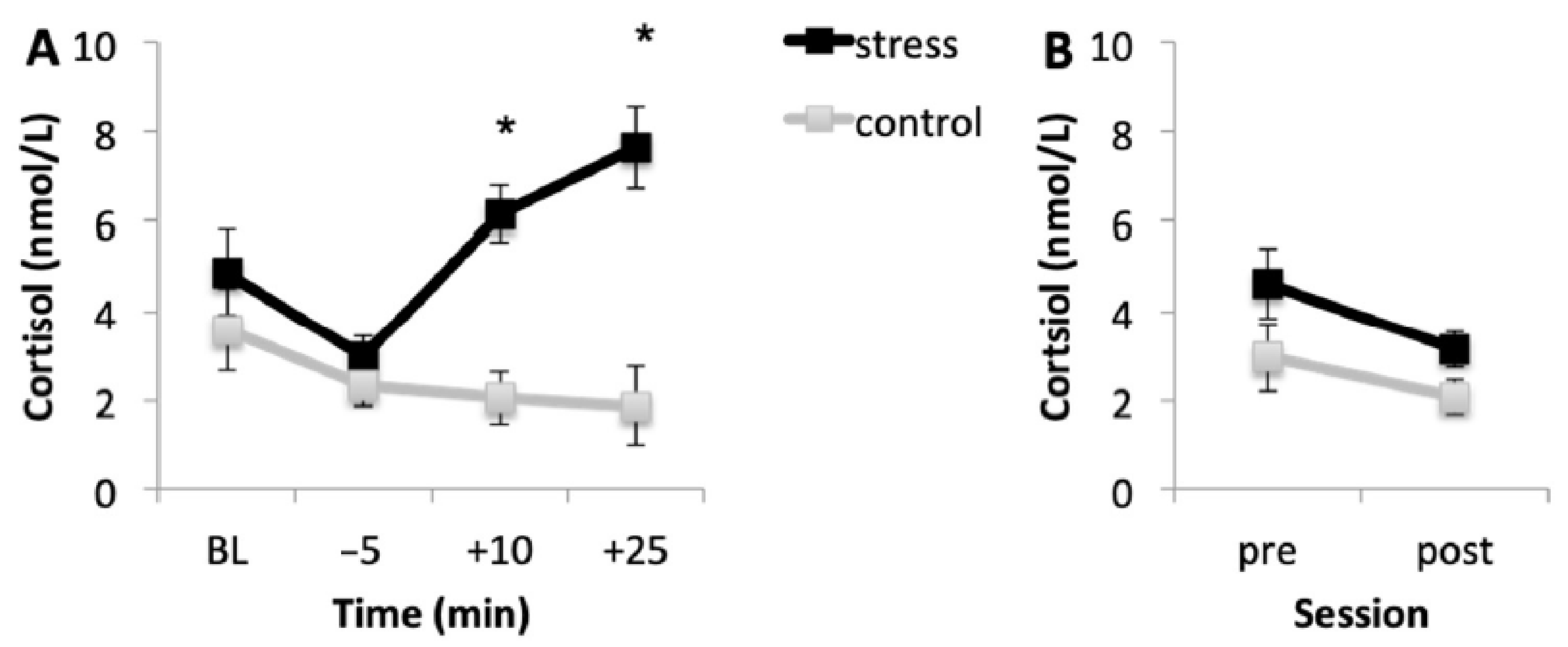
3.2. Cortisol Response
3.2.1. Acquisition Session
3.2.2. Memory Testing Session
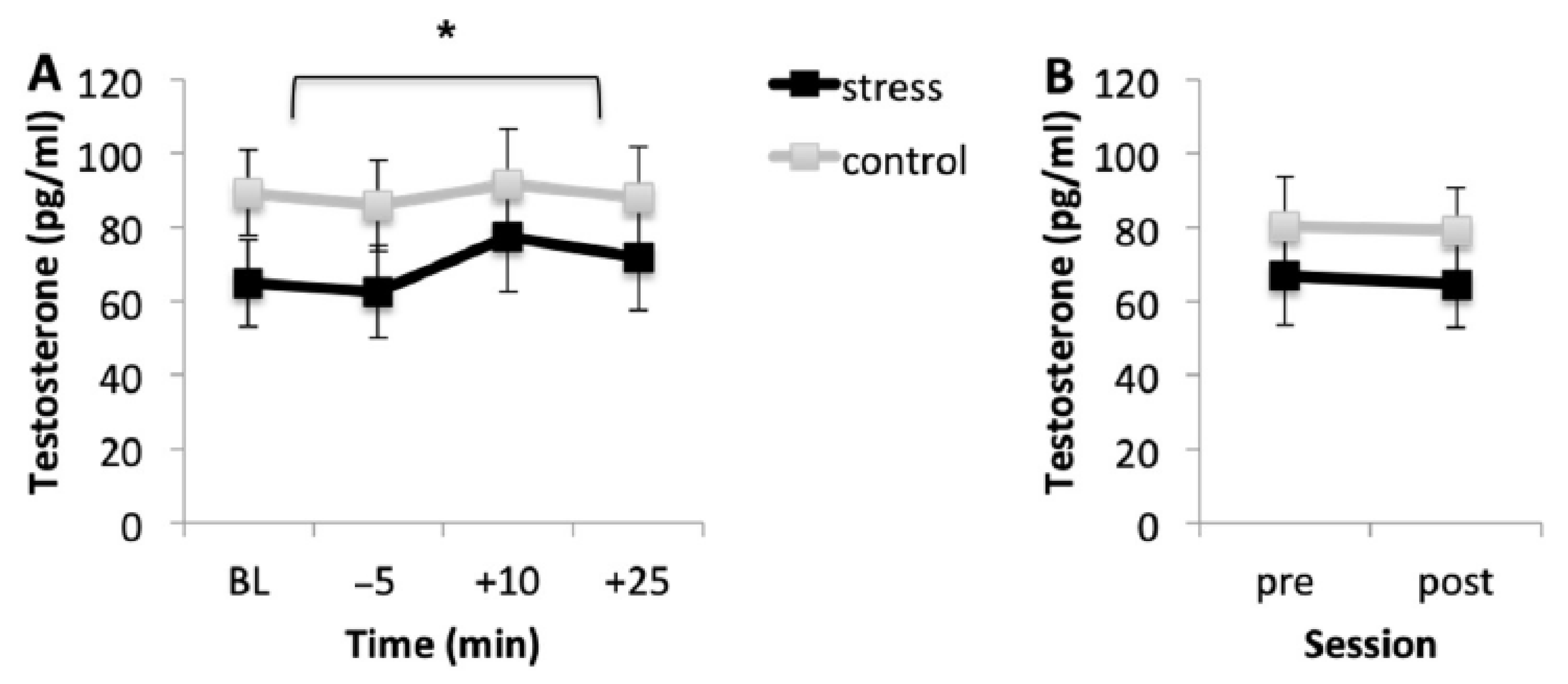
3.3. Testosterone Response
3.3.1. Acquisition Session
3.3.2. Memory Testing Session
3.4. Ratings of Picture Material
3.4.1. Valence
3.4.2. Arousal
3.5. Memory Performance
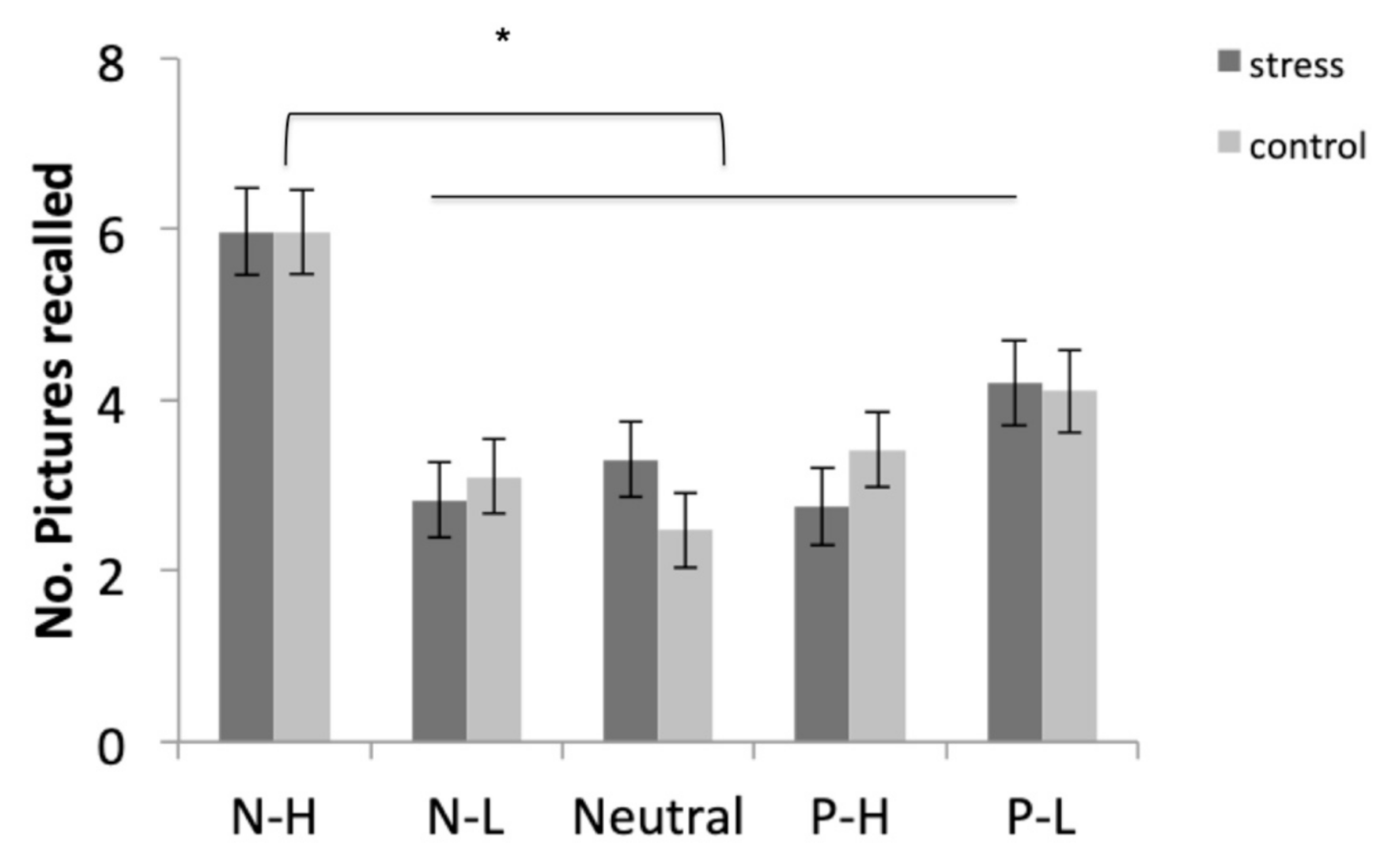
3.5.1. Free Recall
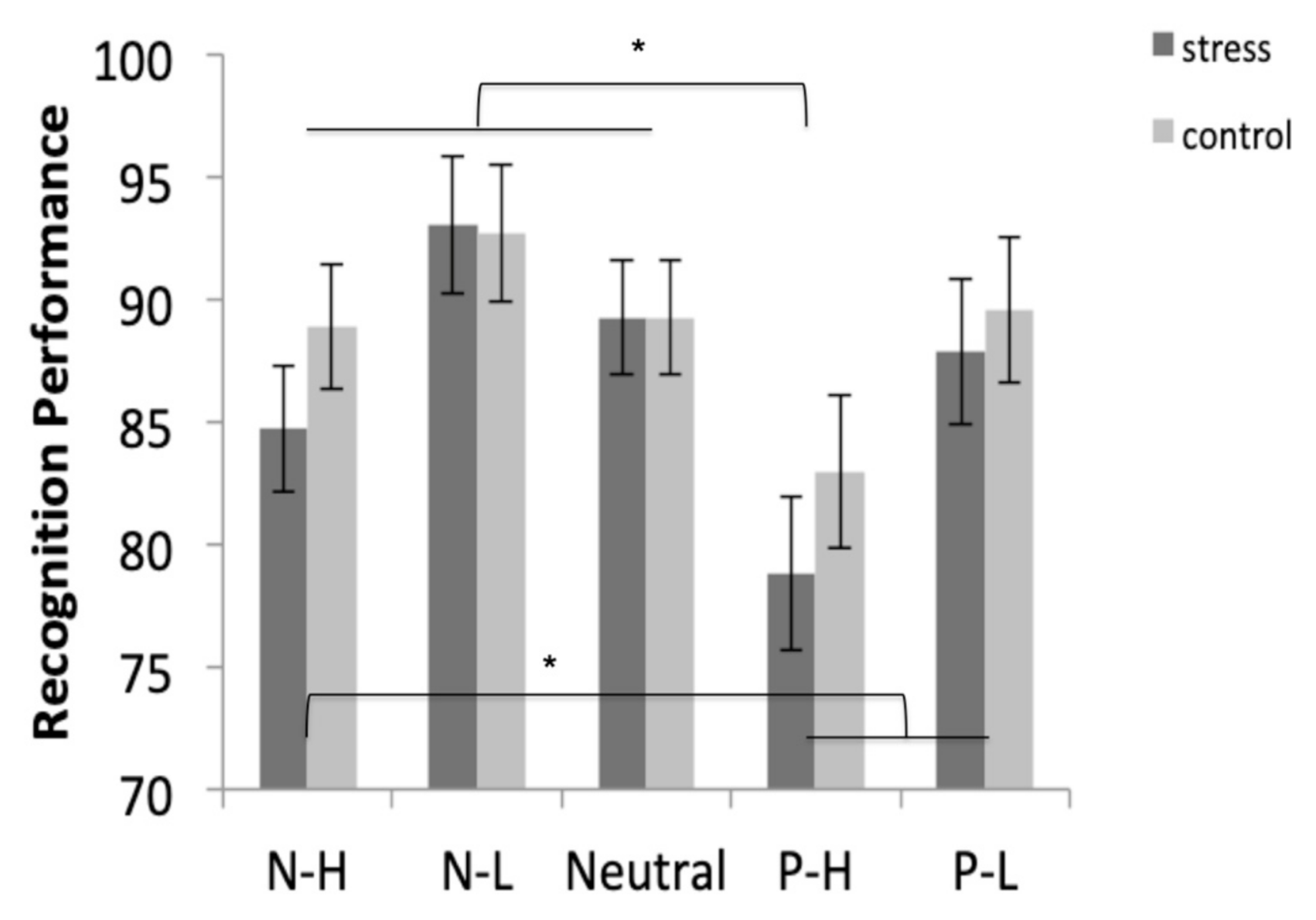
3.5.2. Recognition
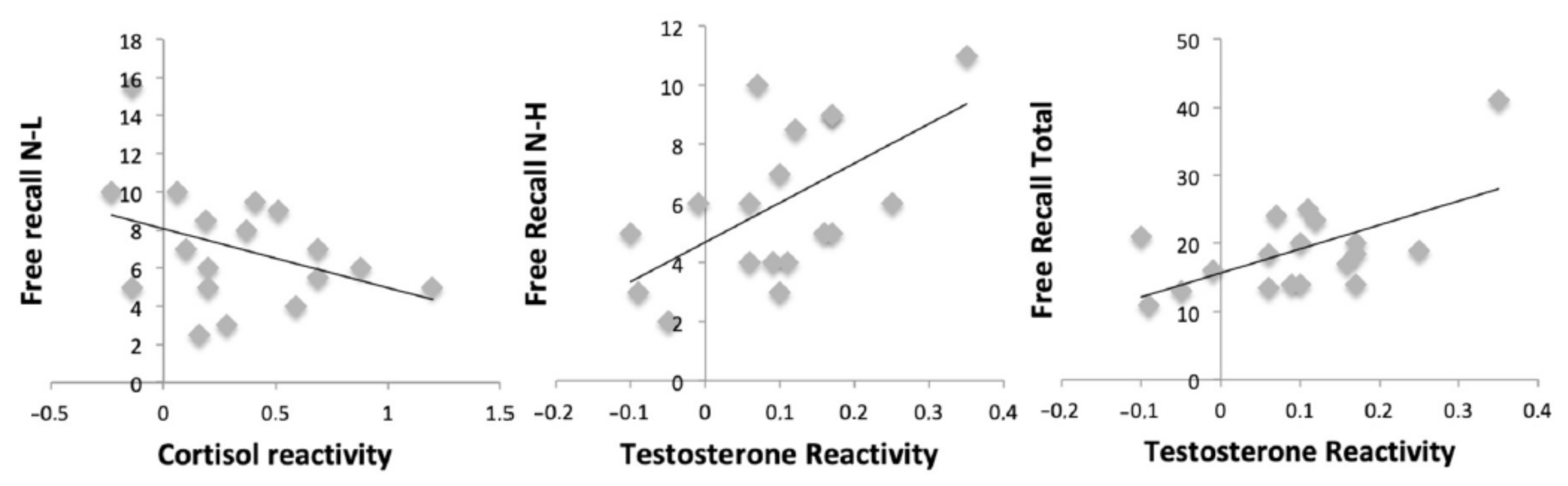
3.6. The Relationship between the Hormonal Response and Consolidation Memory Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sandi, C. Stress and cognition. WIREs Cogn. Sci. 2013, 4, 245–261. [Google Scholar] [CrossRef]
- Hidalgo, V.; Pulopulos, M.M.; Salvador, A. Acute psychosocial stress effects on memory performance: Relevance of age and sex. Neurobiol. Learn. Mem. 2019, 157, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Ulrich-Lai, Y.M.; Herman, J.P. Neural regulation of endocrine and autonomic stress responses. Nat. Rev. Neurosci. 2009, 10, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roozendaal, B.; McEwen, B.S.; Chattarji, S. Stress, memory and the amgydala. Nat. Rev. Neurosci. 2009, 10, 423–433. [Google Scholar] [CrossRef]
- Roozendaal, B. Glucocorticoids on memory consolidation and memory retrieval. Neurobiol. Learn. Mem. 2002, 78, 578–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, L.; Gorski, L.; Le, K. Enhanced human memory consolidation with post-learning stress: Interaction with the degree of arousal at encoding. Learn. Mem. 2003, 10, 270–274. [Google Scholar] [CrossRef] [Green Version]
- Andreano, J.A.; Cahill, L. Glucocorticoid release and memory consolidation in men and women. Psychol. Sci. 2006, 17, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Beckner, V.E.; Tucker, D.M.; Delville, Y.; Mohr, D. Stress facilitates consolidation of verbal memory for a film but does not affect retrieval. Behav. Neurosci. 2006, 120, 518–527. [Google Scholar] [CrossRef]
- Smeets, T.; Otgaar, H.; Candel, I.; Wolf, O.T. True or false? Memory is differentially affected by stress-induced cortisol elevations and sympathetic activity at consolidation and retrieval. Psychoneuroendocrinology 2008, 33, 1378–1386. [Google Scholar] [CrossRef]
- Preuss, D.; Wolf, O.T. Post-learning psychosocial stress enhances consolidation of neutral stimuli. Neurobiol. Learn. Mem. 2009, 318–326. [Google Scholar] [CrossRef]
- Felmingham, K.L.; Fong, W.C.; Bryant, R.A. The impact of progesterone on memory consolidation of threatening images in women. Psychoneuroendocrinology 2012, 37, 1896–1900. [Google Scholar] [CrossRef] [PubMed]
- Felmingham, K.L.; Tran, T.P.; Fong, W.C.; Bryant, R.A. Sex differences in emotional memory consolidation: The effect of stress-induced salivary alpha-amylase and cortisol. Biol. Psychol. 2012, 89, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Bryant, R.A.; McGrath, C.; Felmingham, K.L. The roles of noradrenergic and glucocorticoid activation in the development of intrusive memories. PLoS ONE 2013, 8, e62675. [Google Scholar] [CrossRef] [PubMed]
- McCullough, A.M.; Ritchey, M.; Ranganath, C.; Yonelinas, A. Differential effects of stress-induced cortisol responses on recollection and familiarity-based recognition memory. Neurobiol. Learn. Mem. 2015, 123, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Shileds, G.S.; Rivers, A.M.; Ramey, M.M.; Trainor, B.C.; Yonelinas, A.P. Mild acute stress improves response speed without impairing accuracy or interference control in two selective attention tasks: Implications for theories of stress and cognition. Psychoneuroendocrinology 2019, 108, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Zoladz, P.R.; Peters, D.M.; Cadle, C.E.; Kalchik, A.E.; Aufdenkampe, R.L.; Dailey, A.M.; Brown, C.M.; Scharf, A.R.; Earley, M.B.; Knippen, C.L.; et al. Post-learning stress enhances long-term memory and differentially influences memory in females depending on menstrual stage. Acta Psychol. 2015, 127–133. [Google Scholar] [CrossRef]
- Trammel, J.P.; Clore, G.L. Does stress enhance or impair memory consolidation? Cogn. Emot. 2014, 28, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Andreano, J.M.; Arjomandi, H.; Cahill, L. Menstrual cycle modulation of the relationship between cortisol and long-term memory. Psychoneuroendocrinology 2008, 33, 874–882. [Google Scholar] [CrossRef]
- Yonelinas, A.P.; Parks, C.M.; Koen, J.D.; Jorgenson, J.; Mendoza, S.P. The effects of post-enconding stress on recognition memory: Examining the impact of skydiving in young men and women. Stress 2011, 14, 136–144. [Google Scholar] [CrossRef] [Green Version]
- McCullough, A.M.; Yonelinas, A.P. Cold-pressor stress after learning enhances familiarity-based recognition memory in men. Neurobiol. Learn. Mem. 2013, 106, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Corbett, B.; Weinberg, L.; Duarte, A. The effect of mild acute stress during memory consolidation on emotional recognition memory. Neurobiol. Learn. Mem. 2017, 145, 34–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chichinadze, K.; Chichinadze, N. Stress-induced increase of testosterone: Contributions of social status and sympathetic reactivity. Physiol. Behav. 2008, 94, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, J.C.; Hegner, R.E.; Dufty, A.M.; Ball, G.F. The challenge hypothesis: “Theoretical implications” for patterns of testosterone secretion, mating systems, and breeding strategies. Am. Nat. 1990, 136, 829–846. [Google Scholar] [CrossRef]
- Archer, J. Testosterone and human aggression: An evaluation of the challenge hypothesis. Neurosci. Biobehav. Rev. 2006, 30, 319–345. [Google Scholar] [CrossRef] [PubMed]
- Salvador, A. Steroid hormones and some evolutionary-relevant social interactions. Motiv. Emot. 2012, 36, 74–83. [Google Scholar] [CrossRef]
- Kirschbaum, C.; Pirke, J.; Hellhammer, D.H. The “Trier Social Stress Test”: A tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology 1993, 28, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Salvador, A.; Costa, R. Coping with competition: Neuroendocrine responses and cognitive variables. Neurosci. Biobehav. Rev. 2009, 33, 160–170. [Google Scholar] [CrossRef]
- Lennartsson, A.K.; Kushnir, M.M.; Bergquist, J.; Billig, H.; Jonsdottir, I.H. Sex steroid levels temporarily increase in response to acute psychosocial stress in healthy men and women. Int. J. Psychophysiol. 2012, 84, 246–253. [Google Scholar] [CrossRef]
- Bedgood, D.; Boggiano, M.M.; Turan, B. Testosterone and social evaluative stress: The moderating role of basal cortisol. Psychoneuroendocrinology 2014, 47, 107–115. [Google Scholar] [CrossRef]
- Deuter, C.E.; Schächninger, H.; Best, D.; Neumann, R. Effects of two dominance manipulations on the stress response: Cognitive and embodied influences. Biol. Psychol. 2016, 119, 184–189. [Google Scholar] [CrossRef]
- Phan, J.M.; Schneider, E.; Peres, J.; Miocevic, O.; Meyer, V.; Shirtcliff, E.A. Social evaluative threat with verbal performance feedback alters neuroendocrine response to stress. Horm. Behav. 2017, 96, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Schoofs, D.; Wolf, O.T. Are salivary gonadal steroid concentrations influenced by acute psychosocial stress? A study using the Trier Social Stress Test (TSST). Int. J. Psychophysiol. 2011, 80, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Gerra, G.; Zimovic, A.; Zambelli, U.; Timpano, M.; Reali, N.; Bernasconi, S.; Brambilla, F. Neuroendocrine responses to psychological stress in adolescents with anxiety disorder. Neuropsychobiology. 2000, 42, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Heinz, A.; Hermann, D.; Smolka, M.N.; Rieks, M.; Gräf, K.J.; Pöhlau, D.; Kuhn, W.; Bauer, M. Effects of acute psychological stress on adhesion molecules, interleukins and sex hormones: Implications for coronary heart disease. Psychopharmacology 2003, 165, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Romero-Martínez, A.; González-Bono, E.; Salvador, A.; Moya-Albiol, L. Declarative verbal memory impairments in middle-aged women who are caregivers of offspring with autism spectrum disorders: The role of negative affect and testosterone. Memory 2016, 24, 640. [Google Scholar] [CrossRef] [PubMed]
- Smeets, T.; Cornelisse, S.; Quaedflieg, C.W.E.M.; Meyer, T.; Jelicic, M.; Merckelbach, H. Introducing the Maastricht Acute Stress Test (MAST): A quick and non-invasive approach to elicit robust autonomic and glucocorticoid stress responses. Psychoneuroendocrinology 2012, 37, 1998–2008. [Google Scholar] [CrossRef] [PubMed]
- Janowsky, J.S. Thinking with your gonads: Testosterone and cognition. Trends Cogn. Sci. 2006, 10, 77–82. [Google Scholar] [CrossRef]
- McEwen, B.S.; Milner, T.A. Understanding the broad influence of sex hormones and sex differences in the brain. J. Neurosci. Res. 2017, 95, 24–39. [Google Scholar] [CrossRef] [Green Version]
- Beyenburg, S.; Watzka, M.; Clusmann, H.; Blumcke, I.; Bidlingmaier, F.; Elger, C.E.; Stoffel-Wagner, B. Androgen receptor mRNA expression in the human hippocampus. Neurosci. Lett. 2000, 294, 25–28. [Google Scholar] [CrossRef]
- Cherrier, M.M.; Asthana, S.; Plymate, S.; Baker, L.; Matsumoto, A.M.; Peskind, E.; Raskind, M.A.; Brodkin, K.; Bremner, W.; Petrova, A.; et al. Testosterone supplementation improves spatial and verbal memory in healthy older men. Neurology 2001, 57, 80–88. [Google Scholar] [CrossRef]
- Wolf, O.T.; Kirschbaum, C. Endogenous estradiol and testosterone levels are associated with cognitive performance in older women and men. Horm. Behav. 2002, 41, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Pinquart, M.; Sorensen, S. Ethnic differences in stressors, resources, and psychological outcomes of family caregiving: A meta-analysis. Gerontologist 2005, 45, 90–106. [Google Scholar] [CrossRef]
- Shah, S.; Bell, R.J.; Savage, G.; Goldstat, R.; Papalia, M.A.; Kulkarni, J.; Donath, S.; Davis, S.R. Testosterone aromatization and cognition in women: A randomized, placebo-controlled trial. Menopause 2006, 13, 600–608. [Google Scholar] [CrossRef]
- Boss, L.; Kang, D.H.; Marcus, M.; Bergstrom, N. Endogenous sex hormones and cognitive function in older adults: A systematic review. West. J. Nurs. Res. 2014, 36, 388–426. [Google Scholar] [CrossRef] [PubMed]
- Thilers, P.; MacDonald, W.S.; Herlitz, A. The effect of endogenous free testosterone on cognitive performance: A population-based study in 35 to 90 year old men and women. Psychoneuroendocrinology 2006, 31, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Dettenborn, L.; Hinkelmann, K.; Muhtz, C.; Gao, W.; Wingenfeld, K.; Spitzer, C.; Mortiz, S.; Kirschabaum, C.; Otte, C. Hair testosterone and visuoespatial memory in middle-aged men and women with and without depressive symptoms. Psychoneuroendocrinology 2013, 38, 2373–2377. [Google Scholar] [CrossRef] [PubMed]
- Bove, R.M.; Gerweck, A.V.; Mancuso, S.M.; Bredella, M.A.; Sherman, J.C.; Miller, K.K. Association between adiposity and cognitive function in young men: Hormonal mechanisms. Obesity 2016, 24, 954–961. [Google Scholar] [CrossRef] [Green Version]
- Romero-Martínez, A.; Ruiz-Robledillo, N.; Moya-Albiol, L. Depressive mood and testosterone related to declarative verbal memory decline in middle-aged caregivers of children with eating disorders. Int. J. Environ. Res. Public Health 2016, 13, 286. [Google Scholar] [CrossRef] [Green Version]
- Lovallo, W. The cold pressor test and autonomic function: A review and integration. Psychophysiology 1975, 12, 268–282. [Google Scholar] [CrossRef]
- Bali, A.; Jaggi, A.S. Clinical experimental stress studies: Methods and assessment. Rev. Neurosci. 2015, 26, 555–579. [Google Scholar] [CrossRef]
- Pulopulos, M.M.; Hidalgo, V.; Puig-Perez, S.; Salvador, A. Psychophysiological response to social stressors: Relevance of sex and age. Psicothema 2018, 30, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Vila, J.; Sánchez, M.; Ramírez, I.; Fernández, M.C.; Cobos, P.; Rodríguez, S.; Muñoz, M.A.; Tormo, M.P.; Herrero, M.; Segarra, P.; et al. El sistema internacional de imagines afectivas (IAPS): Adaptación española, Segunda Parte. Rev. Psicol. Gral. Aplic. 2001, 54, 635–657. [Google Scholar] [CrossRef]
- Lang, P.J.; Bradley, M.M.; Cuthbert, B.N. International Affective Picture System (IAPS): Affective Ratings of Pictures and Instruction Manual; Technical Report A-6; University of Florida: Gainesville, FL, USA, 2005. [Google Scholar]
- Lang, P.J. Behavioral treatment and bio-behavioral assessment: Computer applications. In Technology in Mental Health Care Delivery Systems; Sidowski, J.B., Johnson, J.H., Williams, T.A., Eds.; Ablex: Norwood, NJ, USA, 1980; pp. 119–137. [Google Scholar]
- Hidalgo, V.; Pulopulos, M.M.; Puig-Perez, S.; Espin, L.; Gomez-Amor, J.; Salvador, A. Acute stress affects free recall and recognition of pictures differently depending on age and sex. Behav. Brain Res. 2015, 292, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Cornelisse, S.; van Stegeren, A.H.; Joëls, M. Implications of psychosocial stress on memory formation in a typical male versus female student sample. Psychoneuroendocrinology 2011, 36, 569–578. [Google Scholar] [CrossRef]
- Seisdedos, N. State-Trait Anxiety Inventory; TEA Ediciones: Madrid, Spain, 1988. [Google Scholar]
- Lai, J.C.L.; Lee, D.Y.H.; Leung, M.O.Y.; Lam, Y.W. Daily hassles, loneliness, and diurnal salivary cortisol in emerging adults. Horm. Behav. 2019, 115, 104558. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.C.L.; Leung, M.O.Y.; Lee, D.Y.H.; Lam, Y.W.; Berning, K. Loneliness and diurnal salivary cortisol in emerging adults. Int. J. Mol. Sci. 2018, 19, 1944. [Google Scholar] [CrossRef] [Green Version]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Salzmann, S.; Euteneuer, F.; Strahler, J.; Laferton, J.A.C.; Nater, U.M.; Rief, W. Optimizing expectations and distraction leads to lower cortisol levels after acute stress. Psychoneuroendocrinology 2018, 88, 144–152. [Google Scholar] [CrossRef]
- Romero-Martínez, A.; González-Bono, E.; Lila, M.; Moya-Albiol, L. Testosterone/cortisol ratio in response to acute stress: A possible marker of risk for marital violence. Soc. Neurosci. 2013, 8, 240–247. [Google Scholar] [CrossRef]
- Pannizon, M.S.; Hayger, R.L.; Xian, H.; Jacobson, K.; Lyons, M.J.; Franz, C.E.; Kremen, W.S. Interactive effects of testosterone and cortisol on hippocampal volumen and episodic memory in middle-aged men. Psychoneuroendocrinology 2018, 91, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Sazma, M.A.; McCullough, A.M.; Shields, G.S.; Yonelinas, A.P. Using acute stress to improve episodic memory: The critical role of contextual binding. Neurobiol. Learn. Mem. 2019, 158, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shields, G.S.; Dunn, T.M.; Trainor, B.C.; Yonelinas, A.P. Determing the biological associates of acute cold pressor post-encoding stress effects on human memory: The role of salivary interleukin-1 β. Brain Behav. Immun. 2019, 81, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Shields, G.S.; Sazma, M.A.; McCullough, A.M.; Yonelinas, A.P. The effects of acute stress on episodic memory: A meta-analysis and integrative review. Psychol. Bull. 2017, 143, 636–675. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response | Condition | Time | Condition × Time |
| Anxiety Self-Reported | F (1, 33) = 22.23, p < 0.001, η2p = 0.402 | F (1, 33) = 25.88, p < 0.001, η2p = 0.440 | F (1, 33) = 31.67, p < 0.001, η2p = 0.490 |
| Cortisol | |||
| Acquisition Session | F (1, 35) = 19.944, p < 0.001, η2p = 0.363 | F (1.745, 61.084) = 6.474, p = 0.004, η2p = 0.156 | F (1.745, 61.084) = 15.970, p < 0.001, η2p = 0.313 |
| Memory Testing Session | F (1, 35) = 2.163, p = 0.150, η2p = 0.058 | F (1, 35) = 14.170, p = 0.001, η2p = 0.288 | F (1, 35) = 0.001, p = 0.975, η2p = 0.001 |
| Testosterone | |||
| Acquisition Session | F (1, 35) = 0.275, p = 0.604, η2p = 0.008 | F (2.375, 83.131) = 3.747, p = 0.021, η2p = 0.097 | F (2.375, 83.131) = 2.101, p = 0.120, η2p = 0.057 |
| Memory Testing Session | F (1, 35) = 0.135, p = 0.715, η2p = 0.004 | F (1, 35) = 0.038, p = 0.847, η2p = 0.001 | F (1, 35) = 0.214, p = 0.646, η2p = 0.006 |
| Memory | Condition | Category | Condition × Category |
| Ratings Pictures | |||
| Valence | F (1, 35) = 2.366, p = 0.133, η2p = 0.063 | F (2.067, 72.353) = 206.301, p < 0.001, η2p = 0.855 | F (2.067, 72.353) = 0.969, p = 0.387, η2p = 0.027 |
| Arousal | F (1, 35) = 0.331, p = 0.560, η2p = 0.009 | F (2.787, 97.532) = 61.150, p < 0.001, η2p = 0.636 | F (2.787, 97.532) = 0.413, p = 0.729, η2p = 0.012 |
| Memory Performance | |||
| Free Recall | F (1, 35) = 0.000, p = 0.992, η2p = 0.000 | F (4, 140) = 23.400, p < 0.001, η2p = 0.401 | F (4, 140) = 1.042, p = 0.388, η2p = 0.029 |
| Recognition | F (1, 34) = 0.485, p = 0.491, η2p = 0.014 | F (2.758, 83.764) = 8.045, p < 0.001, η2p = 0.191 | F (2.758, 83.764) = 0.493, p = 0.672, η2p = 0.014 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hidalgo, V.; Villada, C.; Salvador, A. Post-Encoding Stress Does Not Enhance Memory Consolidation: The Role of Cortisol and Testosterone Reactivity. Brain Sci. 2020, 10, 995. https://doi.org/10.3390/brainsci10120995
Hidalgo V, Villada C, Salvador A. Post-Encoding Stress Does Not Enhance Memory Consolidation: The Role of Cortisol and Testosterone Reactivity. Brain Sciences. 2020; 10(12):995. https://doi.org/10.3390/brainsci10120995
Chicago/Turabian StyleHidalgo, Vanesa, Carolina Villada, and Alicia Salvador. 2020. "Post-Encoding Stress Does Not Enhance Memory Consolidation: The Role of Cortisol and Testosterone Reactivity" Brain Sciences 10, no. 12: 995. https://doi.org/10.3390/brainsci10120995