Modular Diversity of the BLUF Proteins and Their Potential for the Development of Diverse Optogenetic Tools


Abstract
:1. Introduction
2. Materials and Methods
2.1. Database of Sequences used in this Analysis
2.2. Phylogenetic Analysis
2.3. Analysis and Homology Modeling of the BLUF Domain
3. Results and Discussion
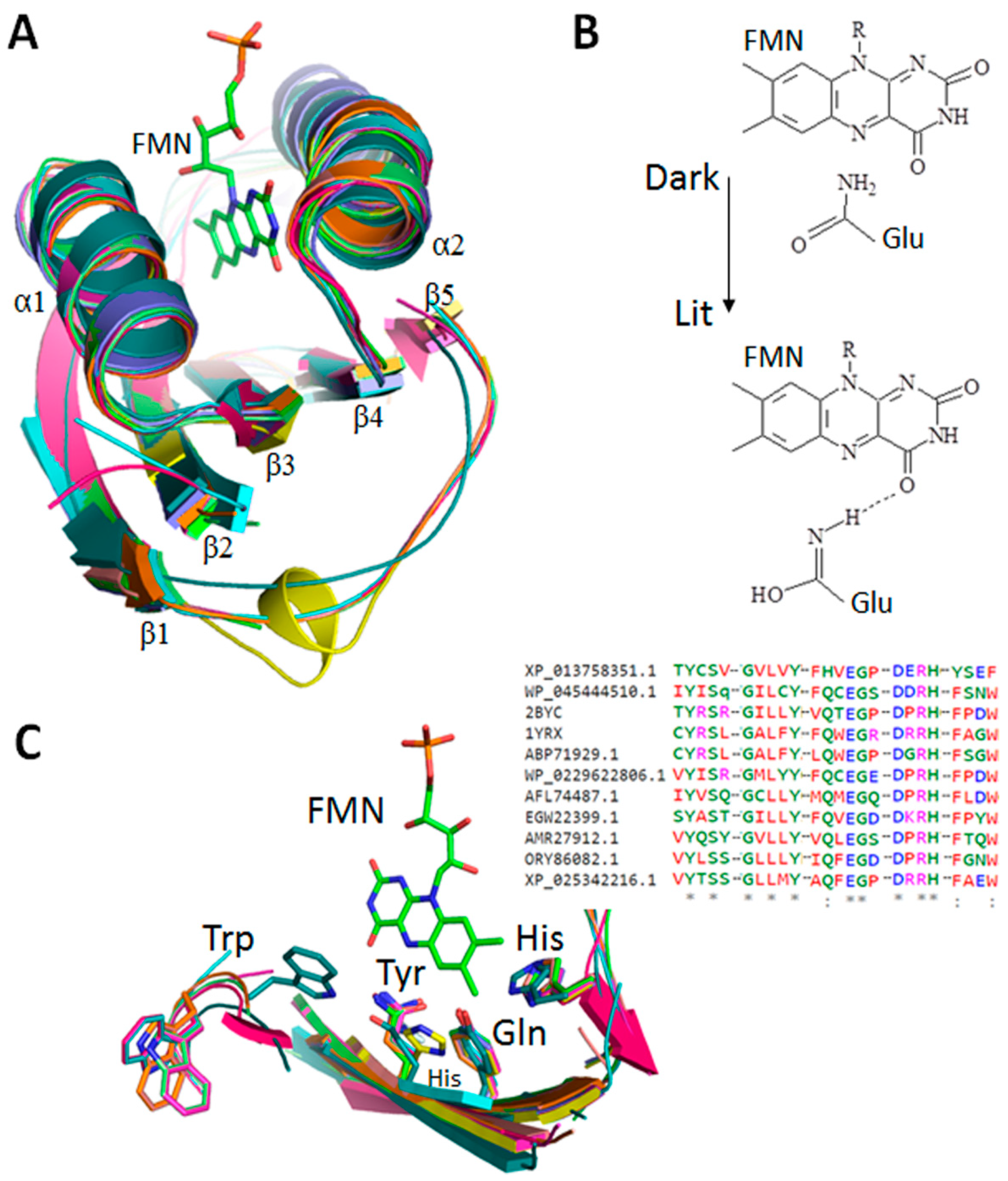
3.1. BLUF Sequences, Modular Domains and Phylogenetic Analysis
3.2. Modular Diversity of BLUF Domains
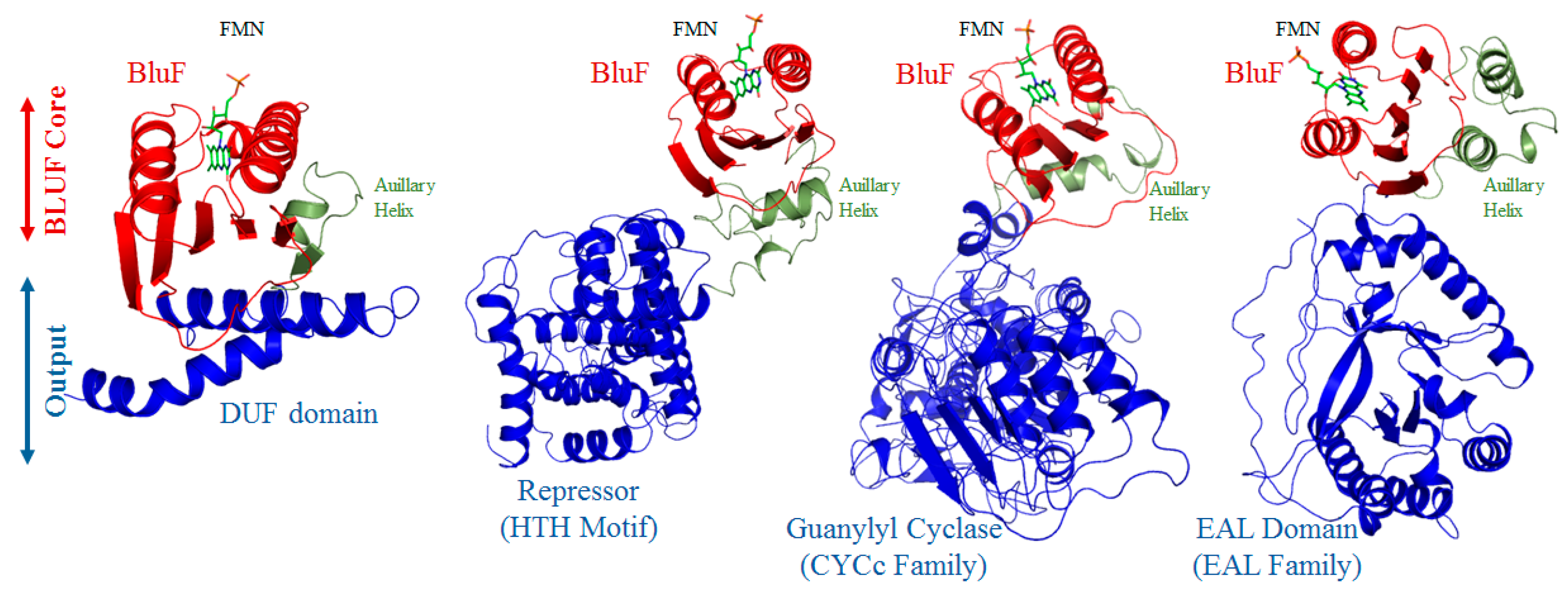
3.3. BLUF Modules in Association with the Effector Domains
3.3.1. EAL and GGDEF Domain
3.3.2. PsiE Domain
3.3.3. Cyclase Homology Domain (CHD)
3.3.4. PAS Domain
3.3.5. B12 Binding Domain
3.3.6. PRK Superfamily
3.3.7. DNA pol 3 gamma3 Superfamily
3.3.8. Cytochrome p450/ p450 Superfamily
3.3.9. REC Domain
3.3.10. TetR and AcrR Domain
3.3.11. Endonuclease 3c and Endonuclease-NS Domain
3.3.12. AraC Domain
3.3.13. Abhydrolase (α/β Hydrolase) Superfamily
3.3.14. Domain of Unknown Function (DUF)
3.3.15. ANK Repeats
3.3.16. RhoGEF Domain
3.3.17. PDZ Domain
3.3.18. GAL4-Fungal TF MHR Domain
3.4. BLUF Proteins for Optogenetic Tools
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Purcell, E.B.; Crosson, S. Photoregulation in prokaryotes. Curr. Opin. Microbiol. 2008, 11, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Ohlendorf, R.; Vidavski, R.R.; Eldar, A.; Moffat, K.; Moglich, A. From dusk till dawn: One-plasmid systems for light-regulated gene expression. J. Mol. Biol. 2012, 416, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Stierl, M.; Stumpf, P.; Udwari, D.; Gueta, R.; Hagedorn, R.; Losi, A.; Gärtner, W.; Petereit, L.; Efetova, M.; Schwarzel, M.; et al. Light modulation of cellular cAMP by a small bacterial photoactivated adenylyl cyclase, bPAC, of the soil bacterium Beggiatoa. J. Biol. Chem. 2011, 286, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Barends, T.R.M.; Hartmann, E.; Griese, J.J.; Beitlich, T.; Kirienko, N.V.; Ryjenkov, D.A.; Reinstein, J.; Shoeman, R.L.; Gomelsky, R.; Schlichting, I. Structure and mechanism of a bacterial light-regulated cyclic nucleotide phosphodiesterase. Nature 2009, 459, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Masuda, S.; Bauer, C.E. AppA is a blue light photoreceptor that antirepresses photosynthesis gene expression in Rhodobacter sphaeroides. Cell 2002, 110, 613–623. [Google Scholar] [CrossRef]
- Tanwar, M.; Nahar, S.; Gulati, S.; Veetil, S.K.; Kateriya, S. Molecular determinant modulates thermal recovery kinetics and structural integrity of the bacterial BLUF photoreceptor. FEBS Lett. 2016, 590, 2146–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Nakasone, Y.; Okajima, K.; Ikeuchi, M.; Tokutomi, S.; Terazima, M. Oligomeric-state dependent conformational change of the BLUF protein TePixD (Tll0078). J. Mol. Biol. 2009, 386, 1290–1300. [Google Scholar] [CrossRef]
- Tanaka, K.; Nakasone, Y.; Okajima, K.; Ikeuchi, M.; Tokutomi, S.; Terazima, M. Light-induced conformational change and transient dissociation reaction of the BLUF photoreceptor Synechocystis PixD (Slr1694). J. Mol. Biol. 2011, 409, 773–785. [Google Scholar] [CrossRef]
- Yuan, H.; Dragnea, V.; Wu, Q.; Gardner, K.H.; Bauer, C.E. Mutational and structural studies of the PixD BLUF output signal that affects light-regulated interactions with PixE. Biochemistry 2011, 50, 6365–6375. [Google Scholar] [CrossRef]
- Laan, W.; Bednarz, T.; Heberle, J.; Hellingwerf, K.J. Chromophore composition of a heterologously expressed BLUF-domain. Photochem. Photobiol. Sci. 2004, 3, 1011–1016. [Google Scholar] [CrossRef]
- Kennis, J.T.M.; Mathes, T. Molecular eyes: Proteins that transform light into biological information. Interface Focus 2013, 3, 20130005. [Google Scholar] [CrossRef] [PubMed]
- Laan, W.; van der Horst, M.A.; van Stokkum, I.H.; Hellingwerf, K.J. Initial characterization of the primary photochemistry of AppA, a blue-light-using flavin adenine dinucleotide-domain containing transcriptional antirepressor protein from Rhodobacter sphaeroides: A key role for reversible intramolecular proton transfer from the flavin adenine dinucleotide chromophore to a conserved tyrosine? Photochem. Photobiol. 2003, 78, 290–297. [Google Scholar] [PubMed]
- Kraft, B.J.; Masuda, S.; Kikuchi, J.; Dragnea, V.; Tollin, G.; Zaleski, J.M.; Bauer, C.E. Spectroscopic and mutational analysis of the blue-light photoreceptor AppA: A novel photocycle involving flavin stacking with an aromatic amino acid? Biochemistry 2003, 42, 6726–6734. [Google Scholar] [CrossRef] [PubMed]
- Unno, M.; Masuda, S.; Ono, T.-A.; Yamauchi, S. Orientation of a key glutamine residue in the BLUF domain from AppA revealed by mutagenesis, spectroscopy, and quantum chemical calculations. J. Am. Chem. Soc. 2006, 128, 5638–5639. [Google Scholar] [CrossRef]
- Penzkofer, A.; Tanwar, M.; Veetil, S.K.; Kateriya, S.; Stierl, M.; Hegemann, P. Photo-dynamics and thermal behavior of the BLUF domain containing adenylate cyclase NgPAC2 from the amoeboflagellate Naegleria gruberi NEG-M strain. Chem. Phys. 2013, 412, 96–108. [Google Scholar] [CrossRef]
- Bonetti, C.; Mathes, T.; van Stokkum, I.H.M.; Mullen, K.M.; Groot, M.-L.; van Grondelle, R.; Hegemann, P.; Kennis, J.T.M. Hydrogen-bond switching among flavin and amino acid side chains in the BLUF photoreceptor observed by ultrafast infrared spectroscopy. Biophys. J. 2008, 95, 4790–4802. [Google Scholar] [CrossRef] [PubMed]
- Gauden, M.; van Stokkum, I.H.M.; Key, J.M.; Luhrs, D.C.; van Grondelle, R.; Hegemann, P.; Kennis, J.T.M. Hydrogen-bond switching through a radical pair mechanism in a flavin-binding photoreceptor. Proc. Natl. Acad. Sci. USA 2006, 103, 10895–10900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Braatsch, S.; Osterloh, L.; Klug, G. A eukaryotic BLUF domain mediates light-dependent gene expression in the purple bacterium Rhodobacter sphaeroides 2. 4. 1. Proc. Natl. Acad. Sci. USA 2004, 101, 12306–12311. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.J.; Yang, Y. Spatiotemporal control of gene expression by a light-switchable transgene system. Nat. Methods 2012, 9, 266–269. [Google Scholar] [CrossRef]
- Strickland, D.; Moffat, K.; Sosnick, T.R. Light activated DNA binding in a designed allosteric protein. Proc. Natl. Acad. Sci. USA 2008, 105, 10709–10714. [Google Scholar] [CrossRef]
- Moglich, A.; Moffat, K. Engineered photoreceptors as novel optogenetic tools. Photochem. Photobiol. Sci. 2010, 9, 1286–1300. [Google Scholar] [CrossRef] [PubMed]
- Moglich, A.; Ayers, R.A.; Moffat, K. Design and signaling mechanism of light-regulated histidine kinases. J. Mol. Biol. 2009, 385, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.I.; Frey, D.; Lungu, O.I.; Jaehrig, A.; Schlichting, I.; Kuhlman, B.; Hahn, K.M. A genetically encoded photoactivatable Rac controls the motility of living cells. Nature 2009, 461, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polstein, L.R.; Gersbach, C.A. Light-inducible spatiotemporal gene regulation using engineered transcription factors. Mol. Ther. 2012, 20, 193–194. [Google Scholar]
- Krauss, U.; Drepper, T.; Jaeger, K.E. Enlightened enzymes: Strategies to create novel photoresponsive proteins. Chem. Eur. J. 2011, 17, 2552–2560. [Google Scholar] [CrossRef] [PubMed]
- Hisert, K.B.; MacCoss, M.; Shiloh, M.U.; Darwin, K.H.; Singh, S.; Jones, R.A.; Ehrt, S.; Zhang, Z.; Gaffney, B.L.; Gandotra, S.; et al. A glutamate-alanine-leucine (EAL) domain protein of Salmonella controls bacterial survival in mice, antioxidant defence and killing of macrophages: Role of cyclic diGMP. Mol. Microbiol. 2005, 56, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.R.; D’argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside antibiotics induce bacterial biofilm formation. Nature 2005, 436, 1171–1175. [Google Scholar] [CrossRef]
- Kulesekara, H.; Lee, V.; Brencic, A.; Liberati, N.; Urbach, J.; Miyata, S.; Lee, D.G.; Neely, A.N.; Hyodo, M.; Hayakawa, Y.; et al. Analysis of Pseudomonas aeruginosa diguanylate cyclases and phosphodiesterases reveals a role for bis (39-59)-cyclic-GMP in virulence. Proc. Natl. Acad. Sci. USA 2006, 103, 2839–2844. [Google Scholar] [CrossRef]
- Claret, L.; Miquel, S.; Vieille, N.; Ryjenkov, D.A.; Gomelsky, M.; Darfeuille-Michaud, A. The flagellar sigma factor FliA regulates adhesion and invasion of Crohn disease-associated Escherichia coli via a cyclic dimeric GMP-dependent pathway. J. Biol. Chem. 2007, 282, 33275–33283. [Google Scholar] [CrossRef]
- Tschowri, N.; Busse, S.; Hengge, R. The BLUF-EAL protein YcgF acts as a direct anti-repressor in a blue-light response of Escherichia coli. Gene Dev. 2009, 23, 522–534. [Google Scholar] [CrossRef]
- Kader, A.; Simm, R.; Gerstel, U.; Morr, M.; Römling, U. Hierarchical involvement of various GGDEF domain proteins in rdar morphotype development of Salmonella enteric serovar Typhimurium. Mol. Microbiol. 2006, 60, 602–616. [Google Scholar] [CrossRef]
- Wei, C.; Jiang, W.; Zhao, M.; Ling, J.; Zeng, X.; Deng, J.; Jin, D.; Dow, J.M.; Sun, W. A systematic analysis of the role of GGDEF-EAL domain proteins in virulence and motility in Xanthomonas oryzae pv. Oryzicola. Sci. Rep. 2016, 6, 23769. [Google Scholar] [CrossRef]
- Lindner, R.; Hartmann, E.; Tarnawski, M.; Winkler, A.; Frey, D.; Reinstein, J.; Meinhart, A.; Schlichting, I. Photoactivation Mechanism of a Bacterial Light-Regulated Adenylyl Cyclase. J. Mol. Biol. 2017, 429, 1336–1351. [Google Scholar] [CrossRef] [PubMed]
- Penzkofer, A.; Tanwar, M.; Veetil, S.K.; Kateriya, S. Photo-dynamics of photoactivated cyclase LiPAC from the spirochetebacterium Leptonema illini strain 3055T. Trends Appl. Spectrol. 2014, 11, 40–62. [Google Scholar]
- Iseki, M.; Matsunaga, S.; Murakami, A.; Ohno, K.; Shiga, K.; Yoshida, K.; Sugai, M.; Takahashi, T.; Hori, T.; Watanabe, M. A blue-light-activated adenylyl cyclase mediates photoavoidance in Euglena gracilis. Nature 2002, 415, 1047–1051. [Google Scholar] [CrossRef] [PubMed]
- Fritz-Laylin, L.K.; Prochnik, S.E.; Ginger, M.L.; Dacks, J.B.; Carpenter, M.L.; Field, M.C.; Kuo, A.; Paredez, A.; Chapman, J.; Pham, J.; et al. The genome of Naegleria gruberi illuminates early eukaryotic versatility. Cell 2010, 140, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.-H.; Moskvin, O.V.; Siltberg-Liberles, J.; Gomelsky, M. Natural and engineered photoactivated nucleotidylcyclases for optogenetic applications. J. Biol. Chem. 2010, 285, 41501–41508. [Google Scholar] [CrossRef] [PubMed]
- Ohki, M.; Sugiyama, K.; Kawai, F.; Tanaka, H.; Nihei, Y.; Unzai, S.; Takebe, M.; Matsunaga, S.; Adachi, S.I.; Shibayama, N.; et al. Structural insight into photoactivation of an Adenylate cyclase from a photosynthetic cyanobacterium. Proc. Natl. Acad. Sci. USA 2016, 113, 6659–6664. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.L.; Zhulin, I.B. PAS domains: Internal sensors of oxygen, redox potential, and light. Microbiol. Mol. Biol. Rev. 1999, 63, 479–506. [Google Scholar] [PubMed]
- Baca, M.; Borgstahl, G.E.; Boissinot, M.; Burke, P.M.; Williams, D.R.; Slater, K.A.; Getzoff, E.D. Complete chemical structure of photoactive yellow protein: Novel thioester-linked 4-hydroxycinnamyl chromophore and photocycle chemistry. Biochemistry 1994, 33, 14369–14377. [Google Scholar] [CrossRef] [PubMed]
- Giraud, E.; Fardoux, J.; Fourrier, N.; Hannibal, L.; Genty, B.; Bouyer, P.; Dreyfus, B.; Verméglio, A. Bacteriophytochrome controls photosystem synthesis in anoxygenic bacteria. Nature 2002, 417, 202. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, B.L. Sensing the light: Photoreceptive systems and signal transduction in cyanobacteria. Mol. Microbiol. 2007, 64, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Penzkofer, A.; Kateriya, S.; Hegemann, P. Photodynamics of the optogenetic BLUF coupled photoactivated adenylyl cyclases (PACs). Dyes Pigments 2016, 135, 102–112. [Google Scholar] [CrossRef]
- Christie, J.M.; Gawthorne, J.; Young, G.; Fraser, N.J.; Roe, A.J. LOV to BLUF: Flavoprotein Contributions to the Optogenetic Toolkit. Mol. Plant 2012, 5, 533–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losi, A.; Gardner, K.H.; Moglich, A. Blue-Light Receptors for optogentetics. Chem. Rev. 2018, 118, 10659–10709. [Google Scholar] [CrossRef] [PubMed]
- Geer, L.Y.; Domrachev, M.; Lipman, D.J.; Bryant, S.H. CDART: Protein homology by domain architecture. Genome Res. 2002, 12, 1619–1623. [Google Scholar] [CrossRef]
- Hall, A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for window 95/98/NT. Nucliec Acid Res. 1999, 41, 95–98. [Google Scholar]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Nobel, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acid Res. 2009, 37, 202–208. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2016, 45, 362–368. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comp. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6. 0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Hooft, R.W.; Vriend, G.; Sander, C.; Abola, E.E. Errors I protein structures. Nature 1996, 381, 272. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Gauden, M.; Yeremenko, S.; Laan, W.; van Stokkum, I.H.M.; Ihalainen, J.A.; van Grondelle, R.; Hellingwerf, K.J.; Kennis, J.T.M. Photocycle of the flavin-binding photoreceptor appa, a bacterial transcriptional antirepressor of photosynthesis genes. Biochemistry 2005, 44, 3653–3662. [Google Scholar] [CrossRef]
- Fukushima, Y.; Okajima, K.; Shibata, Y.; Ikeuchi, M.; Itoh, S. Primary intermediate in the photocycle of a blue-light sensory BLUF FAD-protein, Tll0078, of Synechococcus elongatus BP-1. Biochemistry 2005, 44, 5149–5158. [Google Scholar] [CrossRef]
- Moglich, A.; Ayers, R.A.; Moffat, K. Addition at the molecular level: Signal integration in designed Per-ARNT-Sim receptor proteins. J. Mol. Biol. 2010, 400, 477–486. [Google Scholar] [CrossRef]
- Moglich, A.; Yang, X.; Ayers, R.A.; Moffat, K. Structure and function of plant photoreceptors. Annu. Rev. Plant Biol. 2010, 61, 21–47. [Google Scholar] [CrossRef]
- Christie, J.M.; Swartz, T.E.; Bogomolni, R.A.; Briggs, W.R. Phototropin LOV domains exhibit distinct roles in regulating photoreceptor function. Plant J. 2002, 32, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Christie, J.M. Phototropin blue-light receptors. Ann. Rev. Plant Biol. 2007, 58, 21–45. [Google Scholar] [CrossRef] [PubMed]
- Delano, W.L. PyMOL Molecular Viewer; Delano Scientific: Palo Alto, CA, USA, 2008. [Google Scholar]
- Galperin, M.Y.; Nikolskaya, A.N.; Koonin, E.V. Novel domains of the prokaryotic two-component signal transduction systems. FEMS Microbiol. Lett. 2001, 203, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.; Pultz, I.S.; Kulasekara, H.D.; Miller, S.I. The bacterial second messenger c-di-GMP: Mechanisms of signaling. Cell. Microbiol. 2011, 13, 1122–1129. [Google Scholar] [CrossRef] [PubMed]
- Sondermann, H.; Shikuma, N.J.; Yildiz, F.H. You’ve come a long way: C-di-GMP signaling. Curr. Opin. Microbiol. 2012, 13, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Malone, J.; Williams, R.; Christen, M.; Jenal, U.; Spiers, A.J.; Rainey, P.B. The structure–function relationship of WspR, a Pseudomonas fluorescens response regulator with a GGDEF output domain. Microbiology 2007, 153, 980–994. [Google Scholar] [CrossRef]
- Wassmann, P.; Chan, C.; Paul, R.; Beck, A.; Heerklotz, H.; Jenal, U.; Schirmer, T. Structure of BeF3-modified response regulator PleD: Implications of diguanylate cyclase activation, catalysis, and feedback inhibition. Structure 2007, 15, 915–927. [Google Scholar] [CrossRef] [PubMed]
- Rao, F.; Yang, Y.; Qi, Y.; Liang, Z.X. Catalytic mechanism of c-di-GMP specific phosphodiesterase: A study of the EAL domain-containing RocR from Pseudomonas aeruginosa. J. Bacteriol. 2008, 190, 3622–3631. [Google Scholar] [CrossRef]
- Qi, Y.; Rao, F.; Luo, Z.; Liang, Z.X. A flavin cofactor-binding PAS domain regulates c-di-GMP synthesis in AxDGC2 from Acetobacter xylinum. Biochemistry 2009, 48, 10275–10285. [Google Scholar] [CrossRef]
- Christen, M.; Christen, B.; Folcher, M.; Schauerte, A.; Jenal, U. Identification and characterization of a cyclic di-GMP-specific phosphodiesterase and its allosteric control by GTP. J. Biol. Chem. 2005, 285, 30829–30837. [Google Scholar] [CrossRef]
- Newell, P.D.; Yoshioka, S.; Hvorecny, K.L.; Monds, R.D.; O’Toole, G.A. Systematic analysis of diguanylate cyclases that promote biofilm formation by Pseudomonas fluorescens Pf0-1. J. Bacteriol. 2011, 193, 4685–4698. [Google Scholar] [CrossRef] [PubMed]
- Newell, P.D.; Monds, R.D.; O’Toole, G.A. LapD is a bis-(3′,5′)-cyclic dimeric GMP-binding protein that regulates surface attachment by Pseudomonas fluorescens Pf0-1. Proc. Natl. Acad. Sci. USA 2009, 106, 3461–3466. [Google Scholar] [CrossRef] [PubMed]
- Hengge, R.; Galperin, M.Y.; Ghigo, J.M.; Gomelsky, M.; Green, J.; Hughes, K.T.; Jenal, U.; Landini, P. Systematic nomenclature for GGDEF and EAL domain-containing cyclic di-GMP turnover proteins of Escherichia coli. J. Bacteriol. 2016, 198, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Tian, T.; Li, X.; Fan, S.; Chen, H.; Wu, M.; Yang, C.-H.; He, C. The degenerate EAL GGDEF Domain protein filp functions as a cyclic di-gmp receptor and specifically interacts with the PilZ-domain protein PXO_02715 to regulate virulence in Xanthomonas oryzae pv. oryzae. MPMI 2014, 27, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Babitzke, P.; Kushner, S.R.; Romeo, T. Identification of a novel regulatory protein (CsrD) that targets the global regulatory RNAs CsrB and CsrC for degradation by RNase E. Genes Dev. 2006, 20, 2605–2617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Gardner, K.H. Structure and Insight into Blue Light-Induced Changes in the BlrP1 BLUF Domain. Biochemistry 2009, 48, 2620–2629. [Google Scholar] [CrossRef]
- Yuan, H.; Anderson, S.; Masuda, S.; Dragnea, V.; Moffat, K.; Bauer, C. Crystal structures of the Synechocystis photoreceptor Slr1694 reveal distinct structural states related to signaling. Biochemistry 2006, 45, 12687–12694. [Google Scholar] [CrossRef]
- Jung, A.; Domratcheva, T.; Tarutina, M.; Wu, Q.; Ko, W.H.; Shoeman, R.L.; Gomelsky, M.; Gardner, K.H.; Schlichting, I. Structure of a bacterial BLUF photoreceptor: Insights into blue light-mediated signal transduction. Proc. Natl. Acad. Sci. USA 2005, 102, 12350–12355. [Google Scholar] [CrossRef] [Green Version]
- Kita, A.; Okajima, K.; Morimoto, Y.; Ikeuchi, M.; Miki, K. Structure of a cyanobacterial BLUF protein, Tll0078, containing a novel FAD-binding blue light sensor domain. J. Mol. Biol. 2005, 349, 1–9. [Google Scholar] [CrossRef]
- Wood, T.K. Insights on Escherichia coli biofilm formation and inhibition from whole-transcriptome profiling. Environ. Microbiol. 2009, 11, 1–15. [Google Scholar] [CrossRef]
- Zucca, S.; Pasotti, L.; Politi, N.; Casanova, M.; Mazzini, G.; Cusella De Angelis, M.G.; Magni, P. Multi-faceted characterization of a novel LuxR repressible promoter library for Escherichia coli. PLoS ONE 2015, 10, e0126264. [Google Scholar] [CrossRef] [PubMed]
- Kendall, M.M.; Sperandio, V. Cell-to-cell signaling in E. coli and Salmonella. Eco Sal Plus 2014, 6, 1. [Google Scholar]
- Kim, S.K.; Kimura, S.; Shinagawa, H.; Nakata, A.; Lee, K.S.; Wanner, B.L.; Makino, K. Dual transcriptional regulation of the Escherichia coli phosphate-starvation-inducible psiE gene of the phosphate regulon by phob and the cyclic AMP (cAMP)-cAMP receptor protein complex. J. Bacteriol. 2000, 182, 5596–5599. [Google Scholar] [CrossRef] [PubMed]
- Linder, J.U.; Schultz, J.E. The class III adenylyl cyclases: Multi-purpose signaling modules. Cell. Signal. 2003, 15, 1081–1089. [Google Scholar] [CrossRef]
- Sinha, S.C.; Sprang, S.R. Structures, mechanism, regulation and evolution of class III nucleotidylcyclases. In Reviews of Physiology Biochemistry and Pharmacology; Springer: Berlin/Heidelberg, Germany, 2006; pp. 105–140. [Google Scholar]
- Vercellino, I.; Rezabkova, L.; Olieric, V.; Polyhach, Y.; Weinert, T.; Kammerer, R.A.; Jeschke, G.; Korkhova, V.M. Role of the nucleotidyl cyclase helical domain in catalytically active dimer formation. Proc. Natl. Acad. Sci. USA 2017, 114, 9821–9828. [Google Scholar] [CrossRef]
- McIntosh, B.E.; Hogenesch, J.B.; Bradfield, C.A. Mammalian Per-Arnt-Sim proteins in environmental adaptation. Ann. Rev. Physiol. 2010, 72, 625–645. [Google Scholar] [CrossRef] [PubMed]
- Henry, J.T.; Crosson, S. Ligand-binding PAS domains in a genomic, cellular, and structural context. Ann. Rev. Microbiol. 2011, 65, 261–286. [Google Scholar] [CrossRef]
- Oka, Y.; Matsushita, T.; Mochizuki, N.; Quail, P.H.; Nagatani, A. Mutant screen distinguishes between residues necessary for light-signal perception and signal transfer by phytochrome B. PLoS Genet. 2008, 4, e1000158. [Google Scholar] [CrossRef]
- Pellequer, J.L.; Wager-Smith, K.A.; Kay, S.A.; Getzoff, E.D. Photoactive yellow protein: A structural prototype for the three-dimensional fold of the PAS domain superfamily. Proc. Natl. Acad. Sci. USA 1998, 95, 5884–5890. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Yamamoto, H.; Bauer, C.E. Cobalamin’s (Vitamin B12) Surprising Function as a Photoreceptor. Trends Biochemi. Sci. 2016, 41, 647–650. [Google Scholar] [CrossRef]
- Drennan, C.L.; Huang, S.; Drummond, J.T.; Matthews, R.G.; Lidwig, M.L. How a protein binds B12: A 3.0 A X-ray structure of B12-binding domains of methionine synthase. Science 1994, 266, 1669–1674. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.L.; Matthews, R.G. Structure-based perspectives on B12-dependent enzymes. Annu. Rev. Biochem. 1997, 66, 269–313. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, S.; Jost, M.; Drennan, C.L.; Elías-Arnanz, M. A new facet of vitamin B12: Gene regulation by cobalamin-based photoreceptors. Ann. Rev. Biochem. 2017, 86, 485–514. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Guerrero, J.M.; Polanco, M.C.; Murillo, F.J.; Padmanabhan, S.; Elias-Arnanz, M. Light-dependent gene regulation by a coenzyme B12-based photoreceptor. Proc. Natl. Acad. Sci. USA 2011, 108, 7565–7570. [Google Scholar] [CrossRef] [PubMed]
- Jost, M.; Simpson, J.H.; Drennan, C.L. The transcription factor CarH safeguards use of adenosylcobalamin as a light sensor by altering the photolysis products. Biochemistry 2015, 54, 3231–3234. [Google Scholar] [CrossRef] [PubMed]
- Kutta, R.J.; Hardman, S.J.; Johannissen, L.O.; Bellina, B.; Messiha, H.L.; Ortiz-Guerrero, J.M.; Elias-Arnanz, M.; Padmanabhan, S.; Barran, P.; Scrutton, N.S.; et al. The photochemical mechanism of a B12-dependent photoreceptor protein. Nat. Commun. 2015, 6, 7907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jost, M.; Fernandez-Zapata, J.; Polanco, M.C.; Ortiz-Guerrero, J.M.; Chen, P.Y.; Kang, G.; Padmanabhan, S.; Elias-Arnanz, M.; Drennan, C.L. Structural basis for gene regulation by a B12-dependent photoreceptor. Nature 2015, 526, 536–541. [Google Scholar] [CrossRef] [Green Version]
- Roth, J.R.; Lawrence, J.G.; Bobik, T.A. Cobalamin (coenzymeB12): Synthesis and biological significance. Annu. Rev. Microbiol. 1996, 50, 137–181. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, P.; Guroff, G. Enzymatic formation of phenylpyruvic acid in Pseudomonas sp. (ATCC 11299A) and its regulation. J. Biol. Chem. 1965, 240, 3034–3038. [Google Scholar] [PubMed]
- Cotton, R.G.; Gibson, F. The biosynthesis of phenylalanine and tyrosine; enzymes converting chorismic acid into prephenic acid and their relationships to prephenate dehydratase and prephenate dehydrogenase. Biochim. Biophys. Acta 1965, 100, 76–88. [Google Scholar] [CrossRef]
- Schmidt, J.C.; Zalkin, H. Chorismate mutase-prephenate dehydratase. Partial purification and properties of the enzyme from Salmonella typhimurium. Biochemistry 1969, 8, 174–181. [Google Scholar]
- Kaushik, M.S.; Srivastava, M.; Mishra, A.K. Iron homeostasis in cyanobacteria. In Cyanobacteria: From Basic Science toApplications, 1st ed.; Mishra, A.K., Tiwari, D.N., Rai, A.N., Eds.; Academic Press: Cambridge, MA, USA, 2018; Volume 1, pp. 173–191. [Google Scholar]
- Harrison, D.H.; Runquist, J.A.; Holub, A.; Miziorko, H.M. The crystal structure of phosphoribulokinase from Rhodobacter sphaeroides reveals a fold similar to that of adenylate kinase. Biochemistry 1998, 37, 5074–5085. [Google Scholar] [CrossRef] [PubMed]
- Neuhard, J.; Tarpo, L. Location of the udk gene on the physical map of Escherichia coli. J. Bacteriol. 1993, 175, 5742–5743. [Google Scholar] [CrossRef] [PubMed]
- Song, W.J.; Jackowski, S. Cloning, sequencing, and expression of the pantothenate kinase (coaA) gene of Escherichia coli. J. Bacteriol. 1992, 174, 6411–6417. [Google Scholar] [CrossRef] [PubMed]
- Jergic, S.; Ozawa, K.; Williams, N.K.; Su, X.-C.; Scott, D.D.; Hamdan, S.M.; Crowther, J.A.; Otting, G.; Dixon, N.E. The unstructured C-terminus of the q subunit of Escherichia coli DNA polymerase III holoenzyme is the site of interaction with the a subunit. Nucleic Acid Res. 2007, 35, 2813–2824. [Google Scholar] [CrossRef] [PubMed]
- Snyder, A.K.; Williams, C.R.; Johnson, A.; O’Donnell, M.; Bloom, L.B. Mechanism of loading the Escherichia coli DNA polymerase III sliding clamp. II. Uncoupling the b and DNA binding activities of the g complex. J. Biol. Chem. 2004, 279, 4386–4393. [Google Scholar] [CrossRef] [PubMed]
- Jeruzalmi, D.; O’Donnell, M.; Kuriyan, J. Crystal Structure of the Processivity Clamp Loader Gamma (γ) Complex of E. coli DNA Polymerase III. Cell 2001, 106, 429–441. [Google Scholar] [CrossRef]
- Tsuchihashi, Z.; Kornberg, A. Translational frameshifting generates the g subunit of DNA polymerase III holoenzyme. Proc. Natl. Acad. Sci. USA 1990, 87, 2516–2520. [Google Scholar] [CrossRef]
- Cullman, G.; Fien, K.; Kobayashi, R.; Stillman, B. Characterization of the five replication factor C genes of Saccharomyces cerevisiae. Mol. Cell. Biol. 1995, 15, 4661–4671. [Google Scholar] [CrossRef]
- O’Donnell, M.; Onrust, R.; Dean, F.B.; Chen, M.; Hurwitz, J. Homology in accessory proteins of replicative polymerases—E. coli to humans. Nucleic Acids Res. 1993, 21, 1–3. [Google Scholar] [CrossRef]
- Munro, A.W.; McLean, K.J.; Grant, J.L.; Makris, T.M. Structure and function of the cytochrome P450 peroxygenase enzymes. Biochem. Soc. Trans. 2018, 46, 183–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Chang, Z.; Pan, Z.; Fu, Z.-Q.; Wang, X. Modes of heme binding and substrate access for cytochrome P450 CYP74A revealed by crystal structures of allene oxide synthase. Proc. Natl. Acad. Sci. USA 2008, 105, 13883–13888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujishiro, T.; Shoji, O.; Nagano, S.; Sugimoto, H.; Shiro, Y.; Watanabe, Y. Crystal structure of H2O2-dependent cytochrome P450 SPα with its bound fatty acid substrate: Insight into the regioselective hydroxylation of fatty acids at the α position. J. Biol. Chem. 2011, 286, 29941–29950. [Google Scholar] [CrossRef] [PubMed]
- Pao, G.M.; Saier, M.H. Response regulators of bacterial signal transduction systems: Selective domain shuffling during evolution. J. Mol. Evol. 1995, 40, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Galperin, M.Y.; Nikolskaya, A.N. Identification of sensory and signal-transducing domains in two-component signaling systems. Met. Enzymol. 2007, 422, 47–74. [Google Scholar]
- Orth, P.; Schnappinger, D.; Hillen, W.; Saenger, W.; Hinrichs, W. Structural basis of gene regulation by the tetracycline inducible Tet repressor-operator system. Nat. Struct. Biol. 2000, 7, 215–219. [Google Scholar] [PubMed]
- Ramos, J.L.; Martínez-Bueno, M.; Molina-Henares, A.J.; Terán, W.; Watanabe, K.; Zhang, X.; Gallegos, M.T.; Brennan, R.; Tobes, R. The TetR family of transcriptional repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–356. [Google Scholar] [CrossRef]
- Martinez-Bueno, M.; Molina-Henares, A.J.; Pareja, E.; Ramos, J.L.; Tobes, R. BacT regulators: A database of transcriptional regulators in bacteria and archaea. Bioinformatics 2004, 20, 2787–2791. [Google Scholar] [CrossRef]
- Li, M.; Gu, R.; Su, C.C.; Routh, M.D.; Harris, K.C.; Jewell, E.S.; McDermott, G.; Edward, W.Y. Crystal structure of the transcriptional regulator AcrR from Escherichia coli. J. Mol. Biol. 2007, 374, 591–603. [Google Scholar] [CrossRef]
- Su, C.C.; Rutherford, D.J.; Yu, E.W. Characterization of the multidrug efflux regulator AcrR from Escherichia coli. Biochem. Biophys. Res. Commun. 2007, 361, 85–90. [Google Scholar] [CrossRef]
- Bruner, S.D.; Norman, D.P.; Verdine, G.L. Structural basis for recognition and repair of the endogenous mutagen 8-oxoguanine in DNA. Nature 2000, 403, 859. [Google Scholar] [CrossRef] [PubMed]
- Nash, H.M.; Bruner, S.D.; Schärer, O.D.; Kawate, T.; Addona, T.A.; Spooner, E.; Lane, W.S.; Verdine, G.L. Cloning of a yeast 8-oxoguanine DNA glycosylase reveals the existence of a base-excision DNA-repair protein superfamily. Curr. Biol. 1996, 6, 968–980. [Google Scholar] [CrossRef]
- Friedhoff, P.; Gimadutdinow, O.; Pingoud, A. Identification of catalytically relevant amino acids of the extracellular Serratia marcescens endonuclease by alignment-guided mutagenesis. Nucleic Acid Res. 1994, 22, 3280–3287. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.D.; Tanner, J.; Alpaugh, M.; Benedik, M.J.; Krause, K.L. 2. 1 Å structure of Serratia endonuclease suggests a mechanism for binding to double-stranded DNA. Nat. Struct. Mol. Biol. 1994, 1, 461. [Google Scholar] [CrossRef]
- Schleif, R. AraC protein, regulation of the l-arabinose operon in Escherichia coli, and the light switch mechanism of AraC action. FEMS Microbiol. Rev. 2010, 34, 779–796. [Google Scholar] [CrossRef]
- Rodgers, M.; Schleif, R. DNA tape measurements of AraC. Nucleic Acids Res. 2008, 36, 404–410. [Google Scholar] [CrossRef]
- Ollis, D.L.; Cheah, E.; Cygler, M.; Dijkstra, B.; Frolow, F.; Franken, S.M.; Harel, M.; Remington, S.J.; Silman, I.; Schrag, J.; et al. The α/β hydrolase fold. Protein Eng. 1992, 5, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Nardini, M.; Dijkstra, B.W. α/β hydrolase fold enzymes: The family keeps growing. Curr. Opin. Struct. Biol. 1999, 9, 732–737. [Google Scholar] [CrossRef]
- Carr, P.D.; Ollis, D.L. α/β hydrolase fold: An update. Protein Pept. Lett. 2009, 16, 1137–1148. [Google Scholar]
- Goodacre, N.F.; Gerloff, D.L.; Uetz, P. Protein domains of unknown function are essential in bacteria. MBio 2014, 5, e00744-13. [Google Scholar] [CrossRef]
- Mandiyan, V.; Andreev, J.; Schlessinger, J.; Hubbard, S.R. Crystal structure of the ARF-GAP domain and ankyrin repeats of PYK2-associated protein β. EMBO J. 1999, 18, 6890–6898. [Google Scholar] [CrossRef] [PubMed]
- Lux, S.E.; John, K.M.; Bennett, V. Analysis of cDNA for human erythrocyte ankyrin indicates a repeated structure with homology to tissue-differentiation and cell-cycle control proteins. Nature 1990, 344, 36. [Google Scholar] [CrossRef] [PubMed]
- Bork, P. Hundreds of ankyrin-like repeats in functionally diverse proteins: Mobile modules that cross phyla horizontally? Proteins 1993, 17, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Bork, P.; Doolittle, R.F. Proposed acquisition of an animal protein domain by bacteria. Proc. Natl. Acad. Sci. USA 1992, 89, 8990–8994. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.H.; Otto, E.; Bennett, V. Specific 33-residue repeat (s) of erythrocyte ankyrin associate with the anion exchanger. J. Biol. Chem. 1991, 266, 11163–11169. [Google Scholar]
- Hall, A. Rho GTPases and the actin cytoskeleton. Science 1998, 279, 509–514. [Google Scholar] [CrossRef]
- Crespo, P.; Schuebel, K.E.; Ostrom, A.A.; Gutkind, J.S.; Bustelo, X.R. Phosphotyrosine-dependent activation of Rac-1 GDP/GTP exchange by the vav proto-oncogene product. Nature 1997, 385, 169. [Google Scholar] [CrossRef] [PubMed]
- Hart, M.J.; Eva, A.; Evans, T.; Aaronson, S.A.; Cerione, R.A. Catalysis of guanine nucleotide exchange on the CDC42Hs protein by the dbl oncogene product. Nature 1991, 354, 311. [Google Scholar] [CrossRef]
- Takano, H.; Komuro, I.; Oka, T.; Shiojima, I.; Hiroi, Y.; Mizuno, T.; Yazaki, Y. The Rho family G proteins play a critical role in muscle differentiation. Mol. Cell. Biol. 1998, 18, 1580–1589. [Google Scholar] [CrossRef]
- Moorman, J.P.; Luu, D.; Wickham, J.; Bobak, D.A.; Hahn, C.S. A balance of signaling by Rho family small GTPases RhoA, Rac1 and Cdc42 coordinates cytoskeletal morphology but not cell survival. Oncogene 1999, 18, 47. [Google Scholar] [CrossRef]
- Kaibuchi, K.; Kuroda, S.; Amano, M. Regulation of the cytoskeleton and cell adhesion by the Rho family GTPases in mammalian cells. Ann. Rev. Biochem. 1999, 68, 459–486. [Google Scholar] [CrossRef] [PubMed]
- Chimini, G.; Chavrier, P. Function of Rho family proteins in actin dynamics during phagocytosis and engulfment. Nat. Cell Biol. 2000, 2, 191. [Google Scholar] [CrossRef] [PubMed]
- Prokopenko, S.N.; Saint, R.; Bellen, H.J. Untying the Gordian knot of cytokinesis: Role of small G proteins and their regulators. J. Cell Biol. 2000, 148, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Somlyo, A.P.; Somlyo, A.V. Signal transduction by G-proteins, rho-kinase and protein phosphatase to smooth muscle and non-muscle myosin II. J. Physiol. 2000, 522, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Uehata, M.; Ishizaki, T.; Satoh, H.; Ono, T.; Kawahara, T.; Morishita, T.; Tamakawa, H.; Yamagami, K.; Inui, J.; Maekawa, M.; et al. Calcium sensitization of smooth muscle mediated by a Rho-associated protein kinase in hypertension. Nature 1997, 389, 990. [Google Scholar] [CrossRef]
- Zohn, I.M.; Campbell, S.L.; Khosravi-Far, R.; Rossman, K.L.; Der, C.J. Rho family proteins and Ras transformation: The RHOad less traveled gets congested. Oncogene 1998, 17, 1415. [Google Scholar] [CrossRef] [PubMed]
- Nishida, K.; Kaziro, Y.; Satoh, T. Association of the proto-oncogene product Dbl with G protein βγ subunits. FEBS Lett. 1999, 459, 186–190. [Google Scholar] [CrossRef]
- Cerione, R.A.; Zheng, Y. The Dbl family of oncogenes. Curr. Opin. Cell Biol. 1996, 8, 216–222. [Google Scholar] [CrossRef]
- Longenecker, K.L.; Lewis, M.E.; Chikumi, H.; Gutkind, J.S.; Derewenda, Z.S. Structure of the RGS-like domain from PDZ-RhoGEF: Linking heterotrimeric g protein-coupled signaling to Rho GTPases. Structure 2001, 9, 559–569. [Google Scholar] [CrossRef]
- Ponting, C.P. Evidence for PDZ domains in bacteria, yeast, and plants. Protein Sci. 1997, 6, 464–468. [Google Scholar] [CrossRef] [Green Version]
- Ponting, C.P.; Phillips, C.; Davies, K.E.; Blake, D.J. PDZ domains: Targeting signaling molecules to sub-membranous sites. Bioessays 1997, 19, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Grootjans, J.J.; Zimmermann, P.; Reekmans, G.; Smets, A.; Degeest, G.; Dürr, J.; David, G. Syntenin, a PDZ protein that binds syndecan cytoplasmic domains. Proc. Natl. Acad. Sci. USA 1997, 94, 13683–13688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohr, D.; Venkov, P.; Zlatanova, J. Transcriptional regulation in the yeast GAL gene family: A complex genetic network. FASEB J. 1995, 9, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Marmorstein, R.; Carey, M.; Ptashne, M.; Harrison, S.C. DNA recognition by GAL4: Structure of a protein-DNA complex. Nature 1992, 356, 408. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Coleman, J.E. GAL4 transcription factor is not a “zinc finger” but forms a Zn (II) 2Cys6 binuclear cluster. Proc. Natl. Acad. Sci. USA 1990, 87, 2077–2081. [Google Scholar] [CrossRef] [PubMed]
- Todd, R.B.; Andrianopoulos, A. Evolution of a fungal regulatory gene family: The Zn (II) 2Cys6 binuclear cluster DNA binding motif. Fungal Genet. Biol. 1997, 21, 388–405. [Google Scholar] [CrossRef] [PubMed]
- Pathak, G.P.; Vrana, J.D.; Tucker, C.L. Optogenetic control of cell function using engineered photoreceptors. Bio. Cell 2013, 105, 59–72. [Google Scholar] [CrossRef]
- Fiedler, B.; Broc, D.; Schubert, H.; Rediger, A.; Börner, T.; Wilde, A. Involvement of cyanobacterial phytochromes in growth under different light qualities and quantities. Photochem. Photobiol. 2004, 79, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.J.; Rockwell, N.C.; Jang, A.Y.; Ernst, L.A.; Waggoner, A.S.; Duan, Y.; Lei, H.; Lagarias, J.C. Multiple roles of aconserved GAF domain tyrosine residue in cyanobacterial and plant phytochromes. Biochemistry 2005, 44, 15203–15215. [Google Scholar] [CrossRef]
- Kehoe, D.M.; Grossman, A.R. New classes of mutants in complementary chromatic adaptation provide evidence for a novel four-step phosphorelay system. J. Bacteriol. 1997, 179, 3914–3921. [Google Scholar] [CrossRef] [Green Version]
- Moon, Y.J.; Park, Y.M.; Chung, Y.H.; Choi, J.S. Calcium is involved in photomovement of cyanobacterium Synechocystis sp. PCC 6803. Photochem. Photobiol. 2004, 79, 114–119. [Google Scholar] [CrossRef]
- Hong, K.P.; Spitzer, N.C.; Nicol, X. Improved molecular toolkit for cAMP studies in live cells. BMC Res. Notes 2011, 4, 241. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, S.; Katayama, M.; Geng, X.; Ikeuchi, M. Cyanobacterial phytochrome-like PixJ1 holoprotein shows novel reversible photoconversion between blue and green-absorbing forms. Plant Cell Physiol. 2004, 45, 1729–1737. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, S.; Shimada, T.; Matsuoka, D.; Zikihara, K.; Kohchi, T.; Tokutomi, S. Reconstitution of blue-green reversible photoconversion of a cyanobacterial photoreceptor, PixJ1, in phycocyanobilin-producing Escherichia coli. Biochemistry 2006, 45, 3775–3784. [Google Scholar] [CrossRef]
- Kehoe, D.M.; Grossman, A.R. Similarity of achromatic adaptation sensor to phytochrome and ethylene receptors. Science 1996, 273, 1409–1412. [Google Scholar] [CrossRef] [PubMed]
- Schröder-Lang, S.; Schwärzel, M.; Seifert, R.; Strünker, T.; Kateriya, S.; Looser, J.; Watanabe, M.; Kaupp, U.B.; Hegemann, P.; Nagel, G. Fast manipulation of cellular cAMP level by light in vivo. Nat. Methods 2007, 4, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, T.; Suzuki, T.; Yoshikawa, S.; Iseki, M. Functional transplant of photoactivated adenylyl cyclase (PAC) into Aplysia sensory neurons. Neurosci. Res. 2007, 59, 81–88. [Google Scholar] [CrossRef]
- Weissenberger, S.; Schultheis, C.; Liewald, J.F.; Erbguth, K.; Nagel, G.; Gottschalk, A. PACα—An optogenetic tool for in vivo manipulation of cellular cAMP levels, neurotransmitter release and behavior in Caenorhabditis elegans. J. Neurochem. 2011, 116, 616–625. [Google Scholar] [CrossRef]
- Cheng, Z.; Li, K.; Hammad, L.A.; Karty, J.A.; Bauer, C.E. Vitamin B12 regulates photosystem gene expression via the CrtJantirepressorAerR in Rhodobactercapsulatus. Mol. Microbiol. 2014, 91, 649–664. [Google Scholar] [CrossRef]
- Zoltowski, B.D.; Vaccaro, B.; Crane, B.R. Mechanism-based tuning of a LOV domain photoreceptor. Nat. Chem. Biol 2009, 5, 827–834. [Google Scholar] [CrossRef]
- Stephan, E.; Granzin, J.; Circolone, F.; Stadler, A.; Krauss, U.; Drepper, T.; Svensson, V.; Knieps-Grünhagen, E.; Wirtz, A.; Cousin, A.; et al. Structure and function of a short LOV protein from the marine phototrophic bacterium Dinoroseobactershibae. BMC Microbiol. 2015, 15, 30. [Google Scholar]
- Pillai, V.R. Photoremovable protecting groups in organic synthesis. Synthesis 1980, 1980, 1–26. [Google Scholar] [CrossRef]
- Cardin, J.A.; Carlén, M.; Meletis, K.; Knoblich, U.; Zhang, F.; Deisseroth, K.; Tsai, L.H.; Moore, C.I. Targeted optogenetic stimulation and recording of neurons in vivo using cell-type-specific expression of Channelrhodopsin-2. Nat. Protoc. 2010, 5, 247. [Google Scholar] [CrossRef] [PubMed]
- Levskaya, A.; Weiner, O.D.; Lim, W.A.; Voigt, C.A. Spatiotemporal control of cell signaling using a light-switchable protein interaction. Nature 2009, 461, 997. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, M.J.; Hughes, R.M.; Peteya, L.A.; Schwartz, J.W.; Ehlers, M.; Tucker, C.L. Rapid blue-light–mediated induction of protein interactions in living cells. Nat. Methods 2010, 7, 973. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Domains | Accession No. | BLUF Length | Score | Organisms | Probable Modulations |
|---|---|---|---|---|---|
| BLUF + EAL | ARH96915.1 | 2–91 | e−28 | Escherichia coli | Regulate diguanylate cyclases and Phosphodiesterase activity |
| BLUF + PsiE | EGW22399.1 | 2–92 | e−36 | Methylobacter tundripaludum SV96 | NP* |
| BLUF + CHD | EHQ08139.1 | 3–93 | e−28 | Leptonema illini DSM 21528 | Regulate adenylyl and guanylyl cyclase activity |
| BLUF + B12 Binding domain | ABP71929.1 | 17–106 | e−35 | Rhodobacter sphaeroides ATCC 17025 | Broadens BLUF photosensing ability |
| BLUF + PRK09039 superfamily | WP_012321331.1 | 5–99 | e−36 | Methylobacterium radiotolerans | Regulate phosphoribulokinase, uridine kinase and pantothenate kinase activity |
| BLUF + DNA_pol3_gamma3 superfamily | AIQ92835.1 | 5–99 | e−36 | Methylobacterium oryzae CBMB 20 | Regulate DNA replication process |
| BLUF + REC | AMR27912.1 | 150–240 | e−37 | Hymenobacter sp. PAMC 26554 | Regulate bacterial chemotaxis |
| BLUF + P450 superfamily | WP_045444510.1 | 15–108 | e−35 | Psychrobacter sp. P11F6 | Regulate oxidative degradation of steroids, fatty acid and xenobiotics |
| BLUF + AcrR | WP_058726129.1 | 3–94 | e−28 | Curtobacterium luteum | Regulate antibiotic resistance in bacteria |
| BLUF + TetR_C_6 | WP_051596720.1 | 4–92 | e−34 | Curtobacterium sp. UNCCL17 | Regulate antibiotic resistance in bacteria |
| 343–417 | e−19 | ||||
| BLUF +Endonuclease_NS | WP_058743091.1 | 17–83 | e−30 | Drosophila eracta | Modulate hydrolase activity, nucleic acid and metal ion binding |
| BLUF + PAS | WP_058511962.1 | 4–88 | e−13 | Legionella steelei | Regulate cellular signaling processes |
| BLUF + AraC | WP_053973760.1 | 228–318 | e−19 | Polaribacter dokdonensis | Control synthesis of structural components of arabinose metabolism |
| BLUF + Abhydrolase super family | XP_008692928.1 | 58–99 | e−15 | Ursus maritimus | Modulate hydrolytic enzyme activity |
| BLUF + ENDO3c Superfamily | EYD78138.1 | 1–89 | e−27 | Rubellimicrobium mesophilum DSM 19309 | Control DNA repair regulation |
| BLUF + ANK | EJY80769.1 | 547–593 | e−13 | Oxytricha trifallax | Modulate protein-protein interaction |
| BLUF + DUF1115 Superfamily | WP_0229622806.1 | 5–101 | e−37 | Pseudomonas pelagia | NP* |
| BLUF + RhoGEFSuperfamily | XP_025342216.1 | 171–262 | e−38 | Pseudomicrostroma glucosiphilum | Control activation of Rho family GTPases |
| BLUF + PDZ | jgi_Bigna1_85551 | 55–144 | e−17 | Bigelowiella natans CCMP 2755 | Regulate membrane-bound cell signaling |
| BLUF + AANH_like Superfamily | Jgi_Schag1_101311 | 273–366 | e−24 | Schizochytrium aggregatum ATCC 28209 | NP* |
| BLUF + EAL + GGDEF | AFL74487.1 | 456–546 | e−33 | Thiocystis violascens DSM 198 | Regulation of c-di-GMP level |
| BLUF + EAL+ PRK15043 superfamily | CDW60191.1 | 2–62 | e−25 | Trichuris trichuris | NP* |
| BLUF + GGDEF+ PAS | WP_058465269.1 | 1–91 | e−25 | Legionella cincinnatiensis | NP* |
| BLUF + GGDEF+ COG5001 Superfamily + PAS | WP_058516015.1 | 3–85 | e−23 | Legionella santicrucis | NP* |
| BLUF + COG5001 + PBP1_NHase + PAS | ADC61983.1 | 974–1064 | e−38 | Allochromatium vinosum DSM 180 | NP* |
| BLUF + PsiE +BaeS superfamily | WP_014148160.1 | 3–93 | e−42 | Methylomicrobium alcaliphilum | NP* |
| BLUF + CHD + LRR_R1 Superfamily | Q8S9F2.1 | 57–136 | e−24 | Euglena gracilis | NP* |
| 468–554 | e−27 | ||||
| BLUF + CHD + Med 26_M Superfamily | XP_013758351.1 | 98–186 | e−19 | Thecamonas trahens ATCC 50062 | NP* |
| 673–756 | e−18 | ||||
| BLUF + PRK11633 + DNA pol3 gamma3 family | WP_048452447.1 | 5–99 | e−42 | Methylobacterium tarhaniae | NP* |
| BLUF + TetR_C_11 family + Fer2_2 superfamily | WP_058743091.1 | 4–92 | e−42 | Curtobacterium citreum | NP* |
| BLUF + GAL4 + Fungal TF MHR | ORY86082.1 | 10–100 | e−38 | Protomyces inouyei | Regulate galactose induced genes |
| BLUF + SRPBCC Superfamily + RGS Superfamily | BAV14116.1 | 518–612 | e−26 | Naegleria fowleri | NP* |
| BLUF + SRPBCC + BTB + DUF35522 Superfamily | EFC49155.1 | 1132–1226 | e−28 | Naegleria gruberi | NP* |
| BLUF + SRPBCC + FH2 + Drf_FH1 + PRK13729 + SMC_N | XP_002669619.1 | 1971–2050 | e−28 | Naegleria gruberi | NP* |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaushik, M.S.; Sharma, R.; Veetil, S.K.; Srivastava, S.K.; Kateriya, S. Modular Diversity of the BLUF Proteins and Their Potential for the Development of Diverse Optogenetic Tools. Appl. Sci. 2019, 9, 3924. https://doi.org/10.3390/app9183924
Kaushik MS, Sharma R, Veetil SK, Srivastava SK, Kateriya S. Modular Diversity of the BLUF Proteins and Their Potential for the Development of Diverse Optogenetic Tools. Applied Sciences. 2019; 9(18):3924. https://doi.org/10.3390/app9183924
Chicago/Turabian StyleKaushik, Manish Singh, Ramandeep Sharma, Sindhu Kandoth Veetil, Sandeep Kumar Srivastava, and Suneel Kateriya. 2019. "Modular Diversity of the BLUF Proteins and Their Potential for the Development of Diverse Optogenetic Tools" Applied Sciences 9, no. 18: 3924. https://doi.org/10.3390/app9183924