
Matrix Stiffness Influences Tubular Formation in Renal Tissue Engineering
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
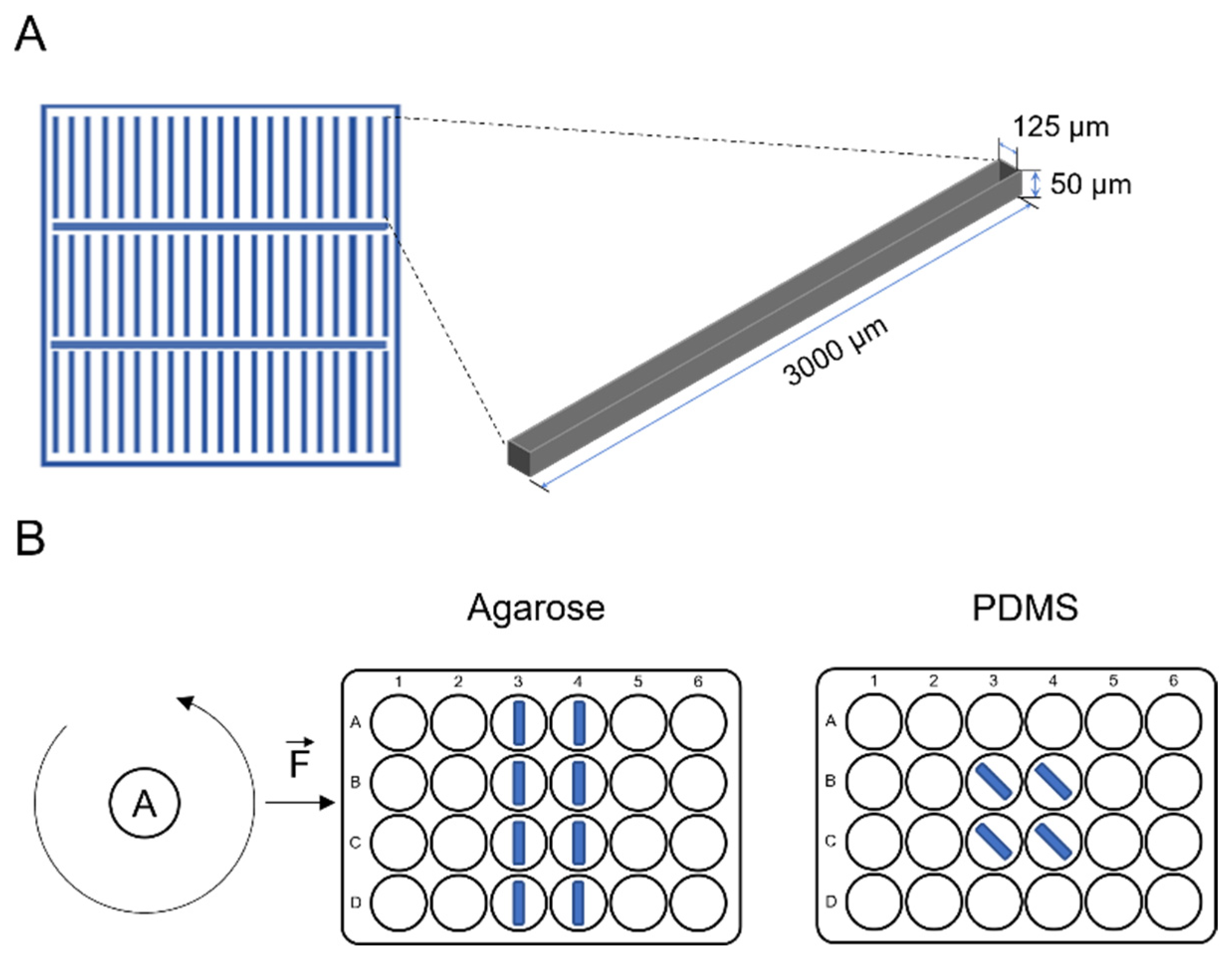
2.1. Measuring Physical Characteristics of Agarose and PDMS Molds
2.2. SU-8 Mold Fabrication
2.3. Polymer and Agarose Based Micropatterned Mold
2.4. Cells and Cell culture
2.5. Tubular Formation
2.6. Confocal Microscopy
2.7. Scoring of Tubular Formation and Statistic Analysis
3. Results
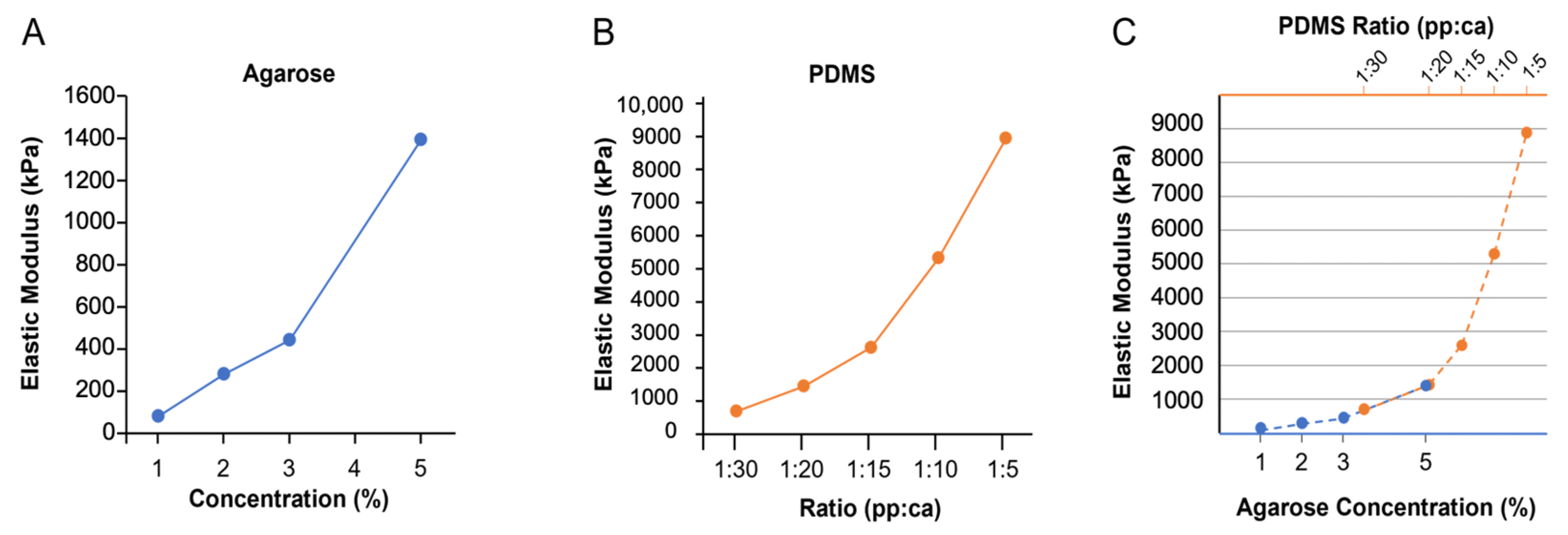
3.1. Polymer Characteristics
3.2. Cell Seeding
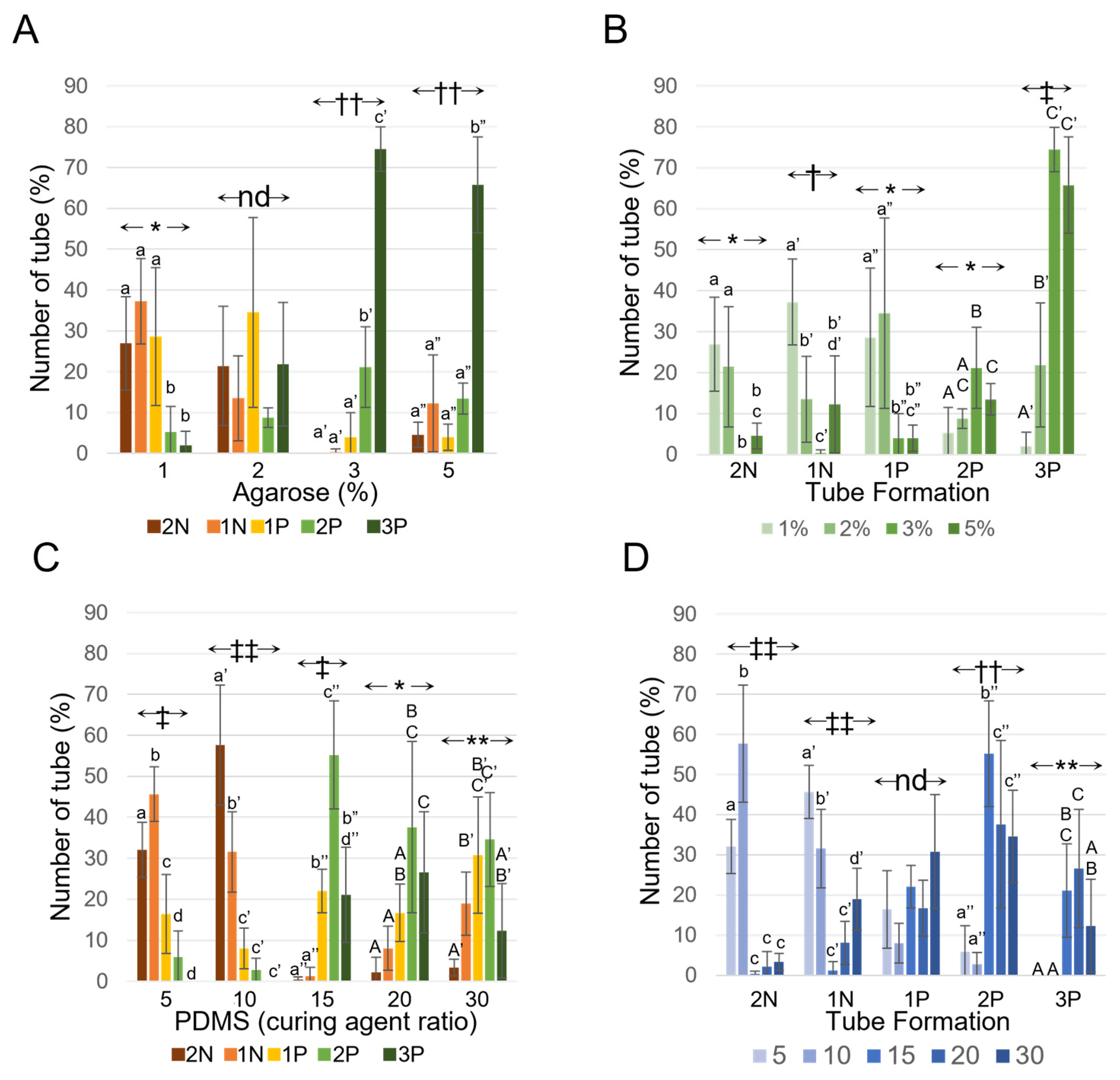
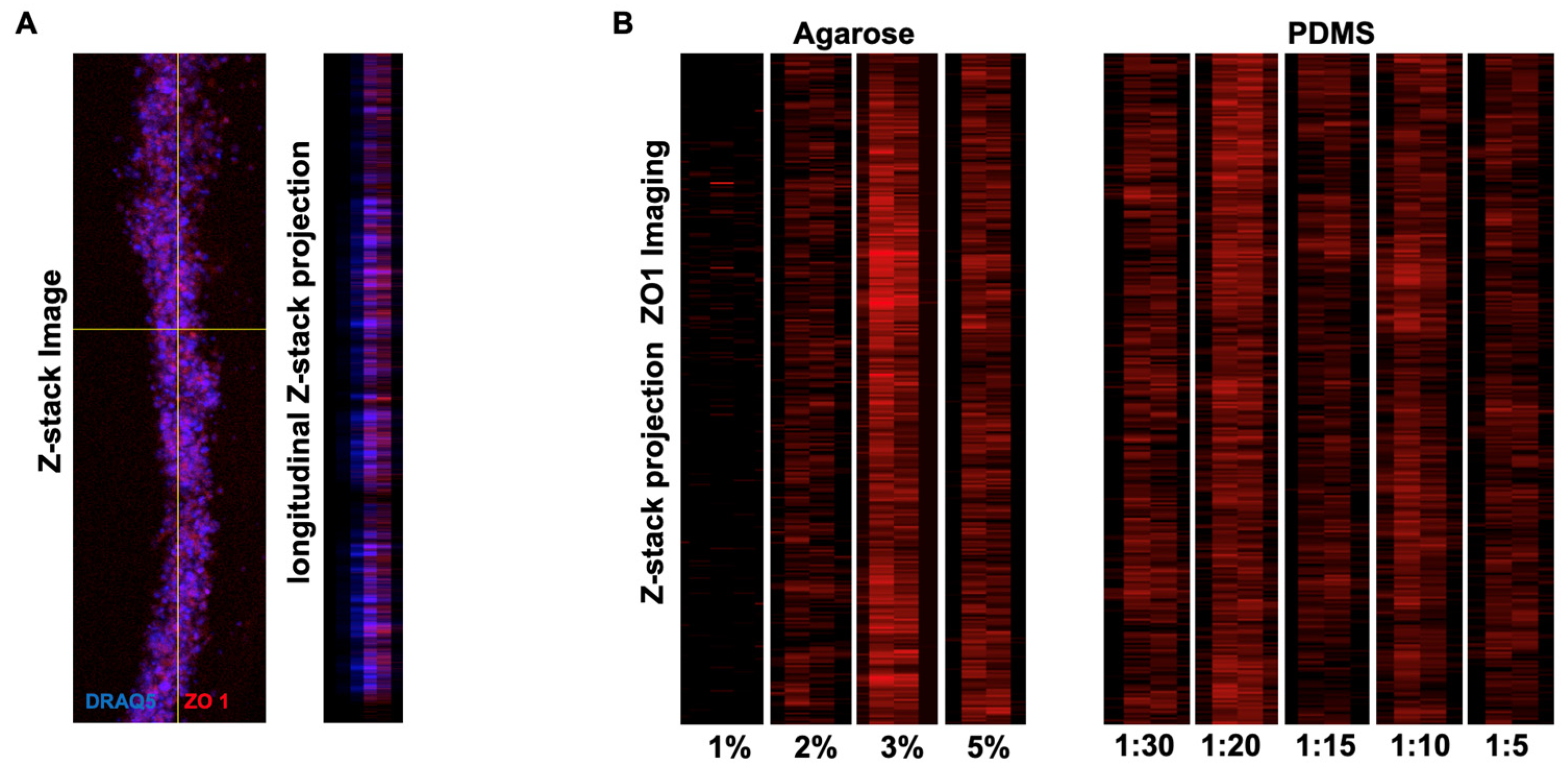
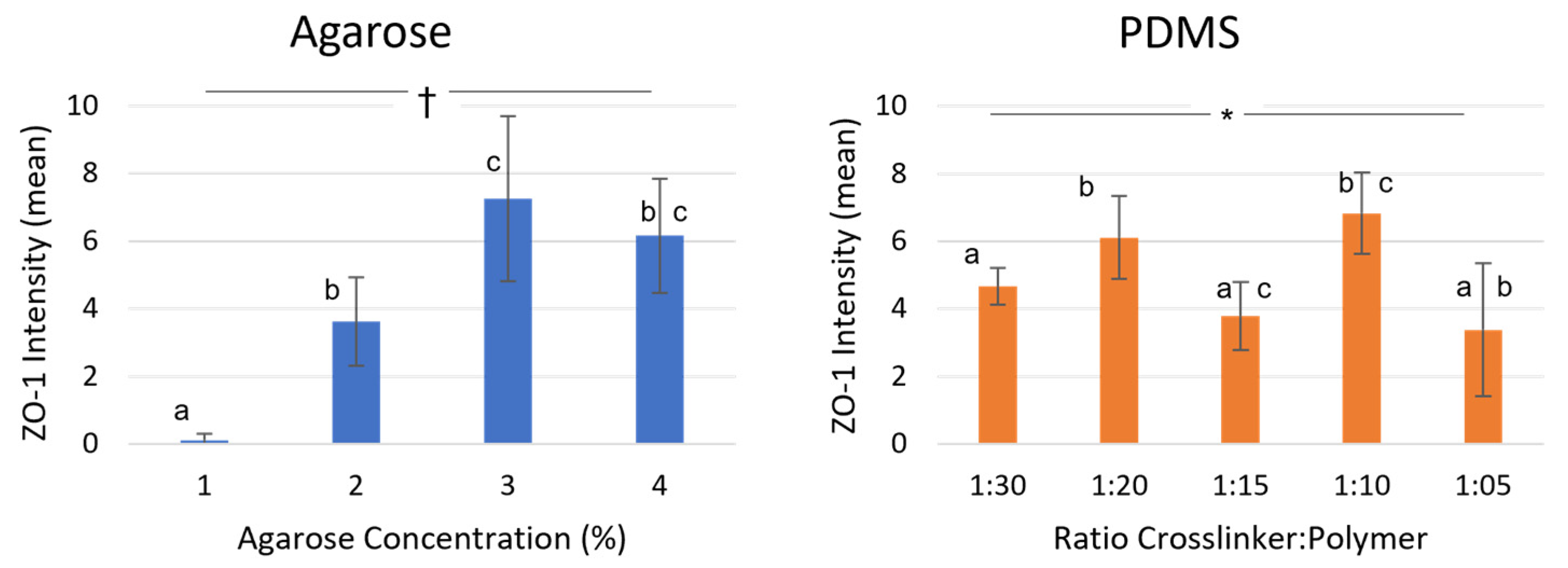
3.3. Tube Formation on Agarose
3.4. Tube Formation on PDMS
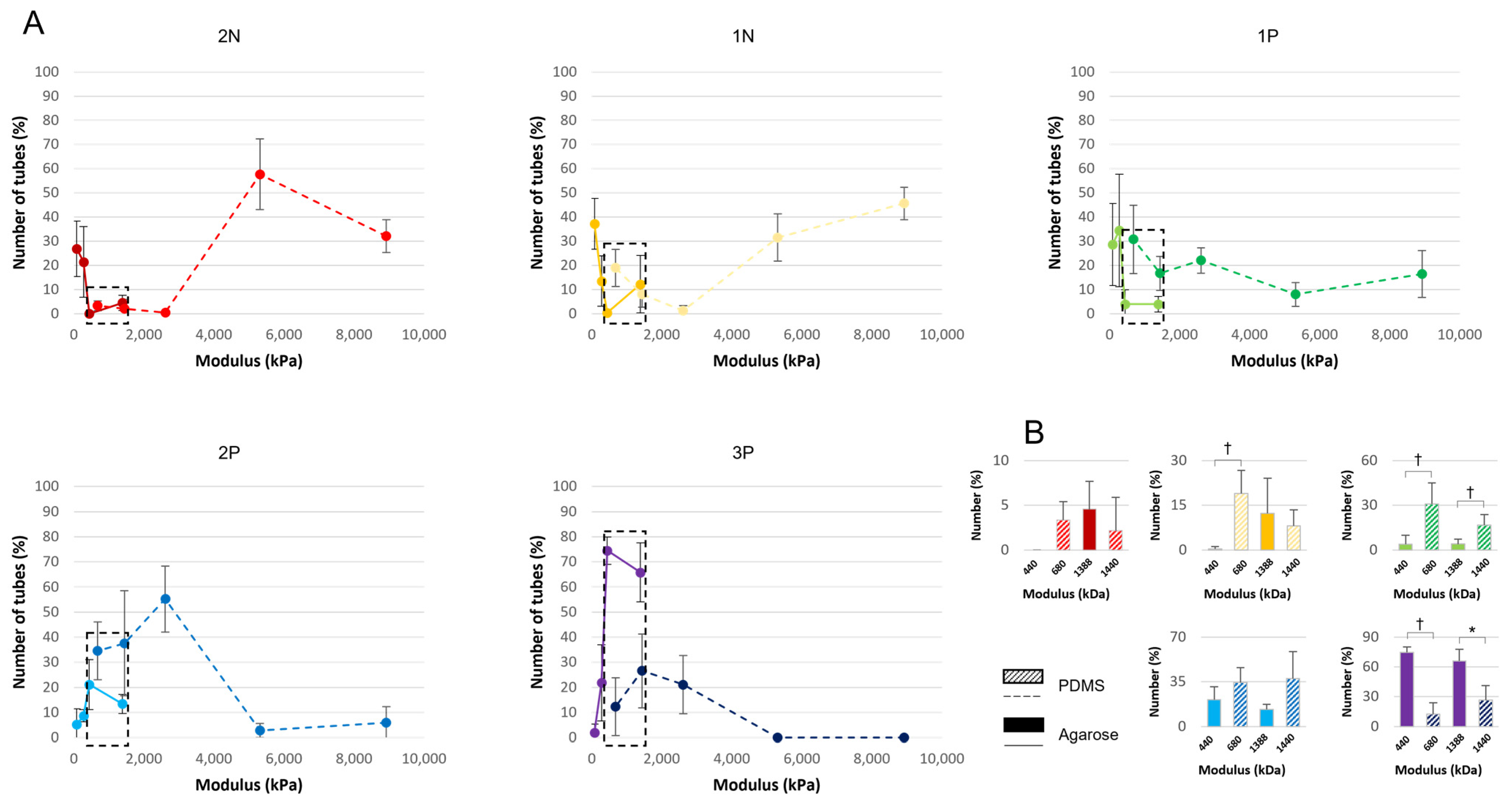
3.5. Comparison of Both Materials with Similar Stiffness
3.6. Polarization Marker
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moharamzadeh, K.; Colley, H.; Hearnden, V.; Murdoch, C. 15-Tissue-engineered models of oral soft tissue diseases. In Biomaterials for Oral and Dental Tissue Engineering; Tayebi, L., Moharamzadeh, K., Eds.; Woodhead Publishing: Sawston, UK, 2017; pp. 245–255. [Google Scholar] [CrossRef]
- Williams, J.K.; Yoo, J.J.; Atala, A. Chapter 59-Regenerative Medicine Approaches for Tissue Engineered Heart Valves. In Principles of Regenerative Medicine, 3rd ed.; Atala, A., Lanza, R., Mikos, A.G., Nerem, R., Eds.; Academic Press: Boston, MA, USA, 2019; pp. 1041–1058. [Google Scholar] [CrossRef]
- Tan, J.K.H.; Watanabe, T. Chapter 5-Artificial Engineering of Secondary Lymphoid Organs. In Advances in Immunology; Alt, F.W., Ed.; Academic Press: Cambridge, MA, USA, 2010; pp. 131–157. [Google Scholar] [CrossRef]
- Nelson, W.J. Tube morphogenesis: Closure, but many openings remain. Trends Cell Biol. 2003, 13, 615–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marciano, D.K. A holey pursuit: Lumen formation in the developing kidney. Pediatr. Nephrol. Berl. Ger. 2017, 32, 7–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlüter, M.A.; Margolis, B. Apical Lumen Formation in Renal Epithelia. J. Am. Soc. Nephrol. 2009, 20, 1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Zimmerman, S.; Brakeman, P.R.; Beaudoin, G.M., 3rd; Reichardt, L.F.; Marciano, D.K. De novo lumen formation and elongation in the developing nephron: A central role for afadin in apical polarity. Dev. Camb. Engl. 2013, 140, 1774–1784. [Google Scholar] [CrossRef] [Green Version]
- Orabi, H.; AbouShwareb, T.; Zhang, Y.; Yoo, J.J.; Atala, A. Cell-seeded tubularized scaffolds for reconstruction of long urethral defects: A preclinical study. Eur. Urol. 2013, 63, 531–538. [Google Scholar] [CrossRef] [Green Version]
- Nelson, C.M.; Vanduijn, M.M.; Inman, J.L.; Fletcher, D.A.; Bissell, M.J. Tissue geometry determines sites of mammary branching morphogenesis in organotypic cultures. Science 2006, 314, 298–300. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, S.; Nelson, C.M.; Baranski, J.D.; Lim, E.; Chen, C.S. Geometrically controlled endothelial tubulogenesis in micropatterned gels. Tissue Eng. Part A 2010, 16, 2255–2263. [Google Scholar] [CrossRef] [Green Version]
- Hauser, P.V.; Nishikawa, M.; Kimura, H.; Fujii, T.; Yanagawa, N. Controlled tubulogenesis from dispersed ureteric bud-derived cells using a micropatterned gel. J. Tissue Eng. Regen. Med. 2016, 10, 762–771. [Google Scholar] [CrossRef] [Green Version]
- Contessi Negrini, N.; Angelova Volponi, A.; Higgins, C.A.; Sharpe, P.T.; Celiz, A.D. Scaffold-based developmental tissue engineering strategies for ectodermal organ regeneration. Mater. Today Bio 2021, 10, 100107. [Google Scholar] [CrossRef]
- Ouyang, M.; Yu, J.-Y.; Chen, Y.; Deng, L.; Guo, C.-L. Cell-extracellular matrix interactions in the fluidic phase direct the topology and polarity of self-organized epithelial structures. Cell Prolif. 2021, 54, e13014. [Google Scholar] [CrossRef]
- Sacharidou, A.; Stratman, A.N.; Davis, G.E. Molecular mechanisms controlling vascular lumen formation in three-dimensional extracellular matrices. Cells Tissues Organs. 2012, 195, 122–143. [Google Scholar] [CrossRef] [Green Version]
- Łabowska, M.B.; Cierluk, K.; Jankowska, A.M.; Kulbacka, J.; Detyna, J.; Michalak, I. A Review on the Adaption of Alginate-Gelatin Hydrogels for 3D Cultures and Bioprinting. Mater. Basel. Switz. 2021, 14, 858. [Google Scholar] [CrossRef]
- Kinoshita, K.; Iwase, M.; Yamada, M.; Yajima, Y.; Seki, M. Fabrication of multilayered vascular tissues using microfluidic agarose hydrogel platforms. Biotechnol. J. 2016, 11, 1415–1423. [Google Scholar] [CrossRef]
- Discher, D.E.; Janmey, P.; Wang, Y.L. Tissue cells feel and respond to the stiffness of their substrate. Science 2005, 310, 1139–1143. [Google Scholar] [CrossRef] [Green Version]
- Ichanti, H.; Sladic, S.; Kalies, S.; Haverich, A.; Andrée, B.; Hilfiker, A. Characterization of Tissue Engineered Endothelial Cell Networks in Composite Collagen-Agarose Hydrogels. Gels. Basel. Switz. 2020, 6, 27. [Google Scholar] [CrossRef]
- Alderfer, L.; Russo, E.; Archilla, A.; Coe, B.; Hanjaya-Putra, D. Matrix stiffness primes lymphatic tube formation directed by vascular endothelial growth factor-C. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2021, 35, e21498. [Google Scholar] [CrossRef]
- Mori, Y.; Kanazawa, S.; Watanabe, M.; Suenaga, H.; Okubo, K.; Nagata, S.; Fujihara, Y.; Takato, T.; Hoshi, K. Usefulness of Agarose Mold as a Storage Container for Three-Dimensional Tissue-Engineered Cartilage. Mater. Sci. Appl. 2013, 4, 733–778. [Google Scholar] [CrossRef] [Green Version]
- Moustafa, M.E.; Gadepalli, V.S.; Elmak, A.A.; Lee, W.; Rao, R.R.; Yadavalli, V.K. Large area micropatterning of cells on polydimethylsiloxane surfaces. J. Biol. Eng. 2014, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Odenwald, M.A.; Choi, W.; Buckley, A.; Shashikanth, N.; Joseph, N.E.; Wang, Y.; Warren, M.H.; Buschmann, M.M.; Pavlyuk, R.; Hildebrand, J.; et al. ZO-1 interactions with F-actin and occludin direct epithelial polarization and single lumen specification in 3D culture. J. Cell Sci. 2017, 130, 243–259. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, R.; Sherratt, M.J.; Cruickshank, J.K.; Derby, B. Characterizing the elastic properties of tissues. Mater. Today 2011, 14, 96–105. [Google Scholar] [CrossRef]
- Butcher, D.T.; Alliston, T.; Weaver, V.M. A tense situation: Forcing tumor progression. Nat. Rev. Cancer 2009, 9, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Levental, I.; Georges, P.C.; Janmey, P.A. Soft biological materials and their impact on cell function. Soft Matter. 2007, 3, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Ogle, M.E.; Doron, G.; Levy, M.J.; Temenoff, J.S. Hydrogel Culture Surface Stiffness Modulates Mesenchymal Stromal Cell Secretome and Alters Senescence. Tissue Eng. Part A 2020, 26, 1259–1271. [Google Scholar] [CrossRef] [PubMed]
- Koser, D.E.; Thompson, A.J.; Foster, S.K.; Dwivedy, A.; Pillai, E.K.; Sheridan, G.K.; Svoboda, H.; Viana, M.; da F Costa, L.; Address, J.; et al. Mechanosensing is critical for axon growth in the developing brain. Nat. Neurosci. 2016, 19, 1592–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skeath, J.B.; Wilson, B.A.; Romero, S.E.; Snee, M.J.; Zhu, Y.; Lacin, H. The extracellular metalloprotease AdamTS-A anchors neural lineages in place within and preserves the architecture of the central nervous system. Dev. Camb. Engl. 2017, 144, 3102–3113. [Google Scholar] [CrossRef] [Green Version]
- Khalilgharibi, N.; Mao, Y. To form and function: On the role of basement membrane mechanics in tissue development, homeostasis and disease. Open Biol. 2021, 11, 200360. [Google Scholar] [CrossRef]
- Reuten, R.; Zendehroud, S.; Nicolau, M.; Fleischhauer, L.; Laitala, A.; Kiderlen, S.; Nikodemus, D.; Wullkopf, L.; Nielsen, S.R.; McNeilly, S.; et al. Basement membrane stiffness determines metastases formation. Nat. Mater. 2021, 20, 892–903. [Google Scholar] [CrossRef]
- Hanjaya-Putra, D.; Yee, J.; Ceci, D.; Truitt, R.; Yee, D.; Gerecht, S. Vascular endothelial growth factor and substrate mechanics regulate in vitro tubulogenesis of endothelial progenitor cells. J. Cell Mol. Med. 2010, 14, 2436–2447. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-C.; Lin, H.-H.; Tang, M.-J. Regulation of proximal tubular cell differentiation and proliferation in primary culture by matrix stiffness and ECM components. Am. J. Physiol. Renal. Physiol. 2014, 307, F695–F707. [Google Scholar] [CrossRef]
- Bhave, G.; Colon, S.; Ferrell, N. The sulfilimine cross-link of collagen IV contributes to kidney tubular basement membrane stiffness. Am. J. Physiol. Renal. Physiol. 2017, 313, F596–F602. [Google Scholar] [CrossRef] [Green Version]
- Crest, J.; Diz-Muñoz, A.; Chen, D.-Y.; Fletcher, D.A.; Bilder, D. Organ sculpting by patterned extracellular matrix stiffness. eLife 2017, 6, e24958. [Google Scholar] [CrossRef]
- Hagelaars, M.J.; Yousef Yengej, F.A.; Verhaar, M.C.; Rookmaaker, M.B.; Loerakker, S.; Bouten, C.V.C. Substrate Stiffness Determines the Establishment of Apical-Basal Polarization in Renal Epithelial Cells but Not in Tubuloid-Derived Cells. Front. Bioeng. Biotechnol. 2022, 10, 820930. [Google Scholar] [CrossRef]
- Guimarães, C.F.; Gasperini, L.; Marques, A.P.; Reis, R.L. The stiffness of living tissues and its implications for tissue engineering. Nat. Rev. Mater. 2020, 5, 351–370. [Google Scholar] [CrossRef]
- Beamish, J.A.; Chen, E.; Putnam, A.J. Engineered extracellular matrices with controlled mechanics modulate renal proximal tubular cell epithelialization. PLoS ONE 2017, 12, e0181085. [Google Scholar] [CrossRef] [Green Version]
- Livoti, C.M.; Morgan, J.R. Self-assembly and tissue fusion of toroid-shaped minimal building units. Tissue Eng. Part A 2010, 16, 2051–2061. [Google Scholar] [CrossRef] [Green Version]
- Brown, X.Q.; Ookawa, K.; Wong, J.Y. Evaluation of polydimethylsiloxane scaffolds with physiologically-relevant elastic moduli: Interplay of substrate mechanics and surface chemistry effects on vascular smooth muscle cell response. Biomaterials 2005, 26, 3123–3129. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamon, M.; Chen, Y.; Srivastava, P.; Chang, H.-M.; Gupta, V.; Jin, L.; Yanagawa, N.; Hauser, P.V. Matrix Stiffness Influences Tubular Formation in Renal Tissue Engineering. Appl. Sci. 2023, 13, 4510. https://doi.org/10.3390/app13074510
Hamon M, Chen Y, Srivastava P, Chang H-M, Gupta V, Jin L, Yanagawa N, Hauser PV. Matrix Stiffness Influences Tubular Formation in Renal Tissue Engineering. Applied Sciences. 2023; 13(7):4510. https://doi.org/10.3390/app13074510
Chicago/Turabian StyleHamon, Morgan, Yuzhen Chen, Pratyush Srivastava, Hsiao-Min Chang, Vijay Gupta, Lihua Jin, Norimoto Yanagawa, and Peter V. Hauser. 2023. "Matrix Stiffness Influences Tubular Formation in Renal Tissue Engineering" Applied Sciences 13, no. 7: 4510. https://doi.org/10.3390/app13074510