

The Potency of Graphitic Carbon Nitride (gC3N4) and Bismuth Sulphide Nanoparticles (Bi2S3) in the Management of Foliar Fungal Pathogens of Maize
 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
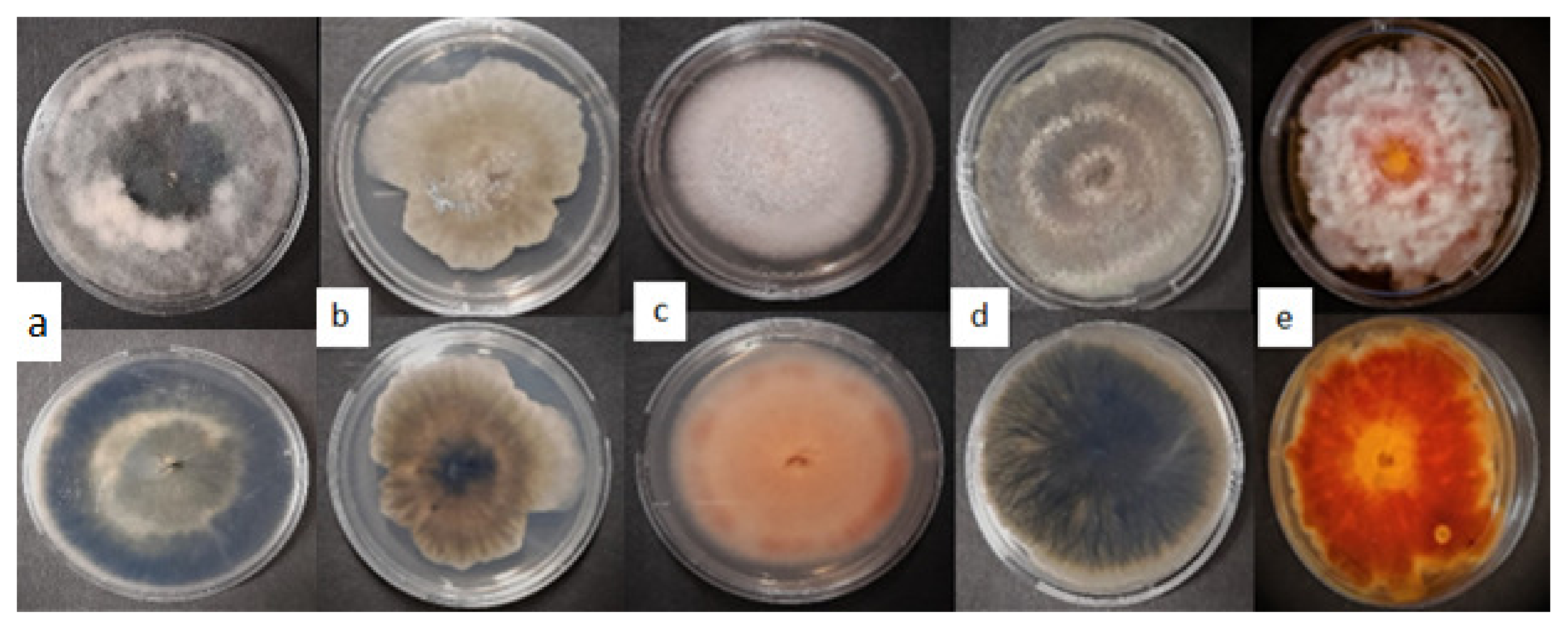
2.1. Sample Collection and Fungal Isolation
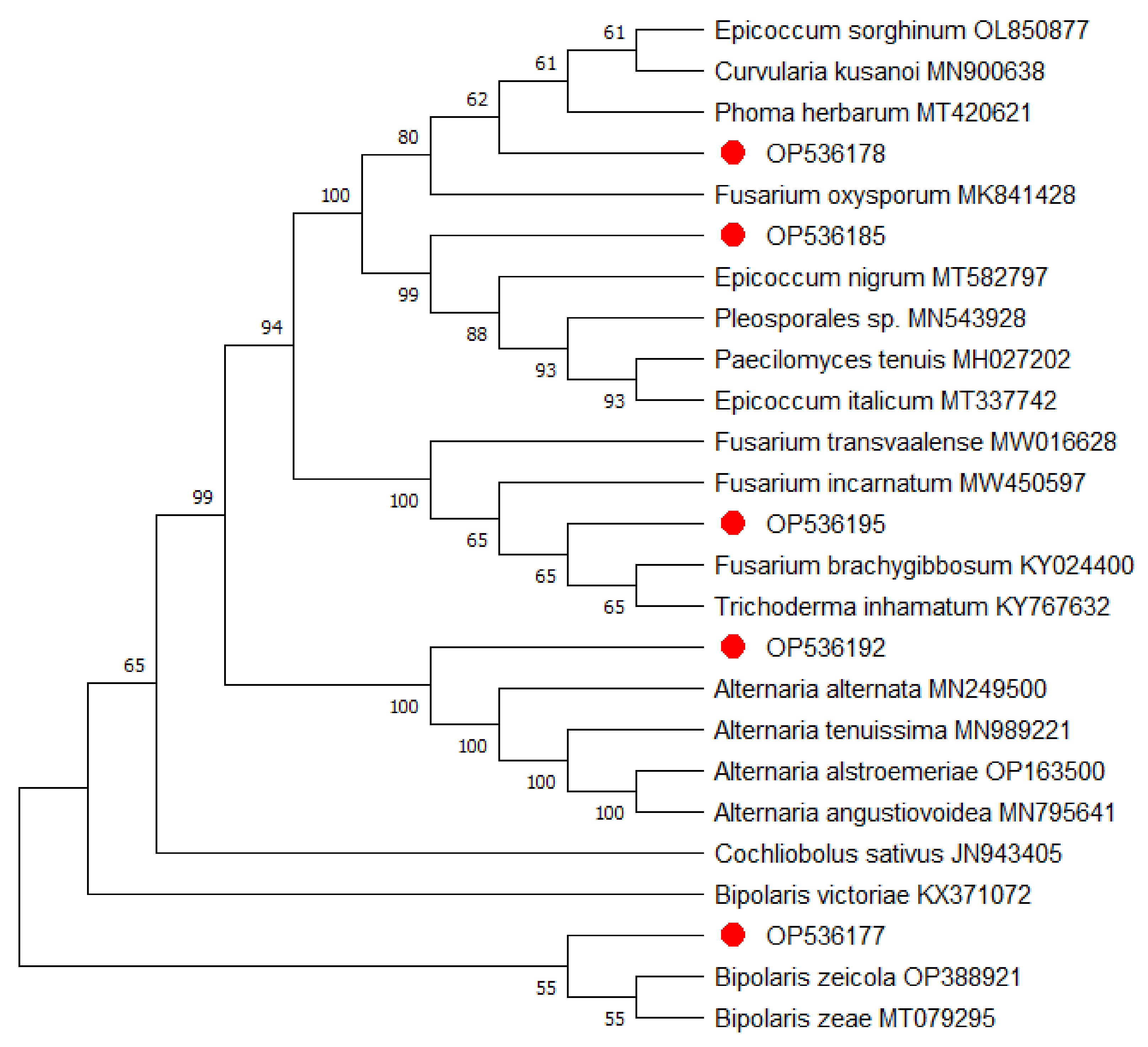
2.2. Molecular Characterization of the Isolated Fungi
2.2.1. Extraction and Amplification of Genomic DNA
2.2.2. Phylogenetic Analysis
3. Synthesis and Characterization of the Nanoparticles
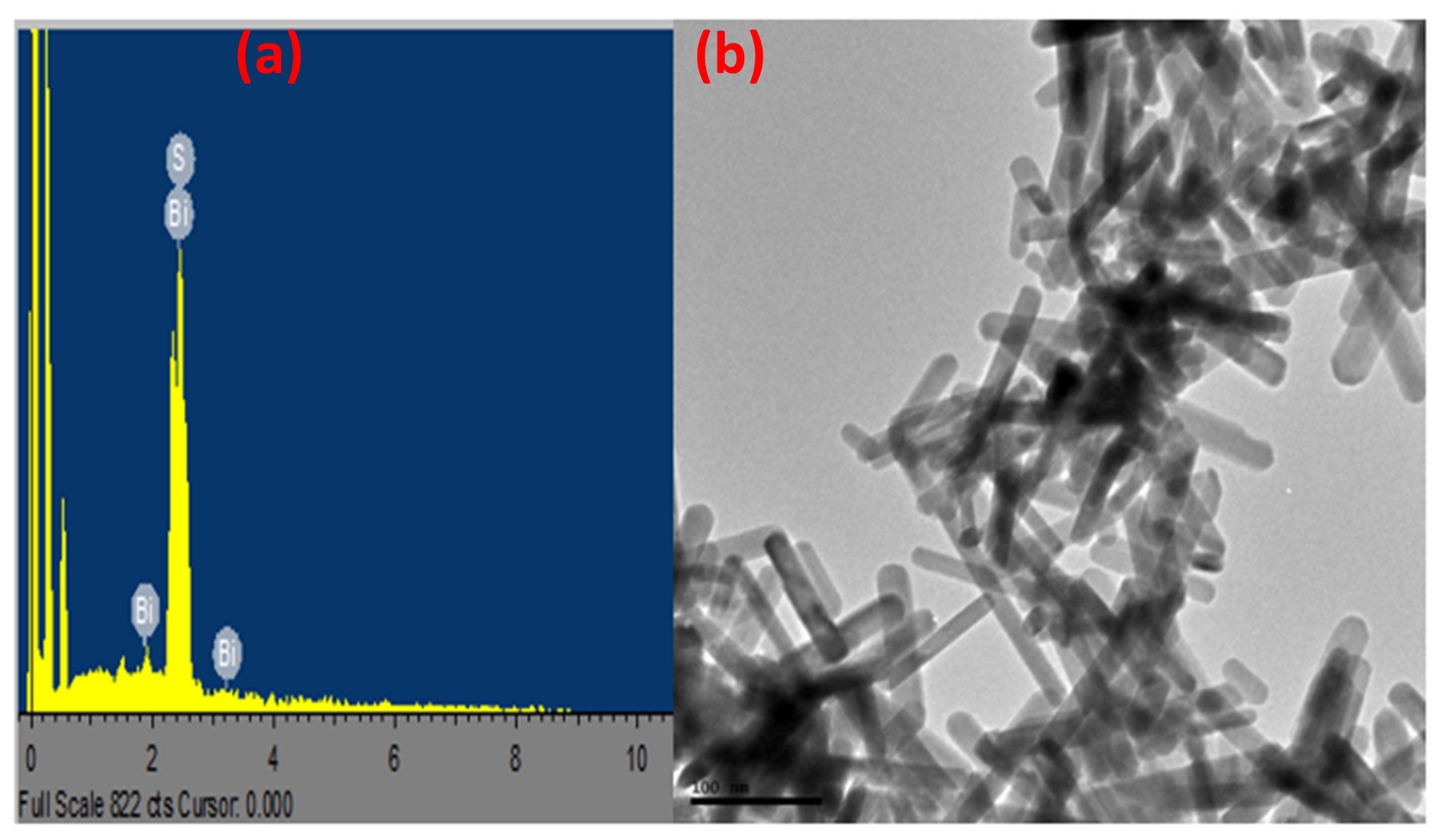
3.1. Synthesis and Characterization of Bismuth Sulphide
3.2. Synthesis of Graphitic Carbon Nitride
4. Nanoparticle Management of Fungi
Statistical Analysis
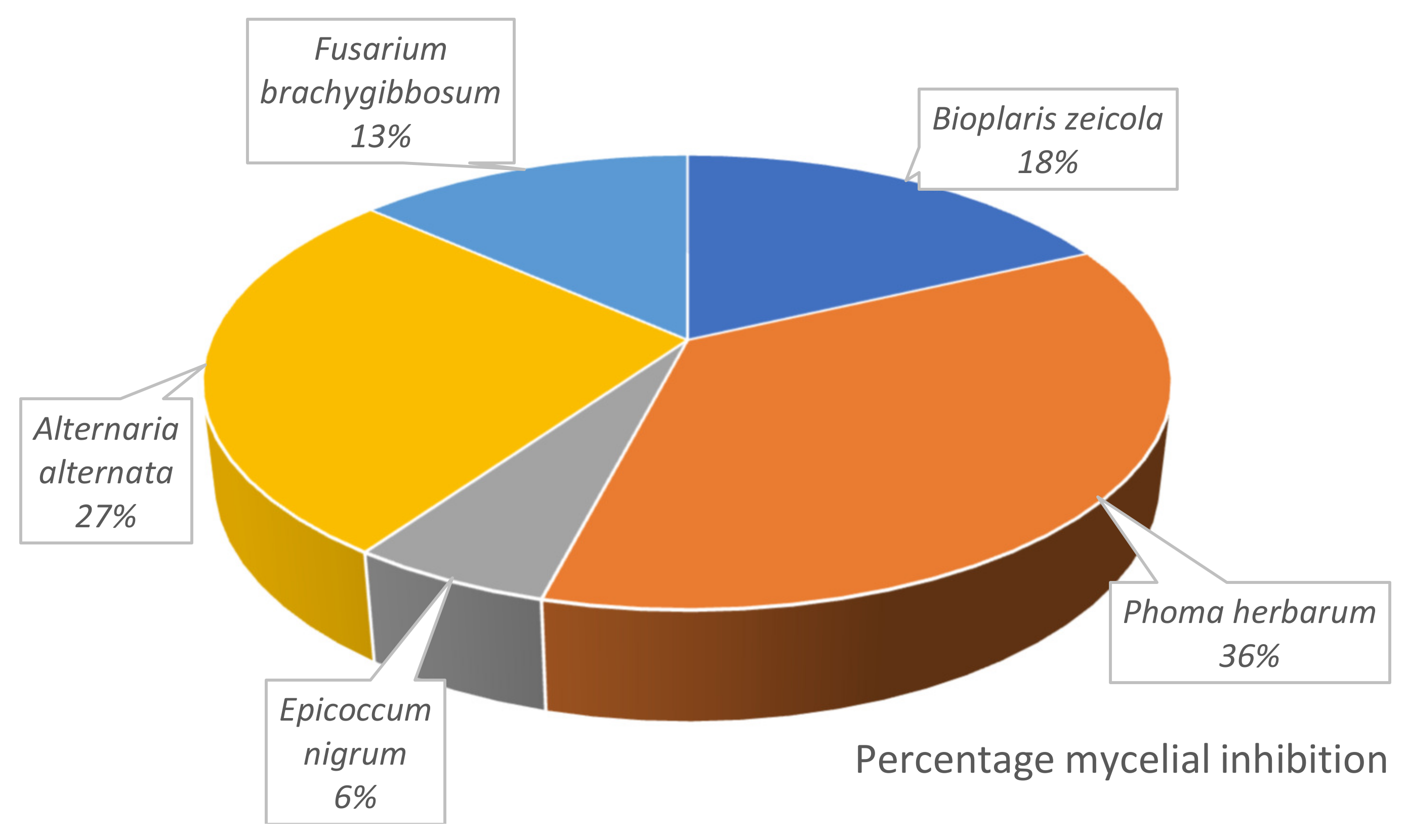
5. Results
6. Discussion
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Bi2S3 | Bismuth sulphide nanoparticles |
| DNA | Deoxyribonucleic acid |
| EDS | Energy-dispersive X-ray spectroscopy |
| gC3N4 | Graphitic carbon nitride nanoparticles |
| ITS | Internal transcribed spacer |
| NaOCl | Sodium hypochlorite |
| NCBI | National Center for Biotechnology Information |
| NH4OH | Ammonium hydroxide solution |
| PCR | Polymerase chain reaction |
| PDA | Potato dextrose agar |
| rRNA | Ribosomal RNA |
| TEM | Transmission electron microscopy |
| XRD | X-ray diffraction |
References
- Tiwari, Y.K.; Yadav, S.K. High temperature stress tolerance in maize (Zea mays L.): Physiological and molecular mechanisms. J. Plant Biol. 2019, 62, 93–102. [Google Scholar] [CrossRef]
- Ngoune, T.L.; Mutengwa, C.S. Estimation of maize (Zea mays L.) yield per harvest area: Appropriate methods. Agronomy 2019, 10, 29. [Google Scholar] [CrossRef] [Green Version]
- Javed, T.; Shabbir, R.; Tahir, A.; Ahmar, S.; Mora-Poblete, F.; Razzaq, M.; Javed, Z.Q.; Zaghum, M.J.; Hussain, S.; Mukhtar, A. Etiology, Epidemiology, and Management of Maize Diseases. In Cereal Diseases: Nanobiotechnological Approaches for Diagnosis and Management; Springer: Berlin/Heidelberg, Germany, 2022; pp. 53–82. [Google Scholar]
- Nji, Q.N.; Babalola, O.O.; Mwanza, M. Aflatoxins in maize: Can their occurrence be effectively managed in Africa in the face of climate change and food insecurity? Toxins 2022, 14, 574. [Google Scholar] [CrossRef] [PubMed]
- Fones, H.N.; Bebber, D.P.; Chaloner, T.M.; Kay, W.T.; Steinberg, G.; Gurr, S.J. Threats to global food security from emerging fungal and oomycete crop pathogens. Nat. Food 2020, 1, 332–342. [Google Scholar] [CrossRef]
- van der Does, H.C.; Rep, M. Virulence genes and the evolution of host specificity in plant-pathogenic fungi. Mol. Plant-Microbe Interact. 2007, 20, 1175–1182. [Google Scholar] [CrossRef] [Green Version]
- Akanmu, A.O.; Sobowale, A.A.; Abiala, M.A.; Olawuyi, O.J.; Odebode, A.C. Efficacy of biochar in the management of Fusarium verticillioides Sacc. causing ear rot in Zea mays L. Biotechnol. Rep. 2020, 26, e00474. [Google Scholar] [CrossRef]
- Belisário, R.; Robertson, A.E.; Vaillancourt, L.J. Maize anthracnose stalk rot in the genomic era. Plant Dis. 2022, 106, 2281–2298. [Google Scholar] [CrossRef]
- Mesterhazy, A.; Szieberth, D.; Toldine, E.T.; Nagy, Z.; Szabó, B.; Herczig, B.; Bors, I.; Tóth, B. Updating the Methodology of Identifying Maize Hybrids Resistant to Ear Rot Pathogens and Their Toxins—Artificial Inoculation Tests for Kernel Resistance to Fusarium graminearum, F. verticillioides, and Aspergillus flavus. J. Fungi 2022, 8, 293. [Google Scholar] [CrossRef]
- Xavier, F.; Kaushik, M. Biological management of charcoal rot of maize caused by Macrophomina phaseolina by using Trichoderma: A. Pharma. Innov. J 2021, 10, 417–441. [Google Scholar]
- ur Rehman, F.; Adnan, M.; Kalsoom, M.; Naz, N.; Husnain, M.G.; Ilahi, H.; Ilyas, M.A.; Yousaf, G.; Tahir, R.; Ahmad, U. Seed-borne fungal diseases of Maize (Zea mays L.): A review. Agrinula J. Agroteknologi Dan Perkeb. 2021, 4, 43–60. [Google Scholar] [CrossRef]
- Ferris, A.C.; Walbot, V. Understanding ustilago maydis infection of multiple maize organs. J. Fungi 2020, 7, 8. [Google Scholar] [CrossRef]
- Akanmu, A.O.; Babalola, O.O.; Venturi, V.; Ayilara, M.S.; Adeleke, B.S.; Amoo, A.E.; Sobowale, A.A.; Fadiji, A.E.; Glick, B.R. Plant Disease Management: Leveraging on the Plant-Microbe-Soil Interface in the Biorational Use of Organic Amendments. Front. Plant Sci. 2021, 12, 1590. [Google Scholar] [CrossRef]
- Sibiya, M.; Sumbwanyambe, M. A computational procedure for the recognition and classification of maize leaf diseases out of healthy leaves using convolutional neural networks. AgriEngineering 2019, 1, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Kotze, R.; Van der Merwe, C.; Crampton, B.; Kritzinger, Q. A histological assessment of the infection strategy of Exserohilum turcicum in maize. Plant Pathol. 2019, 68, 504–512. [Google Scholar] [CrossRef]
- Craven, M.; Morey, L.; Abrahams, A.; Njom, H.A.; van Rensburg, B.J. Effect of northern corn leaf blight severity on Fusarium ear rot incidence of maize. South Afr. J. Sci. 2020, 116, 11–12. [Google Scholar] [CrossRef]
- Latterell, F.M.; Rossi, A.E. Gray leaf spot of corn: A disease on the move. Plant Dis. 1983, 67, 842–847. [Google Scholar] [CrossRef]
- Degani, O.; Gordani, A.; Becher, P.; Chen, A.; Rabinovitz, O. Crop rotation and minimal tillage selectively affect maize growth promotion under late wilt disease stress. J. Fungi 2022, 8, 586. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Ayangbenro, A.S.; Babalola, O.O. Organic farming enhances the diversity and community structure of endophytic archaea and fungi in maize plant: A shotgun approach. J. Soil Sci. Plant Nutr. 2020, 20, 2587–2599. [Google Scholar] [CrossRef]
- Fasusi, O.A.; Cruz, C.; Babalola, O.O. Agricultural sustainability: Microbial biofertilizers in rhizosphere management. Agriculture 2021, 11, 163. [Google Scholar] [CrossRef]
- Gossen, B.D.; McDonald, M.R. New technologies could enhance natural biological control and disease management and reduce reliance on synthetic pesticides. Can. J. Plant Pathol. 2020, 42, 30–40. [Google Scholar] [CrossRef]
- Wise, K.; Mueller, D. Are Fungicides No Longer Just for Fungi? An Analysis of Foliar Fungicide Use in Corn. Available online: https://www.apsnet.org/edcenter/apsnetfeatures/Pages/fungicide.aspx (accessed on 7 March 2023).
- Olowe, O.M.; Nicola, L.; Asemoloye, M.D.; Akanmu, A.O.; Babalola, O.O. Trichoderma: Potential bio-resource for the management of tomato root rot diseases in Africa. Microbiol. Res. 2022, 257, 126978. [Google Scholar] [CrossRef] [PubMed]
- Rani, L.; Thapa, K.; Kanojia, N.; Sharma, N.; Singh, S.; Grewal, A.S.; Srivastav, A.L.; Kaushal, J. An extensive review on the consequences of chemical pesticides on human health and environment. J. Clean. Prod. 2021, 283, 124657. [Google Scholar] [CrossRef]
- Hammami, I.; Alabdallah, N.M. Gold nanoparticles: Synthesis properties and applications. J. King Saud Univ.-Sci. 2021, 33, 101560. [Google Scholar] [CrossRef]
- Ajilogba, C.F.; Babalola, O.O.; Nikoro, D.O. Nanotechnology as vehicle for biocontrol of plant diseases in crop production. In Food Security and Safety; Springer: Cham, Switzerland, 2021; pp. 709–724. [Google Scholar]
- Younas, A.; Yousaf, Z.; Rashid, M.; Riaz, N.; Fiaz, S.; Aftab, A.; Haung, S. Nanotechnology and plant disease diagnosis and management. In Nanoagronomy; Springer: Cham, Switzerland, 2020; pp. 101–123. [Google Scholar]
- Fadiji, A.E.; Mortimer, P.E.; Xu, J.; Ebenso, E.E.; Babalola, O.O. Biosynthesis of nanoparticles using endophytes: A novel approach for enhancing plant growth and sustainable agriculture. Sustainability 2022, 14, 10839. [Google Scholar] [CrossRef]
- Imade, E.E.; Ajiboye, T.O.; Fadiji, A.E.; Onwudiwe, D.C.; Babalola, O.O. Green synthesis of zinc oxide nanoparticles using plantain peel extracts and the evaluation of their antibacterial activity. Sci. Afr. 2022, 16, e01152. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, P.; Li, M.; Shakoor, N.; Adeel, M.; Zhou, P.; Guo, M.; Jiang, Y.; Zhao, W.; Lou, B.; et al. Application and mechanisms of metal-based nanoparticles in the control of bacterial and fungal crop diseases. Pest Manag. Sci. 2023, 79, 21–36. [Google Scholar] [CrossRef]
- Daccò, C.; Nicola, L.; Temporiti, M.E.E.; Mannucci, B.; Corana, F.; Carpani, G.; Tosi, S. Trichoderma: Evaluation of its degrading abilities for the bioremediation of hydrocarbon complex mixtures. Appl. Sci. 2020, 10, 3152. [Google Scholar] [CrossRef]
- Mansoor Ali, S.; Aldawood, S.; AlGarawi, M.S.; AlGhamdi, S.S.; Kassim, H.; Aziz, A. Influence of gamma irradiation on structural, optical, and electrical characterization of Bi2S3 thin films. J. Mater. Sci. Mater. Electron. 2022, 33, 18982–18990. [Google Scholar] [CrossRef]
- Cheng, D.; Wu, H.; Feng, C.; Ding, Y.; Mei, H. Bifunctional photoelectrochemical sensor based on Bi/Bi2S3/BiVO4 for detecting hexavalent chromium and hydrogen peroxide. Sens. Actuators B Chem. 2022, 353, 131108. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L. Synthesis and characterization of Bi2S3 nanorods by solvothermal method in polyol media. Mater. Lett. 2007, 61, 1667–1670. [Google Scholar] [CrossRef]
- Imam, S.S.; Adnan, R.; Mohd Kaus, N.H. Room-temperature synthesis of flower-like BiOBr/Bi2S3 composites for the catalytic degradation of fluoroquinolones using indoor fluorescent light illumination. Colloids Surf. A Physicochem. Eng. Asp. 2020, 585, 124069. [Google Scholar] [CrossRef]
- Ajiboye, T.O.; Kuvarega, A.T.; Onwudiwe, D.C. Graphitic carbon nitride-based catalysts and their applications: A review. Nano-Struct. Nano-Objects 2020, 24, 100577. [Google Scholar] [CrossRef]
- Gashi, A.; Parmentier, J.; Fioux, P.; Marsalek, R. Tuning the C/N Ratio of C-Rich Graphitic Carbon Nitride (g-C3N4) Materials by the Melamine/Carboxylic Acid Adduct Route. Chem.—A Eur. J. 2022, 28, e202103605. [Google Scholar] [CrossRef]
- Ajiboye, T.O.; Oyewo, O.A.; Marzouki, R.; Onwudiwe, D.C. Photocatalytic Reduction of Hexavalent Chromium Using Cu3.21Bi4.79S9/g-C3N4 Nanocomposite. Catalysts 2022, 12, 1075. [Google Scholar] [CrossRef]
- Ajiboye, T.O.; Imade, E.E.; Oyewo, O.A.; Onwudiwe, D.C. Silver functionalized gC3N4: Photocatalytic potency for chromium(VI) reduction, and evaluation of the antioxidant and antimicrobial properties. J. Photochem. Photobiol. A Chem. 2022, 432, 114107. [Google Scholar] [CrossRef]
- Akanmu, A.O.; Odebode, A.C.; Abiala, M.A.; Aiyelaagbe, O.O.; Olaoluwa, O.O. Inhibition of Fusarium pathogens in millet by extracts of Jatropha curcas and Mangifera indica. Int. J. Plant Biol. Res. 2014, 2, 1007. [Google Scholar]
- Olowe, O.M.; Nicola, L.; Aemoloye, M.D.; Akanmu, A.O.; Sobowale, A.A.; Babalola, O.O. Characterization and antagonistic potentials of selected rhizosphere Trichoderma species against some Fusarium species. Front. Microbiol. 2022, 13, 3757. [Google Scholar] [CrossRef]
- Dodd, J. Recent developments in the maize pathogen Bipolaris zeicola Shoemaker. Maydica 1993, 38, 201–204. [Google Scholar]
- Tsukiboshi, T.; Kimigafukuro, T.; Sato, T. Identification of races of Bipolaris zeicola, the causal fungus of Helminthosporium leaf spot on corn in Japan. Jpn. J. Phytopathol. 1987, 53, 647–649. [Google Scholar] [CrossRef]
- Liu, S.; Guo, N.; Ma, H.; Sun, H.; Zheng, X.; Shi, J. First Report of Root Rot Caused by Bipolaris zeicola on Maize in Hebei Province. Plant Dis. 2021, 105, 2247. [Google Scholar] [CrossRef]
- Sun, X.; Qi, X.; Wang, W.; Liu, X.; Zhao, H.; Wu, C.; Chang, X.; Zhang, M.; Chen, H.; Gong, G. Etiology and symptoms of maize leaf spot caused by Bipolaris spp. in Sichuan, China. Pathogens 2020, 9, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleczewski, N.M.; Flory, S.L. Leaf blight disease on the invasive grass Microstegium vimineum caused by a Bipolaris sp. Plant Dis. 2010, 94, 807–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deb, D.; Khan, A.; Dey, N. Phoma diseases: Epidemiology and control. Plant Pathol. 2020, 69, 1203–1217. [Google Scholar] [CrossRef]
- Zhang, Y.; Crous, P.W.; Schoch, C.L.; Hyde, K.D. Pleosporales. Fungal Divers. 2012, 53, 1–221. [Google Scholar] [CrossRef] [Green Version]
- Garibaldi, A.; Gilardi, G.; Matic, S.; Gullino, M. First Report of Phoma herbarum Causing Leaf Spot of Purple Coneflower (Echinacea purpurea) in Northern Italy. Plant Dis. 2019, 103, 1786. [Google Scholar] [CrossRef]
- Higgins, K.L.; Arnold, A.E.; Coley, P.D.; Kursar, T.A. Communities of fungal endophytes in tropical forest grasses: Highly diverse host-and habitat generalists characterized by strong spatial structure. Fungal Ecol. 2014, 8, 1–11. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, L.; Yang, X.; Cao, H.; Li, J.; Cao, P.; Guo, L.; Wang, X.; Zhao, J.; Xiang, W. Alternaria spp. associated with leaf blight of maize in Heilongjiang Province, China. Plant Dis. 2022, 106, 572–584. [Google Scholar] [CrossRef]
- Kgatle, M.G.; Truter, M.; Ramusi, T.; Flett, B.; Aveling, T. Alternaria alternata, the causal agent of leaf blight of sunflower in South Africa. Eur. J. Plant Pathol. 2018, 151, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Rna, A.; Babikar, A.; Dagash, Y.; Elhussein, A.; Elhalim, T.; Babikar, A. Fusarium Brachygibbosum a Plausible Candidate for Deployment as a Bioagent for Striga Hermonthica Management in Sorghum. In Proceedings of the Third Conference of Pests Management in Sudan February, Wad Medani, Sudan, 3–4 February 2014; pp. 3–4. [Google Scholar]
- Shan, L.; Cui, W.; Zhang, D.; Zhang, J.; Ma, N.; Bao, Y.; Dai, X.; Guo, W. First report of Fusarium brachygibbosum causing maize stalk rot in China. Plant Dis. 2017, 101, 837. [Google Scholar] [CrossRef]
- Taguiam, J.D.; Evallo, E.; Balendres, M.A. Epicoccum species: Ubiquitous plant pathogens and effective biological control agents. Eur. J. Plant Pathol. 2021, 159, 713–725. [Google Scholar] [CrossRef]
- Bagy, H.M.K.; Hassan, E.A.; Nafady, N.A.; Dawood, M.F. Efficacy of arbuscular mycorrhizal fungi and endophytic strain Epicoccum nigrum ASU11 as biocontrol agents against blackleg disease of potato caused by bacterial strain Pectobacterium carotovora subsp. atrosepticum PHY7. Biol. Control 2019, 134, 103–113. [Google Scholar] [CrossRef]
- Abdel-Moniem, S.M.; El-Liethy, M.A.; Ibrahim, H.S.; Ali, M.E. Innovative green/non-toxic Bi2S3@ g-C3N4 nanosheets for dark antimicrobial activity and photocatalytic depollution: Turnover assessment. Ecotoxicol. Environ. Saf. 2021, 226, 112808. [Google Scholar] [CrossRef]
- Soliman, G.M. Nanoparticles as safe and effective delivery systems of antifungal agents: Achievements and challenges. Int. J. Pharm. 2017, 523, 15–32. [Google Scholar] [CrossRef]
- Hamayun, M.; Khan, S.A.; Khan, A.L.; Rehman, G.; Sohn, E.-Y.; Shah, A.A.; Kim, S.-K.; Joo, G.-J.; Lee, I.-J. Phoma herbarum as a new gibberellin-producing and plant growth-promoting fungus. J. Microbiol. Biotechnol. 2009, 19, 1244–1249. [Google Scholar]
- Kim, S.W.; Jung, J.H.; Lamsal, K.; Kim, Y.S.; Min, J.S.; Lee, Y.S. Antifungal effects of silver nanoparticles (AgNPs) against various plant pathogenic fungi. Mycobiology 2012, 40, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Rai, M.; Ingle, A.P.; Paralikar, P. Sulfur and sulfur nanoparticles as potential antimicrobials: From traditional medicine to nanomedicine. Expert Rev. Anti-Infect. Ther. 2016, 14, 969–978. [Google Scholar] [CrossRef]
- Rajiv, P.; Mengelizadeh, N.; McKay, G.; Balarak, D. Photocatalytic degradation of ciprofloxacin with Fe2O3 nanoparticles loaded on graphitic carbon nitride: Mineralisation, degradation mechanism and toxicity assessment. Int. J. Environ. Anal. Chem. 2021, 1–15. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
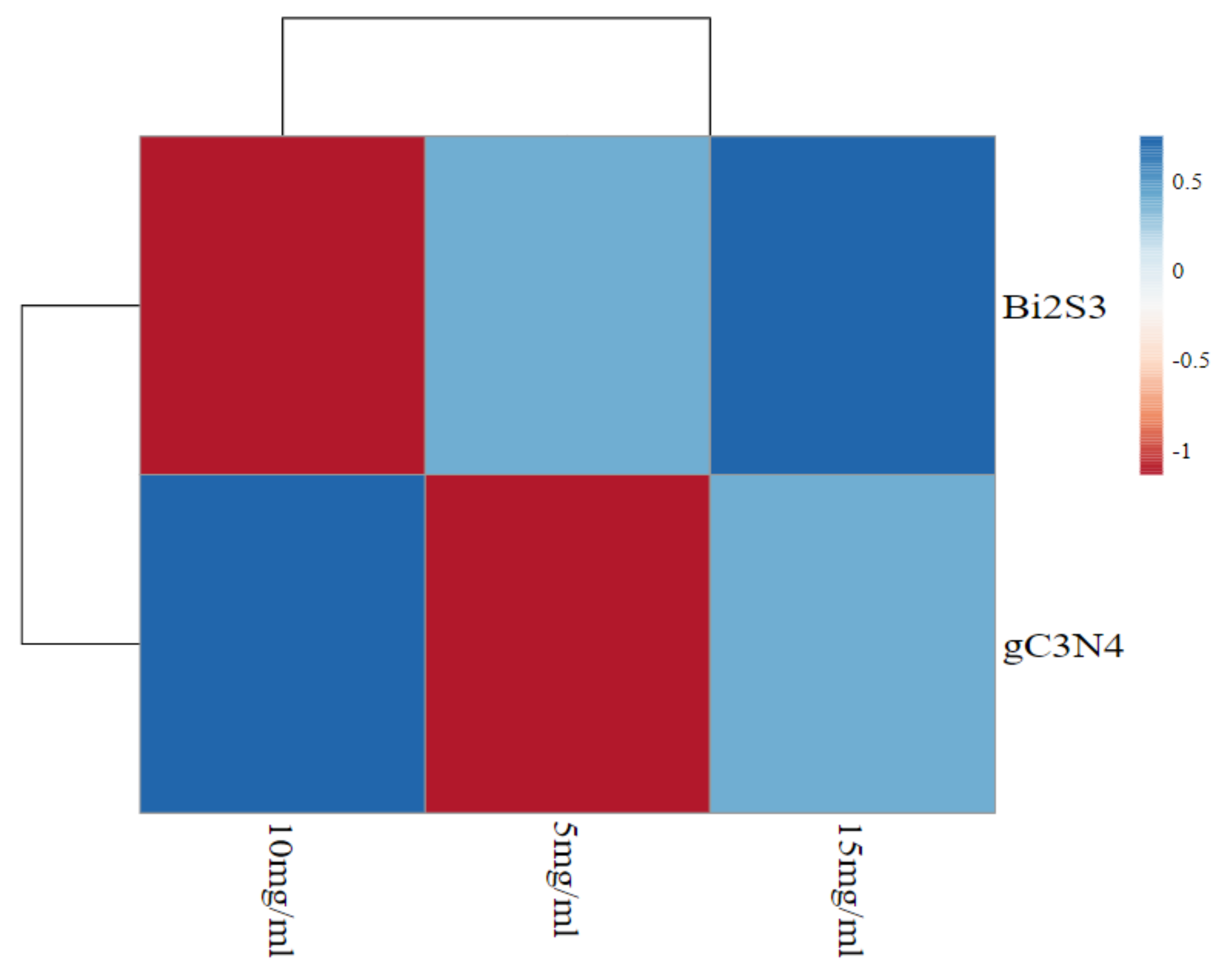{kind=link}
{kind=link}
| Accession Code | Isolate Name | Accession Number | Blastn, Closest Relative | Relative Accession | Similarity % | E Value |
|---|---|---|---|---|---|---|
| AD1 | Bipolaris zeicola strain 1 | OP536177 | Bipolaris zeicola | MK841439 | 99.81 | 0.0 |
| AD6 | Phoma herbarum strain 1 | OP536178 | Phoma herbarum | MT420621 | 100 | 0.0 |
| AH1 | Epicoccum nigrumisolate | OP536185 | Epicoccum nigrum | MT582797 | 100 | 0.0 |
| BD5A | Alternaria alternata strain 2 | OP536192 | Alternaria alternata | MN249500 | 100 | 0.0 |
| BD2 | Fusarium brachygibbosum | OP536195 | Fusarium brachygibbosum | KY024400 | 100 | 0.0 |
| Organisms | Bi2S3 | gC3N4 | ||||
|---|---|---|---|---|---|---|
| 5 mg/mL | 10 mg/mL | 15 mg/mL | 5 mg/mL | 10 mg/mL | 15 mg/mL | |
| Bioplaris zeicola | 31.58 ± 3.30 a | 28.49 ± 2.53 b | 16.97 ± 2.63 b | 13.23 ± 2.80 c | 15.44 ± 2.81 b | 18.53 ± 4.12 b |
| Phoma herbarum | 35.72 ± 4.03 a | 21.09 ± 2.95 c | 39.71 ± 2.90 a | 45.00 ± 3.97 a | 65.64 ± 2.32 a | 41.03 ± 3.21 a |
| Epicoccum nigrum | 4.59 ± 1.21 c | 7.58 ± 1.61 e | 7.54 ± 1.71 c | 5.27 ± 1.37 d | 5.09 ± 1.50 c | 9.06 ± 1.34 c |
| Alternaria alternata | 28.86 ± 5.35 a | 37.56 ± 1.82 a | 35.35 ± 1.87 a | 28.63 ± 2.12 b | 20.42 ± 3.17 b | 34.33 ± 1.79 a |
| Fusarium brachygibbosum | 14.00 ± 2.54 b | 14.13 ± 2.78 d | 16.54 ± 2.92 b | 7.92 ± 2.90 dc | 20.58 ± 2.54 b | 19.37 ± 2.43 b |
| LSD | 7.96 | 6.31 | 5.43 | 6.13 | 6.85 | 7.47 |
| Organisms | 5 mg/mL | 10 mg/mL | 15 mg/mL |
|---|---|---|---|
| Bioplaris zeicola | 22.40 ± 2.53 c | 21.96 ± 2.10 c | 17.75 ± 2.42 c |
| Phoma herbarum | 40.36 ± 2.88 a | 43.37 ± 3.74 a | 40.37 ± 2.14 a |
| Epicoccum nigrum | 4.93 ± 0.90 e | 6.33 ± 1.10 d | 8.30 ± 1.08 d |
| Alternaria alternata | 28.74 ± 2.85 b | 28.99 ± 2.20 b | 34.84 ± 1.28 b |
| Fusarium brachygibbosum | 10.96 ± 1.96 d | 17.35 ± 1.92 c | 17.96 ± 1.89 c |
| LSD | 5.27 | 6.44 | 4.61 |
| Days | Bi2S3 | gC3N4 | ||||
|---|---|---|---|---|---|---|
| 5 mg/mL | 10 mg/mL | 15 mg/mL | 5 mg/mL | 10 mg/mL | 15 mg/mL | |
| 3 | 30.41 ± 4.21 a | 29.32 ± 2.52 a | 31.97 ± 2.98 a | 26.97 ± 3.81 a | 31.71 ± 4.90 a | 30.27 ±3.21 a |
| 5 | 22.80 ± 3.24 b | 21.34 ± 2.35 b | 25.10 ± 2.66 b | 21.82 ± 3.59 ba | 26.27 ± 4.63 ba | 25.82 ± 2.86 ba |
| 7 | 19.65 ± 3.91 b | 19.07 ± 3.13 b | 20.28 ± 3.17 cb | 16.40 ± 3.62 bc | 21.41 ± 3.91 b | 22.88 ± 3.23 b |
| 9 | 18.94 ± 3.61 b | 17.35 ± 2.99 b | 15.53 ± 2.96 c | 14.85 ± 3.35 c | 22.35 ± 4.17 b | 18.88 ± 3.38 b |
| LSD | 7.12 | 5.65 | 4.85 | 5.48 | 6.13 | 6.68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akanmu, A.O.; Ajiboye, T.O.; Seleke, M.; Mhlanga, S.D.; Onwudiwe, D.C.; Babalola, O.O. The Potency of Graphitic Carbon Nitride (gC3N4) and Bismuth Sulphide Nanoparticles (Bi2S3) in the Management of Foliar Fungal Pathogens of Maize. Appl. Sci. 2023, 13, 3731. https://doi.org/10.3390/app13063731
Akanmu AO, Ajiboye TO, Seleke M, Mhlanga SD, Onwudiwe DC, Babalola OO. The Potency of Graphitic Carbon Nitride (gC3N4) and Bismuth Sulphide Nanoparticles (Bi2S3) in the Management of Foliar Fungal Pathogens of Maize. Applied Sciences. 2023; 13(6):3731. https://doi.org/10.3390/app13063731
Chicago/Turabian StyleAkanmu, Akinlolu Olalekan, Timothy Oladiran Ajiboye, Masego Seleke, Sabelo D. Mhlanga, Damian C. Onwudiwe, and Olubukola Oluranti Babalola. 2023. "The Potency of Graphitic Carbon Nitride (gC3N4) and Bismuth Sulphide Nanoparticles (Bi2S3) in the Management of Foliar Fungal Pathogens of Maize" Applied Sciences 13, no. 6: 3731. https://doi.org/10.3390/app13063731