

Comparative Analysis of the Effect of Different Concentrations of Dextran Sodium Sulfate on the Severity and Extent of Inflammation in Experimental Ulcerative Colitis
 , , and
, , and 
Abstract
:1. Introduction
- Organoid culture: This involves the growth of colonic crypts in vitro to generate 3D structures that mimic the native tissue architecture [11].
- Cell line models: These models use immortalised cell lines such as Caco-2 or HT-29 cells, which are commonly used to study epithelial cell biology and inflammation [12].
- Co-culture models: These models use combinations of different cell types, such as epithelial cells and immune cells, to study interactions and inflammation in a more complex environment [13].
- Microfluidic models: These models utilise microfluidic devices to create controlled microenvironments for the study of cellular responses in a controlled and reproducible manner [14].
2. Material and Methods
2.1. Animals and Dextran Sodium Sulfate
2.2. Establishment of a Colitis Model
2.3. Dextran Sodium Sulfate Model of Colitis
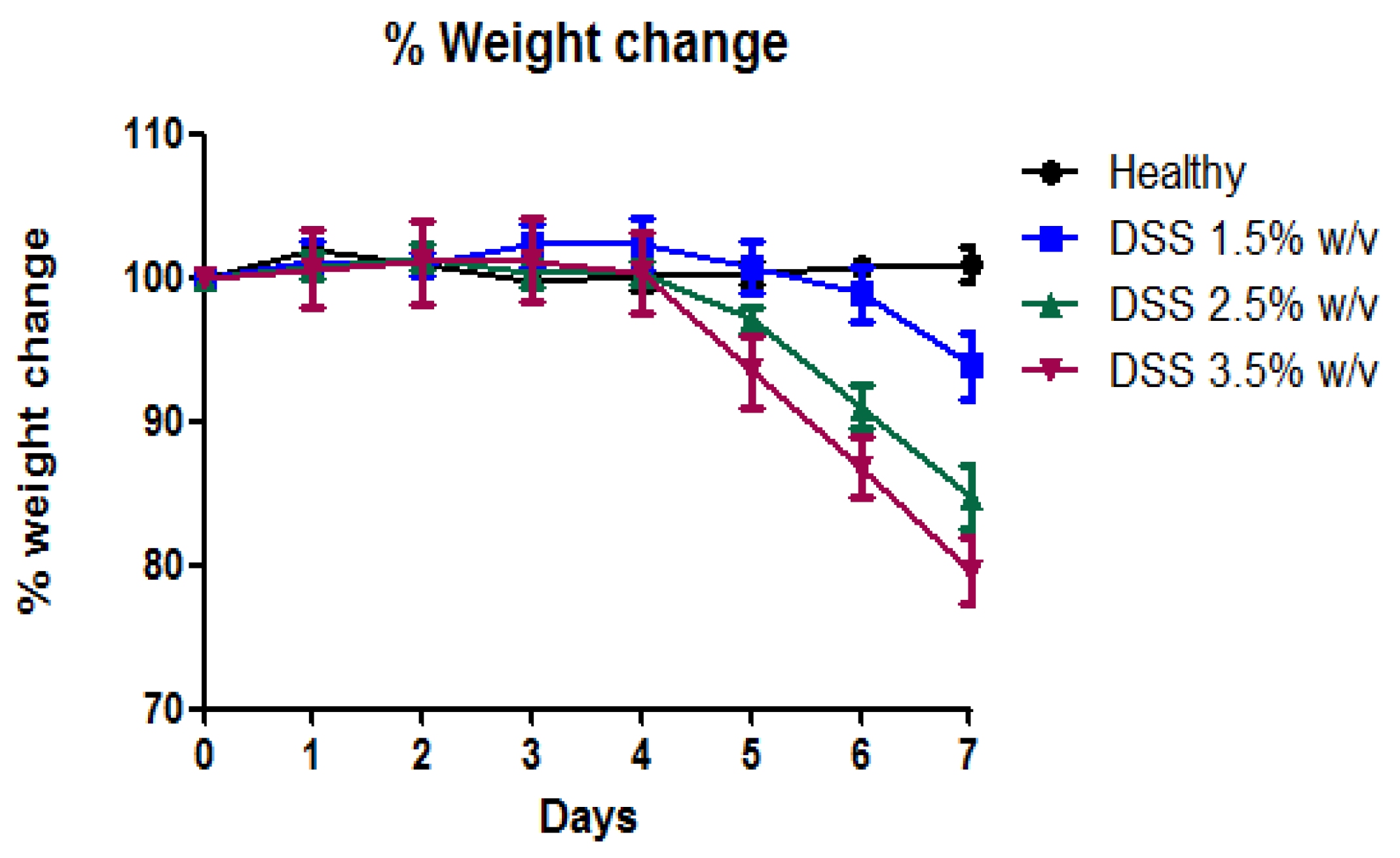
2.4. Percentage Weight Change
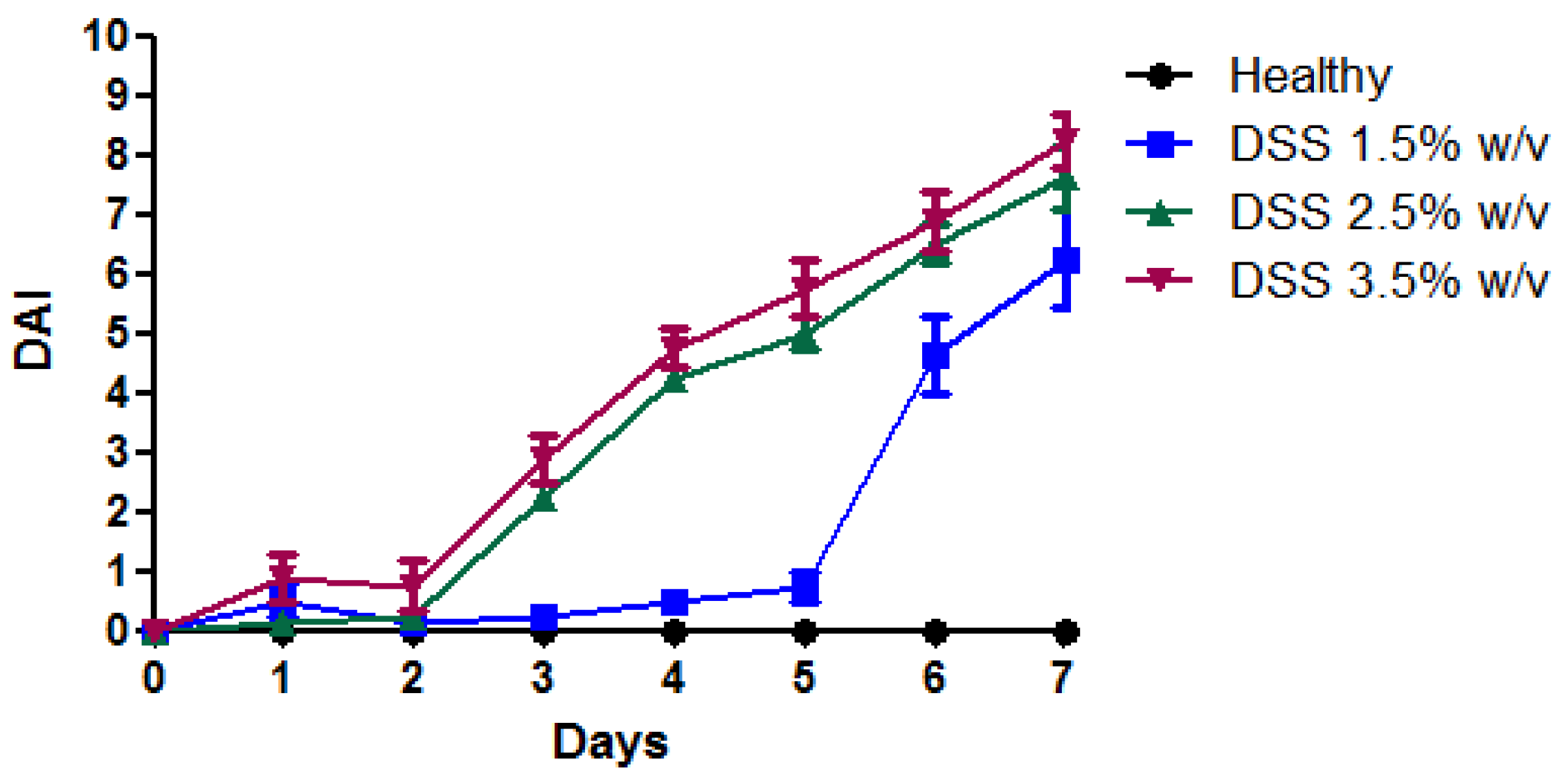
2.5. Disease Activity Index (DAI)
2.6. Change in Colon Length
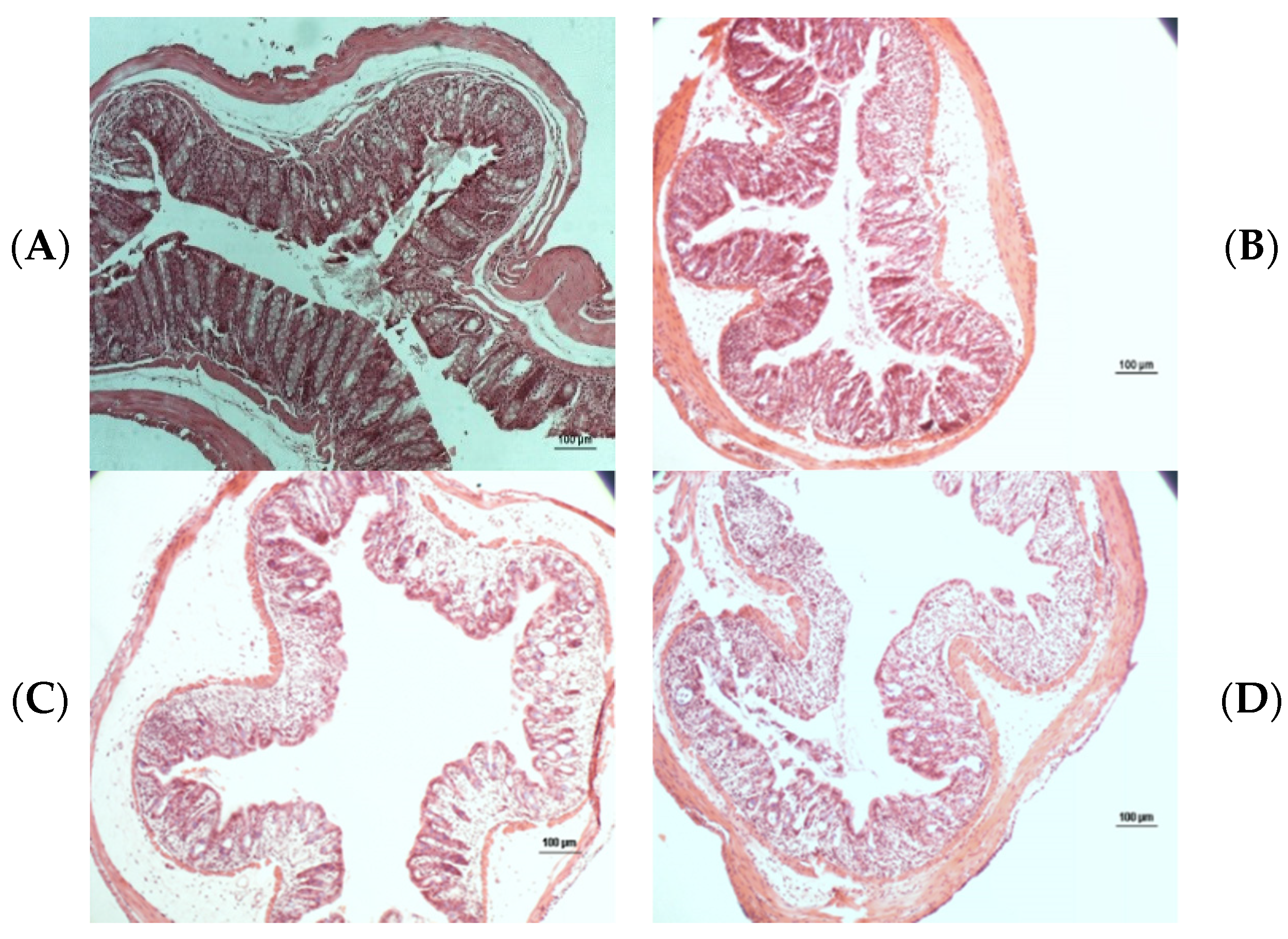
2.7. Histopathological Analysis
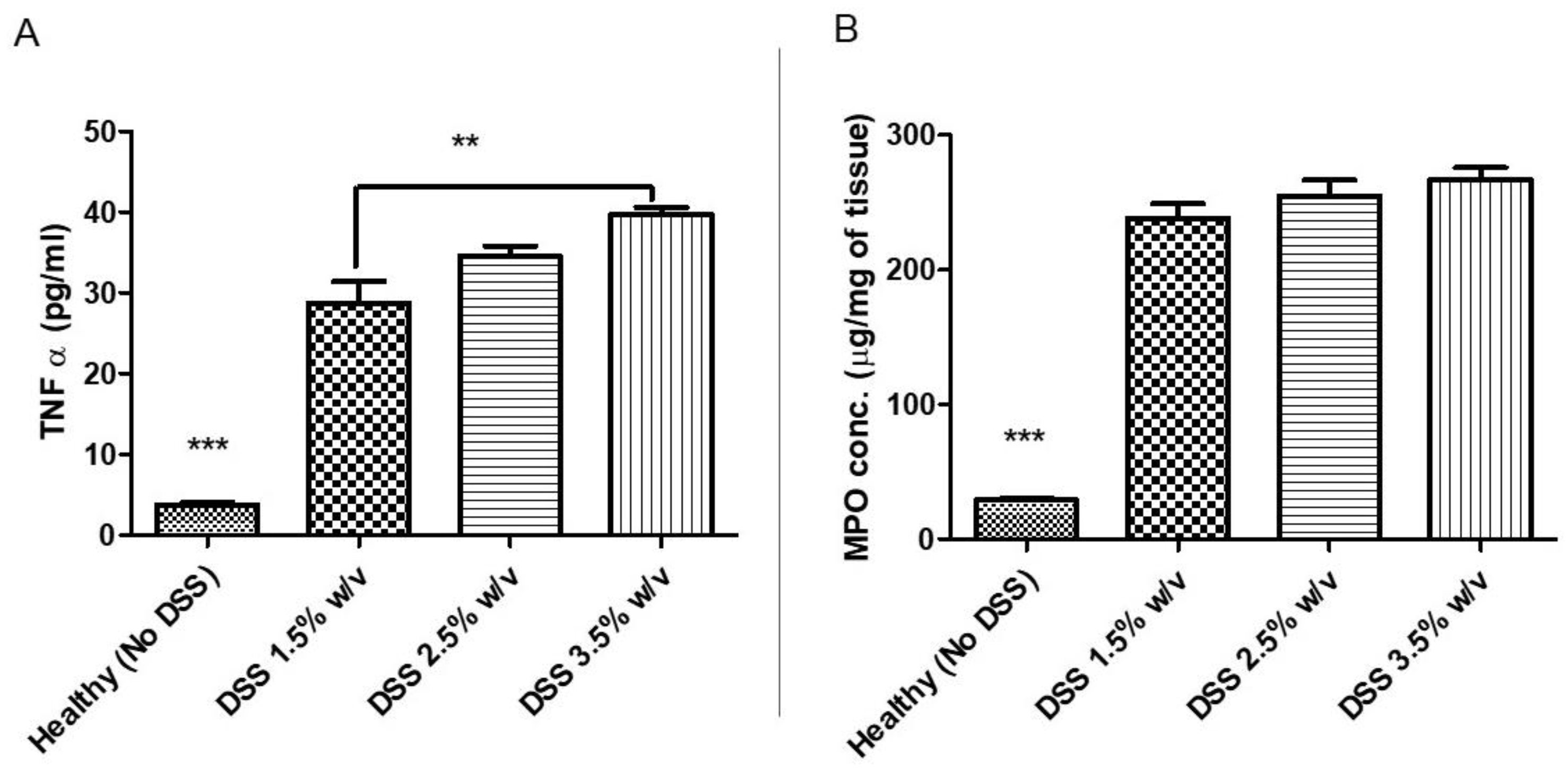
2.8. Colon Cytokine and Myeloperoxidases Measurements
3. Result and Discussion
4. Discussion
5. Conclusions
6. Future Work
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feagan, B.G.; Rutgeerts, P.; Sands, B.E.; Hanauer, S.; Colombel, J.-F.; Sandborn, W.J.; Van Assche, G.; Axler, J.; Kim, H.-J.; Danese, S.; et al. Vedolizumab as Induction and Maintenance Therapy for Ulcerative Colitis. N. Engl. J. Med. 2013, 369, 699–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizoguchi, E.; Nguyen, D.; Low, D. Animal models of ulcerative colitis and their application in drug research. Drug Des. Dev. Ther. 2013, 7, 1341–1357. [Google Scholar] [CrossRef] [Green Version]
- Akolkar, P.N.; Gulwani-Akolkar, B.; Heresbach, D.; Lin, X.Y.; Fisher, S.; Katz, S.; Silver, J. Differences in risk of Crohn’s disease in offspring of mothers and fathers with inflammatory bowel disease. Am. J. Gastroenterol. 1997, 92, 2241–2244. [Google Scholar]
- Anderson, C.A.; Boucher, G.; Lees, C.W.; Franke, A.; D’Amato, M.; Taylor, K.D.; Lee, J.C.; Goyette, P.; Imielinski, M.; Latiano, A.; et al. Meta-analysis identifies 29 additional ulcerative colitis risk loci, increasing the number of confirmed associations to 47. Nat. Genet. 2011, 43, 246–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahid, S.S.; Minor, K.S.; Soto, R.E.; Hornung, C.A.; Galandiuk, S. Smoking and Inflammatory Bowel Disease: A Meta-analysis. Mayo Clin. Proc. 2006, 81, 1462–1471. [Google Scholar] [CrossRef] [PubMed]
- Strober, W.; Fuss, I. Pro-Inflammatory Cytokines in the Pathogenesis of IBD. Gastroenterology 2011, 140, 1756–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, B.B.; Harikumar, K.B. Potential therapeutic effects of curcumin, the anti-inflammatory agent, against neurodegenerative, cardiovascular, pulmonary, metabolic, autoimmune and neoplastic diseases. Int. J. Biochem. Cell Biol. 2009, 41, 40–59. [Google Scholar] [CrossRef] [Green Version]
- Neurath, M.F. New targets for mucosal healing and therapy in inflammatory bowel diseases. Mucosal Immunol. 2014, 7, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Qin, X. Why is damage limited to the mucosa in ulcerative colitis but transmural in Crohn’s disease? World J. Gastrointest. Pathophysiol. 2013, 4, 63–64. [Google Scholar] [CrossRef]
- Chen, H.; Fang, Y.; Li, W.; Orlando, R.C.; Shaheen, N.; Chen, X.L. NFkB and Nrf2 in esophageal epithelial barrier function. Tissue Barriers 2013, 1, e27463. [Google Scholar] [CrossRef] [Green Version]
- D’Aldebert, E.; Quaranta, M.; Sébert, M.; Bonnet, D.; Kirzin, S.; Portier, G.; Duffas, J.-P.; Chabot, S.; Lluel, P.; Allart, S.; et al. Characterization of Human Colon Organoids From Inflammatory Bowel Disease Patients. Front. Cell Dev. Biol. 2020, 8, 363. [Google Scholar] [CrossRef]
- Dieleman, L.A.; Palmen, M.J.H.J.; Akol, H.; Bloemena, E.; Peña, A.S.; Meuwissen, S.G.M.; Van Rees, E.P. Chronic experimental colitis induced by dextran sulphate sodium (DSS) is characterized by Th1 and Th2 cytokines. Clin. Exp. Immunol. 1998, 114, 385–391. [Google Scholar] [CrossRef]
- Kämpfer, A.A.; Urbán, P.; Gioria, S.; Kanase, N.; Stone, V.; Kinsner-Ovaskainen, A. Development of an in vitro co-culture model to mimic the human intestine in healthy and diseased state. Toxicol. Vitr. 2017, 45, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wen, H.; Li, S.; Cao, H.; Li, X.; Ma, Z.; She, X.; Zhou, L.; Huang, S. Emerging microfluidic technologies for microbiome research. Front. Microbiol. 2022, 13, 3116, Erratum in: Front Microbiol. 2022, 13, 1038682. [Google Scholar] [CrossRef] [PubMed]
- Roh, T.T.; Chen, Y.; Paul, H.T.; Guo, C.; Kaplan, D.L. 3D bioengineered tissue model of the large intestine to study inflammatory bowel disease. Biomaterials 2019, 225, 119517. [Google Scholar] [CrossRef] [PubMed]
- Hendow, E.K.; Guhmann, P.; Wright, B.; Sofokleous, P.; Parmar, N.; Day, R.M. Biomaterials for hollow organ tissue engineering. Fibrogenesis Tissue Repair 2016, 9, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, A.; Soni, A.; Acharya, S. In vitro models and ex vivo systems used in inflammatory bowel disease. Vitr. Model. 2022, 1, 213–227. [Google Scholar] [CrossRef]
- Khan, R.; Roy, N.; Ali, H.; Naeem, M. Fecal Microbiota Transplants for Inflammatory Bowel Disease Treatment: Synthetic- and Engineered Communities-Based Microbiota Transplants Are the Future. Gastroenterol. Res. Pract. 2022, 2022, 9999925. [Google Scholar] [CrossRef]
- Hartman, K.G.; Bortner, J.J.D.; Falk, G.; Ginsberg, G.G.; Jhala, N.; Yu, J.; Martín, M.G.; Rustgi, A.K.; Lynch, J.P. Modeling human gastrointestinal inflammatory diseases using microphysiological culture systems. Exp. Biol. Med. 2014, 239, 1108–1123. [Google Scholar] [CrossRef] [Green Version]
- Randhawa, P.K.; Singh, K.; Singh, N.; Jaggi, A.S. A Review on Chemical-Induced Inflammatory Bowel Disease Models in Rodents. Korean J. Physiol. Pharmacol. 2014, 18, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran Sulfate Sodium (DSS)-Induced Colitis in Mice. Curr. Protoc. Immunol. 2014, 104, 15.25.1–15.25.14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, B.; Bajaj-Elliott, M.; Macdonald, T.T.; Inglin, R.; Eysselein, V.E.; Büchler, M.W. Characterisation of Acute Murine Dextran Sodium Sulphate Colitis: Cytokine Profile and Dose Dependency. Digestion 2000, 62, 240–248. [Google Scholar] [CrossRef]
- Okayasu, I.; Hatakeyama, S.; Yamada, M.; Ohkusa, T.; Inagaki, Y.; Nakaya, R. A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 1990, 98, 694–702. [Google Scholar] [CrossRef]
- Friedman, D.J.; Künzli, B.M.; A-Rahim, Y.I.; Sevigny, J.; Berberat, P.O.; Enjyoji, K.; Csizmadia, E.; Friess, H.; Robson, S.C. From the cover: CD39 deletion exacerbates experimental murine colitis and human polymorphisms increase susceptibility to inflammatory bowel disease. Proc. Natl. Acad. Sci. USA 2009, 106, 16788–16793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Si, J.-M.; Liu, W.-L.; Cai, J.-T.; Du, Q.; Wang, L.-J.; Gao, M. Induction of experimental acute ulcerative colitis in rats by administration of dextran sulfate sodium at low concentration followed by intracolonic administration of 30% ethanol. J. Zhejiang Univ. B 2007, 8, 632–637. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, A.; Andoh, A.; Araki, Y.; Bamba, T.; Fujiyama, Y. Neutralization of interleukin-17 aggravates dextran sulfate sodium-induced colitis in mice. Clin. Immunol. 2003, 110, 55–62. [Google Scholar] [CrossRef]
- Tambuwala, M.M.; Manresa, M.C.; Cummins, E.P.; Aversa, V.; Coulter, I.S.; Taylor, C.T. Targeted delivery of the hydroxylase inhibitor DMOG provides enhanced efficacy with reduced systemic exposure in a murine model of colitis. J. Control. Release 2015, 217, 221–227. [Google Scholar] [CrossRef]
- Boirivant, M.; Cossu, A. Inflammatory bowel disease. Oral Dis. 2011, 18, 1–15. [Google Scholar] [CrossRef]
- Kim, J.; Shajib, S.; Manocha, M.M.; Khan, W.I. Investigating Intestinal Inflammation in DSS-induced Model of IBD. J. Vis. Exp. 2012, e3678. [Google Scholar] [CrossRef] [Green Version]
- Taghipour, N.; Molaei, M.; Mosaffa, N.; Rostami-Nejad, M.; Agdaei, H.A.; Anissian, A.; Azimzadeh, P.; Zali, M.R. An experimental model of colitis induced by dextran sulfate sodium from acute progresses to chronicity in C57BL/6: Correlation between conditions of mice and the environment. Gastroenterol. Hepatol. Bed Bench 2015, 1, 45–52. [Google Scholar] [CrossRef]
- Perše, M.; Cerar, A. Dextran Sodium Sulphate Colitis Mouse Model: Traps and Tricks. J. Biomed. Biotechnol. 2012, 2012, 718617. [Google Scholar] [CrossRef] [Green Version]
- Neu, J.; Walker, W.A. Necrotizing Enterocolitis. N. Engl. J. Med. 2011, 364, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Hermiston, M.L.; Gordon, J.I. Inflammatory Bowel Disease and Adenomas in Mice Expressing a Dominant Negative N-Cadherin. Science 1995, 270, 1203–1207. [Google Scholar] [CrossRef]
- Krieglstein, C.F.; Cerwinka, W.H.; Sprague, A.G.; Laroux, F.S.; Grisham, M.B.; Koteliansky, V.E.; Senninger, N.; Granger, D.N.; De Fougerolles, A.R. Collagen-binding integrin α1β1 regulates intestinal inflammation in experimental colitis. J. Clin. Investig. 2002, 110, 1773–1782. [Google Scholar] [CrossRef] [Green Version]
- Weng, Y.-J.; Jiang, D.-X.; Liang, J.; Ye, S.-C.; Tan, W.-K.; Yu, C.-Y.; Zhou, Y. Effects of Pretreatment with Bifidobacterium bifidum Using 16S Ribosomal RNA Gene Sequencing in a Mouse Model of Acute Colitis Induced by Dextran Sulfate Sodium. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2021, 27, e928478-1–e928478-12. [Google Scholar] [CrossRef]
- Lobionda, S.; Sittipo, P.; Kwon, H.Y.; Lee, Y.K. The Role of Gut Microbiota in Intestinal Inflammation with Respect to Diet and Extrinsic Stressors. Microorganisms 2019, 7, 271. [Google Scholar] [CrossRef] [Green Version]
- Dieleman, L.A.; Ridwan, B.U.; Tennyson, G.S.; Beagley, K.W.; Bucy, R.; Elson, C.O. Dextran sulfate sodium-induced colitis occurs in severe combined immunodeficient mice. Gastroenterology 1994, 107, 1643–1652. [Google Scholar] [CrossRef]
- Fukata, M.; Michelsen, K.S.; Eri, R.; Thomas, L.S.; Hu, B.; Lukasek, K.; Nast, C.C.; Lechago, J.; Xu, R.; Naiki, Y.; et al. Toll-like receptor-4 is required for intestinal response to epithelial injury and limiting bacterial translocation in a murine model of acute colitis. Am. J. Physiol. Liver Physiol. 2005, 288, G1055–G1065. [Google Scholar] [CrossRef] [Green Version]
- Pull, S.L.; Doherty, J.M.; Mills, J.C.; Gordon, J.I.; Stappenbeck, T.S. Activated macrophages are an adaptive element of the colonic epithelial progenitor niche necessary for regenerative responses to injury. Proc. Natl. Acad. Sci. USA 2004, 102, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Chami, B.; Yeung, A.W.S.; van Vreden, C.; King, N.J.C.; Bao, S. The Role of CXCR3 in DSS-Induced Colitis. PLoS ONE 2014, 9, e101622. [Google Scholar] [CrossRef] [Green Version]
- Dupaul-Chicoine, J.; Yeretssian, G.; Doiron, K.; Bergstrom, K.S.; McIntire, C.R.; LeBlanc, P.M.; Meunier, C.; Turbide, C.; Gros, P.; Beauchemin, N.; et al. Control of Intestinal Homeostasis, Colitis, and Colitis-Associated Colorectal Cancer by the Inflammatory Caspases. Immunity 2010, 32, 367–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, H.; Conway, K.L.; Zhang, M.; Choi, M.; Morin, B.; Cao, Z.; Villablanca, E.; Li, C.; Wijmenga, C.; Yun, S.H.; et al. Card9 Mediates Intestinal Epithelial Cell Restitution, T-Helper 17 Responses, and Control of Bacterial Infection in Mice. Gastroenterology 2013, 145, 591–601.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cario, E.; Gerken, G.; Podolsky, D. Toll-Like Receptor 2 Controls Mucosal Inflammation by Regulating Epithelial Barrier Function. Gastroenterology 2007, 132, 1359–1374. [Google Scholar] [CrossRef]
- Hsu, D.; Fukata, M.; Hernandez, Y.G.; Sotolongo, J.P.; Goo, T.; Maki, J.; Hayes, L.A.; Ungaro, R.C.; Chen, A.; Breglio, K.J.; et al. Toll-like receptor 4 differentially regulates epidermal growth factor-related growth factors in response to intestinal mucosal injury. Lab. Investig. 2010, 90, 1295–1305. [Google Scholar] [CrossRef] [Green Version]
- Krzywinska, E.; Sobecki, M.; Nagarajan, S.; Zacharjasz, J.; Tambuwala, M.M.; Pelletier, A.; Cummins, E.; Gotthardt, D.; Fandrey, J.; Kerdiles, Y.M.; et al. The transcription factor HIF-1α mediates plasticity of NKp46+ innate lymphoid cells in the gut. J. Exp. Med. 2022, 219, e20210909. [Google Scholar] [CrossRef] [PubMed]
- Koch, S.; Nava, P.; Addis, C.; Kim, W.; Denning, T.L.; Li, L.; Parkos, C.A.; Nusrat, A. The Wnt Antagonist Dkk1 Regulates Intestinal Epithelial Homeostasis and Wound Repair. Gastroenterology 2011, 141, 259–268.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Weight Loss | Stool Consistency | Faecal Blood |
|---|---|---|---|
| 0 | None | Normal | None |
| 1 | 1–5% | ||
| 2 | 5–10% | Loose | Hemoccult + |
| 3 | 10–20% | ||
| 4 | >20% | Diarrhoea | Gross bleeding |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almutary, A.G.; Alnuqaydan, A.M.; Almatroodi, S.A.; Tambuwala, M.M. Comparative Analysis of the Effect of Different Concentrations of Dextran Sodium Sulfate on the Severity and Extent of Inflammation in Experimental Ulcerative Colitis. Appl. Sci. 2023, 13, 3233. https://doi.org/10.3390/app13053233
Almutary AG, Alnuqaydan AM, Almatroodi SA, Tambuwala MM. Comparative Analysis of the Effect of Different Concentrations of Dextran Sodium Sulfate on the Severity and Extent of Inflammation in Experimental Ulcerative Colitis. Applied Sciences. 2023; 13(5):3233. https://doi.org/10.3390/app13053233
Chicago/Turabian StyleAlmutary, Abdulmajeed G., Abdullah M. Alnuqaydan, Saleh A. Almatroodi, and Murtaza M. Tambuwala. 2023. "Comparative Analysis of the Effect of Different Concentrations of Dextran Sodium Sulfate on the Severity and Extent of Inflammation in Experimental Ulcerative Colitis" Applied Sciences 13, no. 5: 3233. https://doi.org/10.3390/app13053233