
The Role of Beetroot Ingredients in the Prevention of Alzheimer’s Disease


Abstract
:1. Introduction
2. General Characteristics of Alzheimer’s Disease
2.1. Neuroprotective Properties of Beets in AD
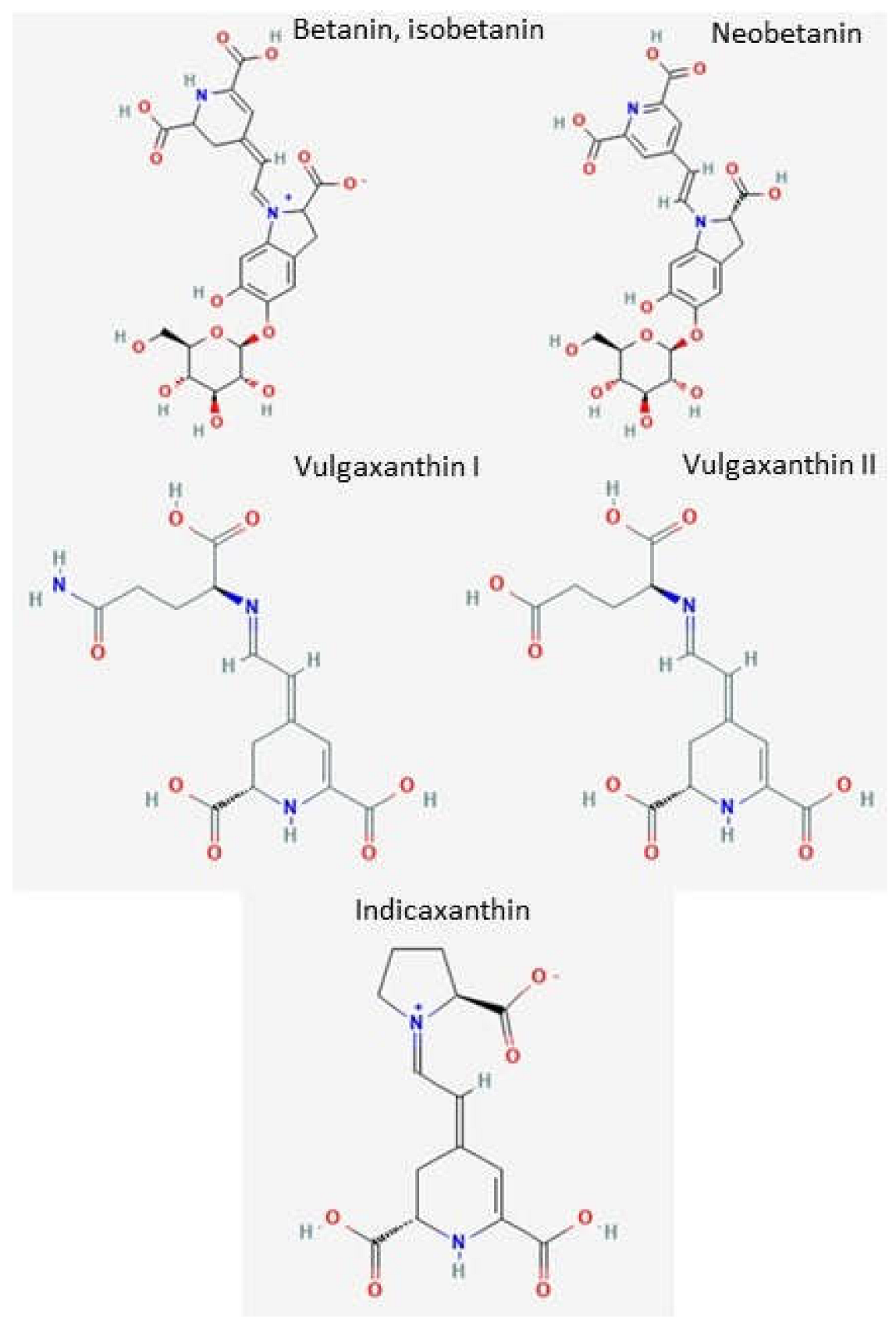
2.2. Individual Bioactive Compounds from Beetroots in the Prevention and Treatment of AD Nitric Pigments
3. Phenolic Compounds
4. Saponins
5. Changes in the Content of Bioactive Compounds in Beetroots during Processing
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABTS | 2,2-azinobis (3-ethylbenzothiazoline- 6-sulfonic acid) |
| AchE | Acetylcholinesterase |
| AD | Alzheimer’s disease |
| BHT | Butylated hydroxytoluene |
| b.w. | Body weight |
| DMSA | 2,3-dimercaptosuccinic acid |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| d.m. | Dry matter |
| FRAP | Ferric reducing antioxidant power |
| GSH | Glutathione |
| IC50 | Inhibitory concentrations 50% of cells |
| MDA | Malondialdehyde |
| TEAC | Value expressed as Trolox equivalent |
References
- Gitler, A.D.; Dhillon, P.; Shorter, J. Neurodegenerative disease: Models, mechanisms, and a new hope. Dis. Model. Mech. 2017, 10, 499–502. [Google Scholar] [CrossRef] [Green Version]
- Winiarska-Mieczan, A.; Baranowska-Wójcik, E.; Kwiecień, M.; Grela, E.R.; Szwajgier, D.; Kwiatkowska, K.; Kiczorowska, B. The role of dietary antioxidants in the pathogenesis of neurodegenerative diseases and their impact on cerebral oxidoreductive balance. Nutrients 2020, 12, 435. [Google Scholar] [CrossRef] [Green Version]
- Chin-Chan, M.; Navarro-Yepes, J.; Quintanilla-Vega, B. Environmental pollutants as risk factors for neurodegenerative disorders: Alzheimer and Parkinson diseases. Front. Cell. Neurosci. 2015, 9, 124. [Google Scholar] [CrossRef] [Green Version]
- Theodosis-Nobelos, P.; Rekka, E.A. The Multiple Sclerosis Modulatory Potential of Natural Multi-Targeting Antioxidants. Molecules 2022, 27, 8402. [Google Scholar] [CrossRef]
- Elfawy, H.A.; Das, B. Crosstalk between mitochondrial dysfunction, oxidative stress, and age-related neurodegenerative disease: Etiologies and therapeutic strategies. Life Sci. 2019, 218, 165–184. [Google Scholar] [CrossRef]
- Winiarska-Mieczan, A. Protective effect of tea against lead and cadmium-induced oxidative stress—A review. Biometals 2018, 31, 909–926. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.Y.; Ke, D.S.; Chen, J.Y. Essential fatty acids and human brain. Acta Neurol. Taiwan 2009, 18, 231–241. [Google Scholar]
- Lei, P.; Ayton, S.; Bush, A.I. The essential elements of Alzheimer’s disease. J. Biol. Chem. 2021, 296, 100105. [Google Scholar] [CrossRef]
- Fargo, K.N.; Aisen, P.; Albert, M.; Au, R.; Corrada, M.M.; DeKosky, S.; Drachman, D.; Fillit, H.; Gitlin, L.; Alzheimer’s Association National Plan Milestone Workgroup; et al. 2014 report on the milestones for the US national plan to address Alzheimer’s disease. Alzheimers Dement. 2014, 10, 430–452. [Google Scholar] [CrossRef] [Green Version]
- Peric, A.; Annaert, W. Early etiology of Alzheimer’s disease: Tipping the balance toward autophagy or endosomal dysfunction? Acta Neuropathol. 2015, 129, 363–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knopman, D.S.; Amieva, H.; Petersen, R.C.; Chételat, G.; Holtzman, D.M.; Hyman, B.T.; Nixon, R.A.; Jones, D.T. Alzheimer disease. Nat. Rev. Dis. Prim. 2021, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Prince, M.; Albanese, E.; Guerchet, M.; Prina, M. World Alzheimer Report 2014: Dementia and Risk Reduction: An Analysis of Protective and Modifiable Risk Factors; Alzheimer’s Disease International: London, UK, 2014. [Google Scholar]
- Buccafusco, J.J.; Terry, A.V. Multiple central nervous system targets for eliciting beneficial effects on memory and cognition. J. Pharmacol. Exp. Ther. 2000, 295, 438–446. [Google Scholar] [PubMed]
- Spires-Jones, T.L.; Hyman, B.T. The intersection of amyloid beta and tau at synapses in Alzheimer’s disease. Neuron 2014, 82, 756–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.O.; Alperstein, A.M.; Zanni, M.T. Amyloid β-sheet secondary structure identified in UV-induced cataracts of porcine lenses using 2D IR spectroscopy. J. Mol. Biol. 2017, 429, 1705–1721. [Google Scholar] [CrossRef] [PubMed]
- Rice, H.C.; de Malmazet, D.; Schreurs, A.; Frere, S.; Van Molle, I.; Volkov, A.N.; Creemers, E.; Vertkin, I.; Nys, J.; Ranaivoson, F.M.; et al. Secreted amyloid-β precursor protein functions as a GABABR1a ligand to modulate synaptic transmission. Science 2019, 363, eaao4827. [Google Scholar] [CrossRef] [PubMed]
- Kopeikina, K.; Hyman, B.; Spires-Jones, T. Soluble forms of tau are toxic in Alzheimer’s disease. Transl. Neurosci. 2012, 3, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muralidar, S.; Ambi, S.V.; Sekaran, S.; Thirumalai, D.; Palaniappan, B. Role of tau protein in Alzheimer’s disease: The prime pathological player. Int. J. Biol. Macromol. 2020, 163, 1599–1617. [Google Scholar] [CrossRef]
- Hoskin, J.L.; Sabbagh, M.N.; Al-Hasan, Y.; Decourt, B. Tau immunotherapies for Alzheimer’s disease. Expert Opin. Investig. Drugs 2019, 6, 545–554. [Google Scholar] [CrossRef]
- Ogawa, S. Nutritional management of older adults with cognitive decline and dementia. J. Gerontol. Geriatr. 2014, 14, 17–22. [Google Scholar] [CrossRef]
- Baiao, D.D.S.; da Silva, D.V.; Del Aguila, E.M.; Paschoalin, V.M.F. Nutritional, bioactive and physicochemical characteristics of different beetroot formulations. Food Addit. 2017, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Chhikara, N.; Kushwaha, K.; Sharma, P.; Gat, Y.; Panghal, A. Bioactive compounds of beetroot and utilization in food processing industry: A critical review. Food Chem. 2019, 272, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Ceclu, L.; Nistor, O.V. Red Beetroot: Composition and Health Effects-A Review. J. Nutr. Med. Diet Care 2020, 6, 043. [Google Scholar] [CrossRef]
- Wootton-Beard, P.C.; Ryan, L. A beetroot juice shot is a significant and convenient source of bioaccessible antioxidants. J. Funct. Foods 2011, 4, 329–334. [Google Scholar] [CrossRef]
- Presley, T.D.; Morgan, A.R.; Bechtold, E.; Clodfelter, W.; Dove, R.W.; Jennings, J.M.; Kraft, R.A.; King, S.B.; Laurienti, P.J.; Rejeski, W.J.; et al. Acute effect of a high nitrate diet on brain perfusion in older adults. Nitric Oxide 2011, 1, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Wightman, E.L.; Haskell-Ramsay, C.F.; Thompson, K.G.; Blackwell, J.R.; Winyard, P.G.; Forster, J.; Jones, A.M.; Kennedy, D.O. Dietary nitrate modulates cerebral blood flow parameters and cognitive performance in humans: A double-blind, placebo-controlled, crossover investigation. Physiol. Behav. 2015, 149, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulakhiya, K.; Patel, V.K.; Saxena, R.; Dashore, J.; Srivastava, A.K.; Rathore, M. Effect of Beta vulgaris Linn. leaves extract on anxiety-and depressive-like behavior and oxidative stress in mice after acute restraint stress. Phcog. Res. 2016, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mokhtari, S.; Rabiei, Z.; Shahrani, M.; Rafieian-Kopaei, M. The Ameliorating Effect of Beta Vulgaris Extract on Memory and Learning Impairment Induced by Lesions of the Nucleus Basalis of Meynert in Rat. J. Clin. Diagnostic Res. 2017, 11, CC09–CC14. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Oyeleye, S.I.; Ibeji, C.U.; Oboh, G. Beetroot supplemented diet exhibit anti-amnesic effect via modulation of cholinesterases, purinergic enzymes, monoamine oxidase and attenuation of redox imbalance in the brain of scopolamine treated male rats. Nutr. Neurosci. 2020, 5, 1011–1025. [Google Scholar] [CrossRef]
- Rehman, S.; Shah, S.; Butt, A.M.; Shah, S.M.; Jabeen, Z.; Nadeem, A. Biochemical profiling and elucidation of biological activities of Beta vulgaris L. leaves and roots extracts. Saudi J. Biol. Sci. 2021, 1, 592–602. [Google Scholar] [CrossRef]
- Aliahmadi, M.; Amiri, F.; Bahrami, L.S.; Hosseini, A.F.; Abiri, B.; Vafa, M. Effects of raw red beetroot consumption on metabolic markers and cognitive function in type 2 diabetes patients. J. Diabetes Metab. Disord. 2021, 20, 673–682. [Google Scholar] [CrossRef]
- Ertas, B.; Topa, F.; Gülhan, R.; Yanardağ, R.; Sacan, Ö.; Sener, G. Beta vulgaris L. var. cicla improves memory deficits in intracerebroventricular streptozotocin injected rats: Role on neuroinflammation. J. Res. Pharm. 2021, 5, 589–599. [Google Scholar] [CrossRef]
- Shaban, N.Z.; Abd El-Kader, S.E.; Mogahed, F.A.; El-Kersh, M.A.; Habashy, N.H. Synergistic protective effect of Beta vulgaris with meso-2,3-dimercaptosuccinic acid against lead-induced neurotoxicity in male rats. Sci. Rep. 2021, 11, 252. [Google Scholar] [CrossRef]
- Wulandari, D.D.; Santoso, A.P.R.; Wulansari, D.D. The Effect of Beetroot (Beta vulgaris L.) Juice on Cholinesterase Activity in Farmers Exposed to Organophosphate Pesticides. Indones. J. Med Lab. Sci. Technol. 2019, 2, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Nemzer, B.; Pietrzkowski, Z.; Spórna, A.; Stalica, P.; Thresher, W.; Michałowski, T.; Wybraniec, S. Betalainic and nutritional profiles of pigment-enriched red beet root (Beta vulgaris L.) dried extracts. Food Chem. 2011, 1, 42–53. [Google Scholar] [CrossRef]
- Mikołajczyk-Bator, K.; Czapski, J. Effect of pH changes on antioxidant capacity and the content of betalain pigments during the heating of a solution of red beet betalains. Pol. J. Food Nutr. Sci. 2017, 2, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Gengatharan, A.; Dykes, G.A.; Choo, W.S. Betalains: Natural plant pigments with potential application in functional foods. LWT Food Sci. Technol. 2015, 2, 645–649. [Google Scholar] [CrossRef]
- Kathiravan, T.; Nadanasabapathi, S.; Kumar, R. Standardization of process condition in batch thermal pasteurization and its effect on antioxidant, pigment and microbial inactivation of Ready to Drink (RTD) beetroot (Beta vulgaris L.) juice. Int. Food Res. J. 2014, 4, 1305–1312. [Google Scholar]
- Cai, Y.; Sun, M.; Corke, H. Antioxidant activity of betalains from plants of the Amaranthaceae. J. Agric. Food Chem. 2003, 8, 2288–2294. [Google Scholar] [CrossRef]
- Vulić, J.J.; Ćebović, T.N.; Čanadanović-Brunet, J.M.; Ćetković, G.S.; Čanadanović, V.M.; Djilas, S.M.; Šaponjac, V.T.T. In vivo and in vitro antioxidant effects of beetroot pomace extracts. J. Funct. Foods 2014, 6, 168–175. [Google Scholar] [CrossRef]
- Hacioglu, C.; Kar, F.; Kanbak, G. Rat brain synaptosomes: In vitro neuroprotective effects of betaine against fluoride toxicity. Med. Sci. Discov. 2018, 5, 192–197. [Google Scholar] [CrossRef] [Green Version]
- Motawi, T.K.; Ahmed, S.A.; El-Boghdady, N.A.; Metwally, N.S.; Nasr, N.N. Protective effects of betanin against paracetamol and diclofenac induced neurotoxicity and endocrine disruption in rats. J. Biomark. 2019, 7, 645–651. [Google Scholar] [CrossRef]
- Thong-Asa, W.; Prasartsri, S.; Klomkleaw, N.; Thongwan, N. The neuroprotective effect of betanin in trimethyltin-induced neurodegeneration in mice. Metab. Brain Dis. 2020, 8, 1395–1405. [Google Scholar] [CrossRef]
- Imamura, T.; Koga, H.; Higashimura, Y.; Isozumi, N.; Matsumoto, K.; Ohki, S.; Mori, M. Red-beet betalain pigments inhibit amyloid-β aggregation and toxicity in amyloid-β expressing Caenorhabditis elegans. Plant Foods Hum. Nutr. 2022, 1, 90–97. [Google Scholar] [CrossRef]
- Elkewawy, H.; Elbadrawy, E. The Protective effect of Pomegranate and Red Beetroot Juices against Aluminum Toxicity in Brains of Male Rats. J. Dairy. Sci. 2020, 1, 1–8. [Google Scholar] [CrossRef]
- Shunan, D.; Yu, M.; Guan, H.; Zhou, Y. Neuroprotective effect of Betalain against AlCl3-induced Alzheimer’s disease in Sprague Dawley Rats via putative modulation of oxidative stress and nuclear factor kappa B (NF-κB) signaling pathway. Biomed. Pharmacother. 2021, 137, 111369. [Google Scholar] [CrossRef]
- Ahmadi, H.; Nayeri, Z.; Minuchehr, Z.; Sabouni, F.; Mohammadi, M. Betanin purification from red beetroots and evaluation of its anti-oxidant and anti-inflammatory activity on LPS-activated microglial cells. PLoS ONE 2020, 5, e0233088. [Google Scholar] [CrossRef]
- Vauzour, D. Effect of flavonoids on learning, memory and neurocognitive performance: Relevance and potential implications for Alzheimer’s disease pathophysiology. J. Sci. Food Agric. 2014, 6, 1042–1056. [Google Scholar] [CrossRef]
- Zhao, Z.; Moghadasian, M.H. Chemistry, natural sources, dietary intake and pharmacokinetic properties of ferulic acid: A review. Food Chem. 2008, 4, 691–702. [Google Scholar] [CrossRef]
- Maraie, N.K.; Abdul-Jalil, T.Z.; Alhamdany, A.T.; Janabi, H.A. Phytochemical study of the Iraqi Beta vulgaris leaves and its clinical applications for the treatment of different dermatological diseases. World J. Pharm. Pharm. Sci. 2014, 8, 5–19. [Google Scholar]
- Baiao, D.D.S.; da Silva, D.V.; Paschoalin, V.M. Beetroot, a remarkable vegetable: Its nitrate and phytochemical contents can be adjusted in novel formulations to benefit health and support cardiovascular disease therapies. Antioxidants 2020, 9, 960. [Google Scholar] [CrossRef]
- Leite, A.; Leite, D.C.D.A.; Del Aguila, E.; Alvares, T.; Peixoto, R.; Miguel, M.; Silva, J.; Paschoalin, V. Microbiological and chemical characteristics of Brazilian kefir during fermentation and storage processes. J. Dairy Sci. 2013, 7, 4149–4159. [Google Scholar] [CrossRef] [Green Version]
- Wruss, J.; Waldenberger, G.; Huemer, S.; Uygun, P.; Lanzerstorfer, P.; Müller, U.; Höglinger, O.; Weghuber, J. Compositional characteristics of commercial beetroot products and beetroot juice prepared from seven beetroot varieties grown in Upper Austria. J. Food Compost. Anal. 2015, 42, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, F.H.A.; Salgado, H.R.N. Gallic acid: Review of the methods of determination and quantification. Crit. Rev. Anal. Chem. 2016, 3, 257–265. [Google Scholar] [CrossRef]
- Sharma, N.; Tiwari, N.; Vyas, M.; Khurana, N.; Muthuraman, A.; Utreja, P. An overview of therapeutic effects of vanillic acid. Plant Arch. 2020, 20, 3053–3059. [Google Scholar]
- Singh, J.C.H.; Kakalij, R.M.; Kshirsagar, R.P.; Kumar, B.H.; Komakula, S.S.B.; Diwan, P.V. Cognitive effects of vanillic acid against streptozotocin-induced neurodegeneration in mice. Pharm. Biol. 2015, 5, 630–636. [Google Scholar] [CrossRef]
- Alam, M. Anti-hypertensive effect of cereal antioxidant ferulic acid and its mechanism of action. Front. Nutr. 2019, 6, 121. [Google Scholar] [CrossRef]
- Singh, Y.P.; Rai, H.; Singh, G.; Singh, G.K.; Mishra, S.; Kumar, S.; Srikrishna, S.; Modi, G. A review on ferulic acid and analogs-based scaffolds for the management of Alzheimer’s disease. Eur. J. Med. Chem. 2021, 215, 113278. [Google Scholar] [CrossRef]
- Sgarbossa, A.; Giacomazza, D.; Di Carlo, M. Ferulic acid: A hope for Alzheimer’s disease therapy from plants. Nutrients 2015, 7, 5764–5782. [Google Scholar] [CrossRef] [Green Version]
- Ono, K.; Hirohata, M.; Yamada, M. Ferulic acid destabilizes preformed β-amyloid fibrils in vitro. Biochem. Biophys. Res. Commun. 2005, 2, 444–449. [Google Scholar] [CrossRef]
- Yan, J.J.; Jung, J.S.; Kim, T.K.; Hasan, M.A.; Hong, C.W.; Nam, J.S.; Song, D.K. Protective effects of ferulic acid in amyloid precursor protein plus presenilin-1 transgenic mouse model of Alzheimer disease. Biol. Pharm. Bull. 2013, 1, 140–143. [Google Scholar] [CrossRef] [Green Version]
- Mori, T.; Koyama, N.; Guillot-Sestier, M.V.; Tan, J.; Town, T. Ferulic acid is a nutraceutical β-secretase modulator that improves behavioral impairment and alzheimer-like pathology in transgenic mice. PLoS ONE 2013, 8, e55774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q. Effect of Ferulic Acid on Learning and Memory Impairment by the Repairing of Mitochondrial Fission-Fusion Imbalance in AD Mice. Chin. Pharm. J. 2019, 24, 703–710. [Google Scholar]
- Wang, E.J.; Wu, M.Y.; Lu, J.H. Ferulic Acid in Animal Models of Alzheimer’s Disease: A Systematic Review of Preclinical Studies. Cells 2021, 10, 2653. [Google Scholar] [CrossRef]
- Mori, T.; Koyama, N.; Tan, J.; Segawa, T.; Maeda, M.; Town, T. Combination therapy with octyl gallate and ferulic acid improves cognition and neurodegeneration in a transgenic mouse model of Alzheimer’s disease. Int. J. Biol. Chem. 2017, 27, 11310–11325. [Google Scholar] [CrossRef] [Green Version]
- Szwajgier, D.; Pielecki, J.; Targoński, Z. Antioxidant activities of cinnamic and benzoic acid derivaties. Acta Sci. Pol. Technol. Aliment. 2005, 2, 129–142. [Google Scholar]
- Albasher, G.; Alsaleh, A.S.; Alkubaisi, N.; Alfarraj, S.; Alkahtani, S.; Farhood, M.; Almeer, R. Red beetroot extract abrogates chlorpyrifos-induced cortical damage in rats. Oxid. Med. Cell. Longev. 2020, 2020, 2963020. [Google Scholar] [CrossRef] [Green Version]
- Vincken, J.P.; Heng, L.; de Groot, A.; Gruppen, H. Saponins, classification and occurrence in the plant kingdom. Phytochemistry 2007, 3, 275–297. [Google Scholar] [CrossRef]
- Mroczek, A.; Kapusta, I.; Janda, B.; Janiszowska, W. Triterpene saponin content in the roots of red beet (Beta vulgaris L.) cultivars. J. Agric. Food Chem. 2012, 50, 12397–12402. [Google Scholar] [CrossRef]
- Mikolajczyk-Bator, K.; Kikut-Ligaj, D. Triterpene saponins as bitter components of beetroot. Żywność Nauka Technol. Jakość 2016, 1, 128–141. [Google Scholar] [CrossRef]
- Mikołajczyk-Bator, K.; Błaszczyk, A.; Czyżniejewski, M.; Kachlicki, P. Characterisation and identification of triterpene saponins in the roots of red beets (Beta vulgaris L.) using two HPLC–MS systems. Food Chem. 2016, 192, 979–990. [Google Scholar] [CrossRef]
- Mroczek, A.; Kapusta, I.; Stochmal, A.; Janiszowska, W. MS/MS and UPLC-MS profiling of triterpenoid saponins from leaves and roots of four red beet (Beta vulgaris L.) cultivars. Phytochem. Lett. 2019, 30, 333–337. [Google Scholar] [CrossRef]
- Khalil, W.K.; Roshdy, H.M.; Kassem, S.M. The potential therapeutic role of Fenugreek saponin against Alzheimer’s disease: Evaluation of apoptotic and acetylcholinesterase inhibitory activities. J. Appl. Pharm. Sci. 2016, 9, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Różańska, D.; Regulska-Ilow, B.; Ilow, R. Influence of selected culinary processes on the antioxidant capacity and polyphenol content in food. Probl. Hig. Epidemiol. 2014, 95, 215–222. [Google Scholar]
- Szwajgier, D.; Baranowska-Wójcik, E.; Sosnowska, B.; Kukula-Koch, W.; Paduch, R.; Sokołowska, B.; Waśko, A.; Solarska, E. High-pressure processing at increased temperatures provides enhanced removal of indigenous microbial contamination in beet/carrot juice without damaging the bioactive components. Int. J. Food Sci. Technol. 2022, 1, 276–289. [Google Scholar] [CrossRef]
- Sawicki, T.; Wiczkowski, W.; Hrynkiewicz, M.; Bączek, N.; Hornowski, A.; Honke, J.; Topolska, J. Characterization of the phenolic acid profile and in vitro bioactive properties of white beetroot products. Int. J. Food Sci. Technol. 2021, 2, 629–638. [Google Scholar] [CrossRef]
- Sawicki, T.; Wiczkowski, W. The effects of boiling and fermentation on betalain profiles and antioxidant capacities of red beetroot products. Food Chem. 2018, 259, 292–303. [Google Scholar] [CrossRef]
- Ramírez-Melo, L.M.; del Socorro Cruz-Cansino, N.; Delgado-Olivares, L.; Ramírez-Moreno, E.; Zafra-Rojas, Q.Y.; Hernández-Traspeña, J.L.; Suárez-Jacobo, Á. Optimization of antioxidant activity properties of a thermosonicated beetroot (Beta vulgaris L.) juice and further in vitro bioaccessibility comparison with thermal treatments. LWT 2022, 154, 112780. [Google Scholar] [CrossRef]
- Bárta, J.; Bártová, V.; Sindelkova, T. Effect of boiling on colour, contents of betalains and total phenolics and on antioxidant activity of colourful powder derived from six different beetroot (Beta vulgaris L. var. conditiva) cultivars. Pol. J. Food Nutr. Sci. 2020, 4, 377–385. [Google Scholar] [CrossRef]
- Dalmau, M.E.; Eim, V.; Rosselló, C.; Cárcel, J.A.; Simal, S. Effects of convective drying and freeze-drying on the release of bioactive compounds from beetroot during in vitro gastric digestion. Food Funct. 2019, 6, 3209–3223. [Google Scholar] [CrossRef]
- Kaur, S.; Kaur, N.; Aggarwal, P.; Grover, K. Bioactive compounds, antioxidant activity, and color retention of beetroot (Beta vulgaris L.) powder: Effect of steam blanching with refrigeration and storage. J. Food Process. Preserv. 2021, 45, e15247. [Google Scholar] [CrossRef]
- Mohsen, S.; Alsaman, M.; Mahrous, H. A Study of the effect of Lactic acid bacteria on antioxidant content of beetroot (Beta vulgaris). Biotechnol. Res. 2018, 3, 103–108. [Google Scholar]


{kind=link}
| Name of Compound | Reference |
|---|---|
| Ferulic acid | [22,49] |
| 5,50,6,60-tetrahydroxy-3,30-biindolyl; 5,6-dihydroxyindole carboxylic acid dimer; phenol amides—N-trans-feruloyltyramine; N-trans-feruloylohomovanillylamine | [35] |
| Phenolic acids: caffeic, vanillic, p-coumaric, protocatechuic | [50] |
| Betagarin, vanillic acid, betavulgarin, p-coumaric acid | [51] |
| Gallic acid | [52,53,54] |
| Betagarin, cochliofilin A, betavulgarin, dihydroisorhamnetin | [40] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szymański, J.; Szwajgier, D.; Baranowska-Wójcik, E. The Role of Beetroot Ingredients in the Prevention of Alzheimer’s Disease. Appl. Sci. 2023, 13, 1044. https://doi.org/10.3390/app13021044
Szymański J, Szwajgier D, Baranowska-Wójcik E. The Role of Beetroot Ingredients in the Prevention of Alzheimer’s Disease. Applied Sciences. 2023; 13(2):1044. https://doi.org/10.3390/app13021044
Chicago/Turabian StyleSzymański, Julian, Dominik Szwajgier, and Ewa Baranowska-Wójcik. 2023. "The Role of Beetroot Ingredients in the Prevention of Alzheimer’s Disease" Applied Sciences 13, no. 2: 1044. https://doi.org/10.3390/app13021044