

The Influences of Self-Introspection and Credit Evaluation on Self-Organized Flocking
 , and
, and
Abstract
:1. Introduction
2. Models
2.1. Vicsek Model (VM)
2.2. Two Models Based on Local Consistency
2.2.1. Interaction Mechanisms under Local Consistency
2.2.2. Two Models
- (1)
- Self-introspection model (SIM)
- (2)
- Credit Evaluation Model (CEM)
3. Simulations and Discussions
| Algorithm 1: |
| Step 1. Randomly assign the position (xi, yi) and velocity (vi, θi) for ith individual, i = 1, 2, ⋯, N, with (xi, yi, θi) ∈ P, vi = v is a constant. |
 |
| Step 3. Set T = t. |
3.1. Noise-Free Calculation and Analysis
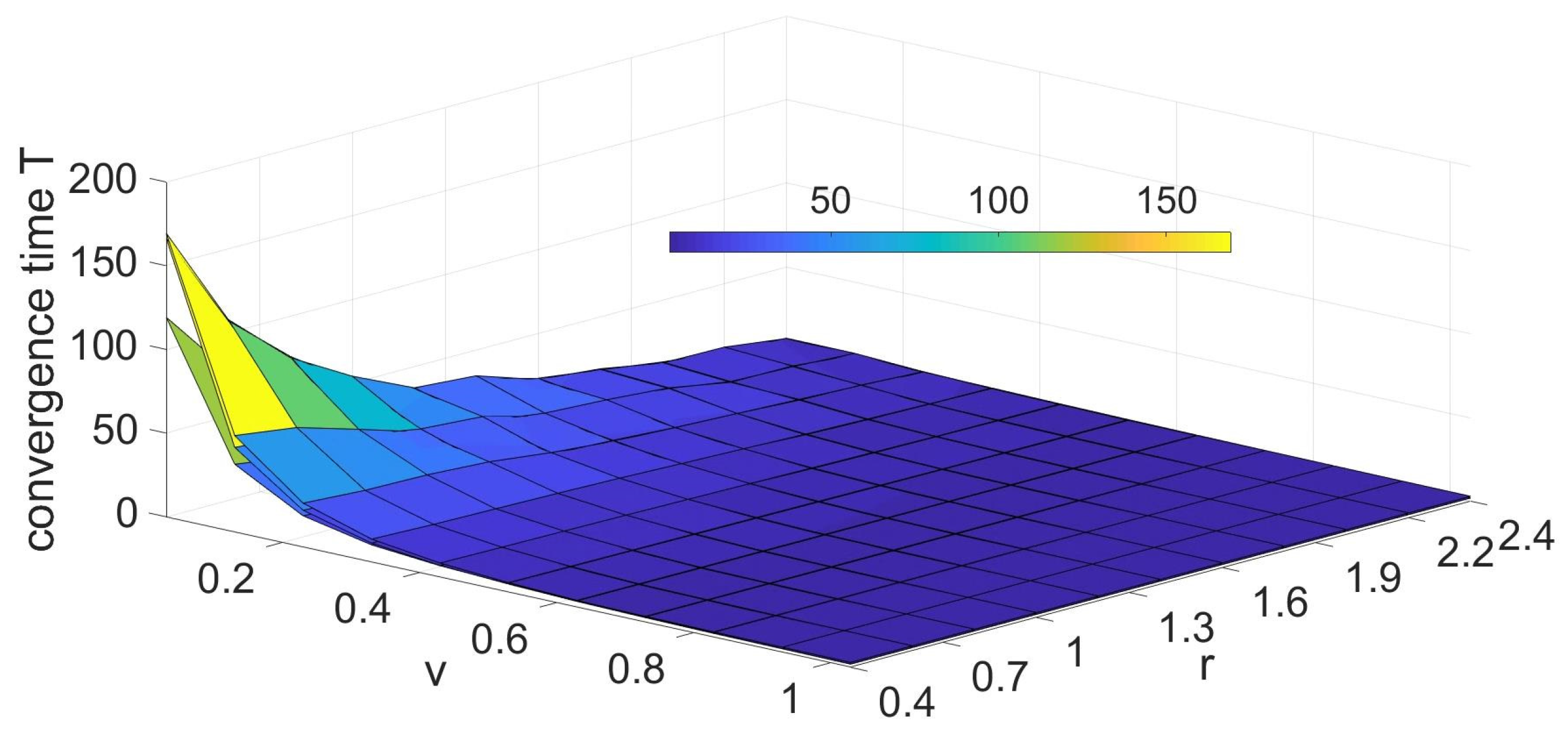
3.1.1. Influence of Parameters on the Convergence Time T
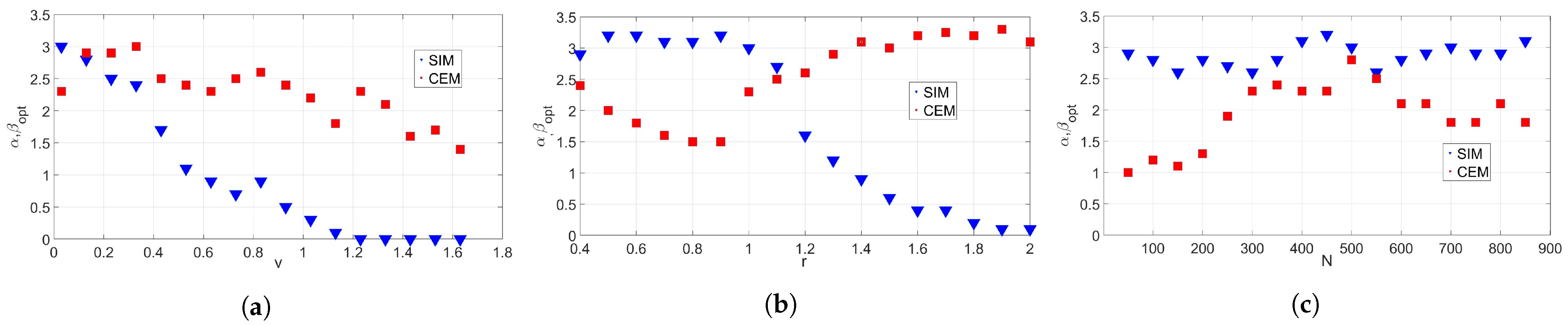
3.1.2. Optimal Values of the Model Parameters
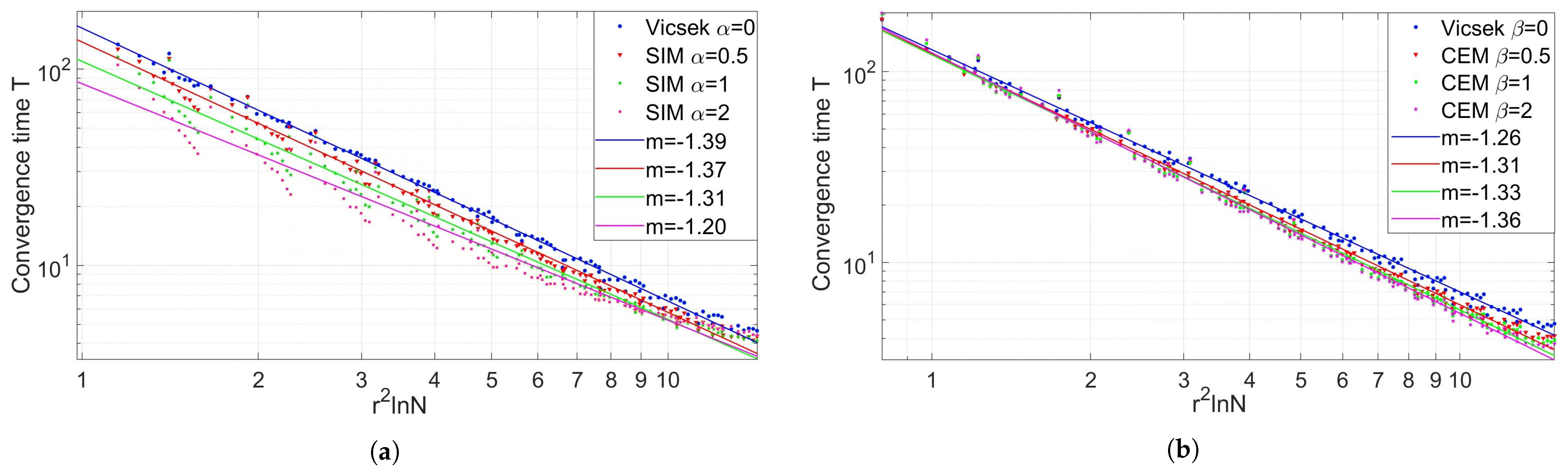
3.1.3. Model Comparison
3.2. Simulations with Noise
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Debets, V.E.; Janssen, L.; Storm, C. Enhanced persistence and collective migration in cooperatively aligning cell clusters. Biophys. J. 2021, 120, 1483–1497. [Google Scholar] [CrossRef] [PubMed]
- Vourc’h, T.; Léopoldès, J.; Peerhossaini, H. Clustering of bacteria with heterogeneous motility. Phys. Rev. E 2020, 101, 022612. [Google Scholar] [CrossRef] [PubMed]
- Larrieu, R.; Quilliet, C.; Dupont, A.; Peyla, P. Collective orientation of an immobile fish school and effect on rheotaxis. Phys. Rev. E 2021, 103, 022137. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Mclvor, G.E.; van der Vaart, K.; Vaughan, R.T.; Thornton, A.; Ouellette, N.T. Local interactions and their group-level consequences in flocking jackdaws. Proc. R. Soc. B 2019, 286, 20190865. [Google Scholar] [CrossRef] [PubMed]
- Torney, C.J.; Lamont, M.; Debell, L.; Angohiatok, R.J.; Leclerc, L.M.; Berdahl, A.M. Inferring the rules of social interaction in migrating caribou. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170385. [Google Scholar] [CrossRef]
- Helbing, D.; Farkas, I.; Vicsek, T. Simulating dynamical features of escape panic. Nature 2000, 407, 487–490. [Google Scholar] [CrossRef]
- Gu, Z.; Pan, S.; Lin, Z.; Hu, L.; Zhan, X. Climate-driven flyway changes and memory-based long-distance migration. Nature 2021, 591, 259–264. [Google Scholar] [CrossRef]
- Janosov, M.; Virágh, C.; Vásárhelyi, G.; Vicsek, T. Group chasing tactics: How to catch a faster prey? New J. Phys. 2017, 19, 053003. [Google Scholar] [CrossRef]
- Abdi, H.; Pishkenari, H.N. Controlled swarm motion of self-propelled microswimmers for energy saving. J. Micro-Bio Robot. 2021, 17, 23–33. [Google Scholar] [CrossRef]
- Hughey, L.F.; Hein, A.M.; Strandburg-Peshkin, A.; Jensen, F.H. Challenges and solutions for studying collective animal behaviour in the wild. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170005. [Google Scholar] [CrossRef]
- Papageorgiou, D.; Farine, D.R. Group size and composition influence collective movement in a highly social terrestrial bird. Elife 2020, 9, e59902. [Google Scholar] [CrossRef] [PubMed]
- Sarfati, R.; Hayes, J.C.; Peleg, O. Self-organization in natural swarms of Photinus carolinus synchronous fireflies. Sci. Adv. 2021, 7, eabg9259. [Google Scholar] [CrossRef] [PubMed]
- Olfati-Saber, R. Flocking for multi-agent dynamic systems: Algorithms and theory. IEEE Trans. Autom. Control 2006, 51, 401–420. [Google Scholar] [CrossRef]
- Giardina, I. Collective behavior in animal groups: Theoretical models and empirical studies. HFSP J. 2008, 2, 205–219. [Google Scholar] [CrossRef]
- Ariel, G.; Ayali, A. Locust collective motion and its modeling. PLoS Comput. Biol. 2015, 11, e1004522. [Google Scholar] [CrossRef]
- Deutsch, A.; Theraulaz, G.; Vicsek, T. Collective motion in biological systems. Interface Focus 2012, 2. [Google Scholar] [CrossRef]
- Vicsek, T.; Czirók, A.; Ben-Jacob, E.; Cohen, I.; Shochet, O. Novel type of phase transition in a system of self-driven particles. Phys. Rev. Lett. 1995, 75, 1226. [Google Scholar] [CrossRef]
- George, M.; Ghose, D. Reducing convergence times of self-propelled swarms via modified nearest neighbor rules. Phys. A Stat. Mech. Its Appl. 2012, 391, 4121–4127. [Google Scholar] [CrossRef]
- Aldana, M.; Dossetti, V.; Huepe, C.; Kenkre, V.; Larralde, H. Phase transitions in systems of self-propelled agents and related network models. Phys. Rev. Lett. 2007, 98, 095702. [Google Scholar] [CrossRef]
- Baglietto, G.; Albano, E.V.; Candia, J. Complex network structure of flocks in the Standard Vicsek Model. J. Stat. Phys. 2013, 153, 270–288. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, S.; Wang, G.; Wang, C.; Xie, G. A Local Consistency Algorithm to Shorten the Convergence Time and Improve the Robustness of Self-propelled Swarms. In Proceedings of the Chinese Automation Congress (CAC), Shanghai, China, 6–8 November 2020; pp. 4153–4157. [Google Scholar]
- Zou, Y.; Zhang, H.; Li, Y.; Wang, B. Accelerating consensus of self-driven swarm via a weighted model. Phys. A Stat. Mech. Its Appl. 2013, 392, 3329–3334. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, Y.; Tian, B.; Peng, L.; Zhang, H.; Wang, B.; Zhou, T. Accelerating consensus of self-driven swarm via adaptive speed. Phys. A Stat. Mech. Its Appl. 2009, 388, 1237–1242. [Google Scholar] [CrossRef]
- Zhao, M.; Su, H.; Wang, M.; Wang, L.; Chen, M.Z. A weighted adaptive-velocity self-organizing model and its high-speed performance. Neurocomputing 2016, 216, 402–408. [Google Scholar] [CrossRef]
- Shang, Y.; Bouffanais, R. Influence of the number of topologically interacting neighbors on swarm dynamics. Sci. Rep. 2014, 4, 4184. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xie, G. Lazy workers benefit group performance in circle formation tasks. IFAC-PapersOnLine 2017, 50, 10383–10388. [Google Scholar] [CrossRef]
- del Mar Delgado, M.; Miranda, M.; Alvarez, S.J.; Gurarie, E.; Fagan, W.F.; Penteriani, V.; di Virgilio, A.; Morales, J.M. The importance of individual variation in the dynamics of animal collective movements. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373. [Google Scholar] [CrossRef]
- Aguilar, J.; Monaenkova, D.; Linevich, V.; Savoie, W.; Dutta, B.; Kuan, H.S.; Betterton, M.; Goodisman, M.; Goldman, D. Collective clog control: Optimizing traffic flow in confined biological and robophysical excavation. Science 2018, 361, 672–677. [Google Scholar] [CrossRef]
- Albert, R.; Barabási, A.L. Statistical mechanics of complex networks. Rev. Mod. Phys. 2002, 74, 47. [Google Scholar] [CrossRef]
- Bode, N.W.; Wood, A.J.; Franks, D.W. Social networks and models for collective motion in animals. Behav. Ecol. Sociobiol. 2011, 65, 117–130. [Google Scholar] [CrossRef]
- Ojer, J.; Pastor-Satorras, R. Flocking dynamics mediated by weighted social networks. arXiv 2022, arXiv:2205.07533. [Google Scholar] [CrossRef]
- Eriksson, A.; Nilsson Jacobi, M.; Nyström, J.; Tunstrøm, K. Determining interaction rules in animal swarms. Behav. Ecol. 2010, 21, 1106–1111. [Google Scholar] [CrossRef]
- LaChance, J.; Suh, K.; Clausen, J.; Cohen, D.J. Learning the rules of collective cell migration using deep attention networks. PLoS Comput. Biol. 2022, 18, e1009293. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Escobedo, R.; Sanchez, S.; Sire, C.; Han, Z.; Theraulaz, G. The impact of individual perceptual and cognitive factors on collective states in a data-driven fish school model. PLoS Comput. Biol. 2022, 18, e1009437. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.C.; Parley, J.T.; Pastor-Satorras, R. Effects of heterogeneous social interactions on flocking dynamics. Phys. Rev. Lett. 2018, 120, 068303. [Google Scholar] [CrossRef] [PubMed]
- Demartsev, V.; Gersick, A.S.; Jensen, F.H.; Thomas, M.; Roch, M.A.; Manser, M.B.; Strandburg-Peshkin, A. Signalling in groups: New tools for the integration of animal communication and collective movement. Methods Ecol. Evol. 2022, 14, 1852–1863. [Google Scholar] [CrossRef]
- Cook, C.N.; Lemanski, N.J.; Mosqueiro, T.; Ozturk, C.; Gadau, J.; Pinter-Wollman, N.; Smith, B.H. Individual learning phenotypes drive collective behavior. Proc. Natl. Acad. Sci. USA 2020, 117, 17949–17956. [Google Scholar] [CrossRef]
- Tang, W.; Davidson, J.D.; Zhang, G.; Conen, K.E.; Fang, J.; Serluca, F.; Li, J.; Xiong, X.; Coble, M.; Tsai, T.; et al. Genetic control of collective behavior in zebrafish. iScience 2020, 23, 100942. [Google Scholar] [CrossRef]
- Cucker, F.; Huepe, C. Flocking with informed agents. Math. Action 2008, 1, 1–25. [Google Scholar] [CrossRef]
- Jolles, J.W.; Boogert, N.J.; Sridhar, V.H.; Couzin, I.D.; Manica, A. Consistent individual differences drive collective behavior and group functioning of schooling fish. Curr. Biol. 2017, 27, 2862–2868. [Google Scholar] [CrossRef]
- Cavagna, A.; Del Castello, L.; Giardina, I.; Grigera, T.; Jelic, A.; Melillo, S.; Mora, T.; Parisi, L.; Silvestri, E.; Viale, M.; et al. Flocking and turning: A new model for self-organized collective motion. J. Stat. Phys. 2015, 158, 601–627. [Google Scholar] [CrossRef]
- Procaccini, A.; Orlandi, A.; Cavagna, A.; Giardina, I.; Zoratto, F.; Santucci, D.; Chiarotti, F.; Hemelrijk, C.K.; Alleva, E.; Parisi, G.; et al. Propagating waves in starling, Sturnus vulgaris, flocks under predation. Anim. Behav. 2011, 82, 759–765. [Google Scholar] [CrossRef]
- Cavagna, A.; Cimarelli, A.; Giardina, I.; Parisi, G.; Santagati, R.; Stefanini, F.; Viale, M. Scale-free correlations in starling flocks. Proc. Natl. Acad. Sci. USA 2010, 107, 11865–11870. [Google Scholar] [CrossRef] [PubMed]
- Ballerini, M.; Cabibbo, N.; Candelier, R.; Cavagna, A.; Cisbani, E.; Giardina, I.; Orlandi, A.; Parisi, G.; Procaccini, A.; Viale, M.; et al. Empirical investigation of starling flocks: A benchmark study in collective animal behaviour. Anim. Behav. 2008, 76, 201–215. [Google Scholar] [CrossRef]
- Dill, L.M.; Holling, C.; Palmer, L.H. Predicting the three-dimensional structure of animal aggregations from functional considerations: The role of information. In Animal Groups in Three Dimensions; Cambridge University Press: Cambridge, UK, 1997; pp. 207–224. [Google Scholar]
- Wolf, M.; Weissing, F.J. Animal personalities: Consequences for ecology and evolution. Trends Ecol. Evol. 2012, 27, 452–461. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Simulation Settings | Convergence Time T | |||
|---|---|---|---|---|
| Number of Individuals N | Models | |||
| 100 | VM | 139.08 | 18.80 | 5.42 |
| SIM | 112.81 | 14.38 | 5.18 | |
| CEM | 135.22 | 15.60 | 4.35 | |
| 300 | VM | 93.82 | 14.16 | 4.74 |
| SIM | 57.39 | 9.07 | 4.30 | |
| CEM | 86.12 | 10.78 | 3.75 | |
| 500 | VM | 84.93 | 12.93 | 4.70 |
| SIM | 43.65 | 8.33 | 4.38 | |
| CEM | 78.51 | 10.45 | 3.82 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Q.; Luan, Y.; Li, S.; Wang, G.; Xu, M.; Wang, C.; Xie, G. The Influences of Self-Introspection and Credit Evaluation on Self-Organized Flocking. Appl. Sci. 2023, 13, 10361. https://doi.org/10.3390/app131810361
Zhao Q, Luan Y, Li S, Wang G, Xu M, Wang C, Xie G. The Influences of Self-Introspection and Credit Evaluation on Self-Organized Flocking. Applied Sciences. 2023; 13(18):10361. https://doi.org/10.3390/app131810361
Chicago/Turabian StyleZhao, Qiang, Yu Luan, Shuai Li, Gang Wang, Minyi Xu, Chen Wang, and Guangming Xie. 2023. "The Influences of Self-Introspection and Credit Evaluation on Self-Organized Flocking" Applied Sciences 13, no. 18: 10361. https://doi.org/10.3390/app131810361