


Changes in the Power and Coupling of Infra-Slow Oscillations in the Signals of EEG Leads during Stress-Inducing Cognitive Tasks

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
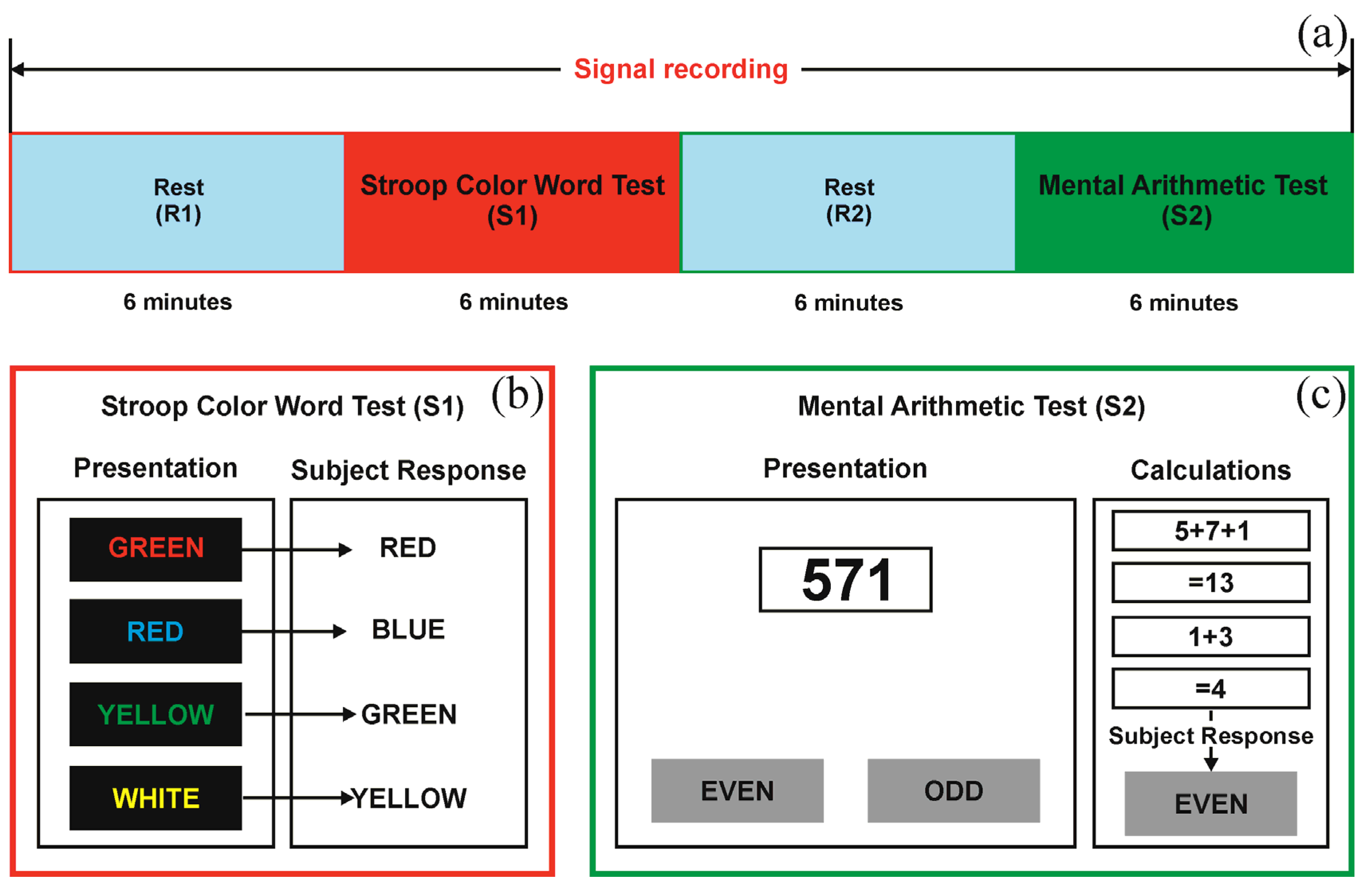
2.1. Design of the Study and the Experimental Data
2.2. Data Analysis
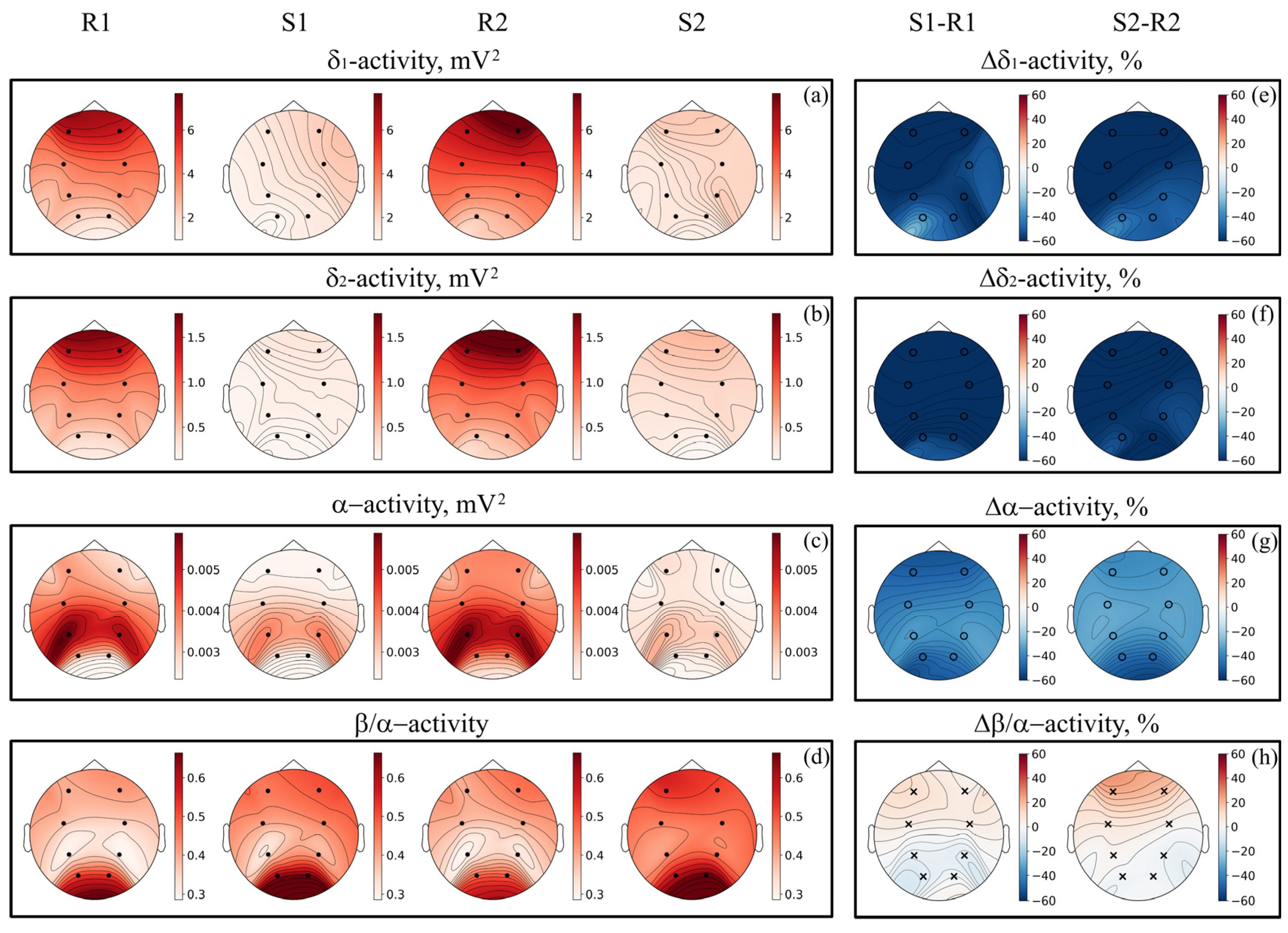
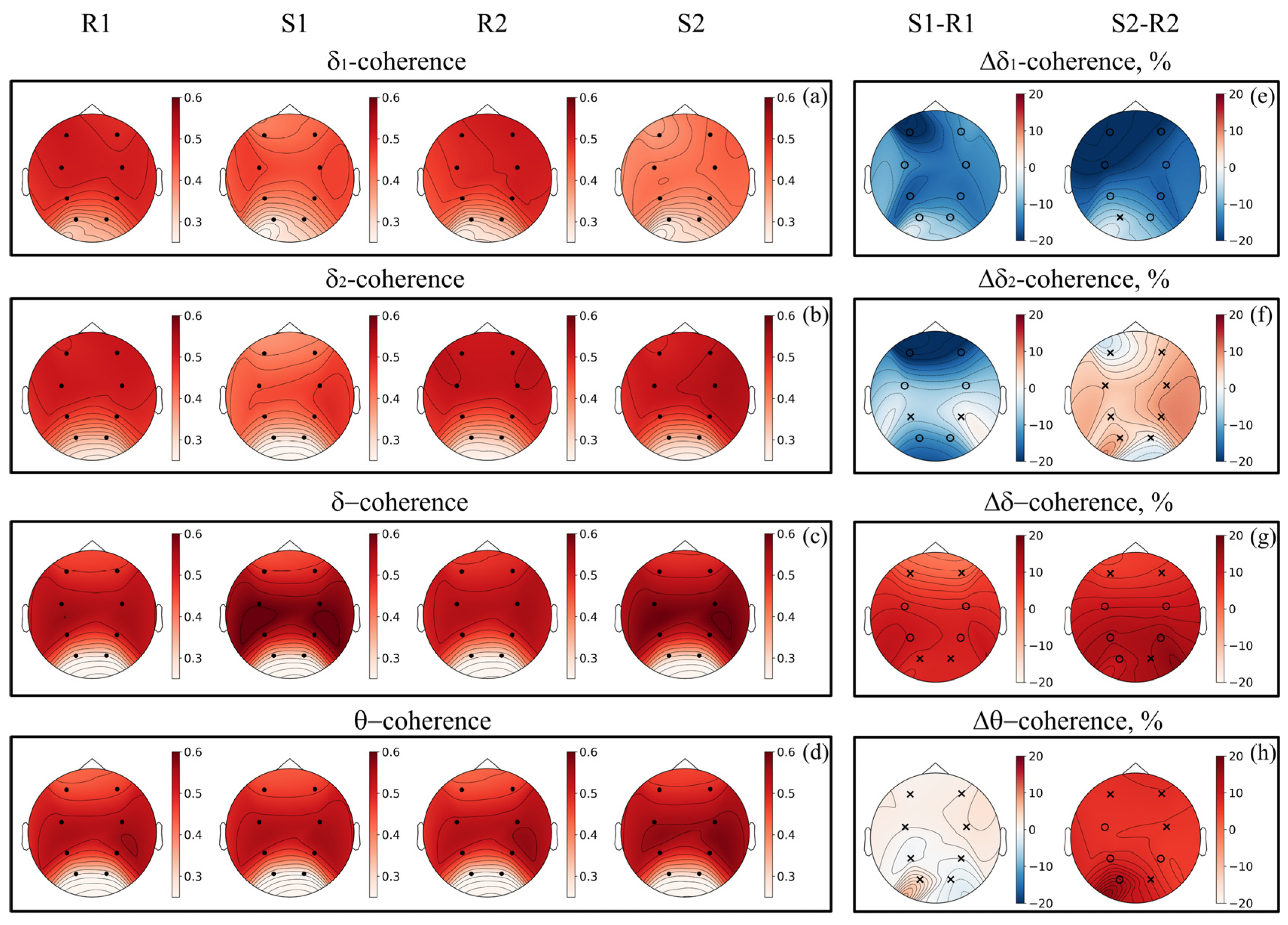
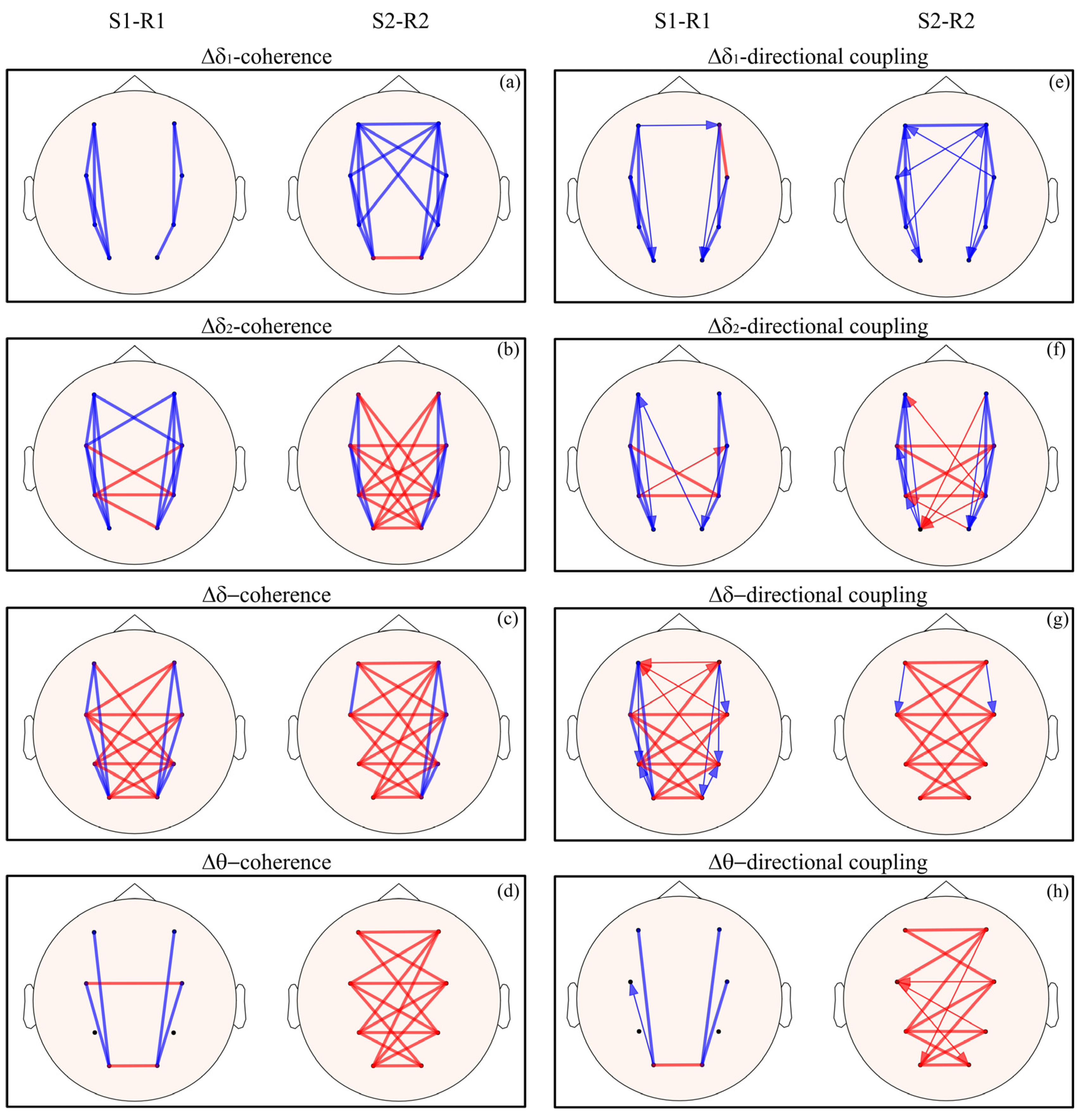
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Camm, A.J.; Malik, M.; Bigger, J.T.; Breithardt, G.; Cerutti, S.; Cohen, R.J.; Coumel, P.; Fallen, E.L.; Kennedy, H.L.; Kleiger, R.E.; et al. Heart Rate Variability: Standards of Measurement, Physiological Interpretation and Clinical Use. Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology. Circulation 1996, 93, 1043–1065. [Google Scholar]
- Wessel, N.; Riedl, M.; Kurths, J. Is the Normal Heart Rate “Chaotic” Due to Respiration? Chaos 2009, 19, 28508. [Google Scholar] [CrossRef] [PubMed]
- Aladjalova, N.A. Infra-Slow Rhythmic Oscillations of The Steady Potential of the Cerebral Cortex. Nature 1957, 179, 957–959. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J. Functional and Effective Connectivity: A Review. Brain Connect. 2011, 1, 13–36. [Google Scholar] [CrossRef]
- Katmah, R.; Al-Shargie, F.; Tariq, U.; Babiloni, F.; Al-Mughairbi, F.; Al-Nashash, H. A Review on Mental Stress Assessment Methods Using EEG Signals. Sensors 2021, 21, 5043. [Google Scholar] [CrossRef]
- Schulz, S.; Adochiei, F.-C.; Edu, I.-R.; Schroeder, R.; Costin, H.; Bär, K.-J.; Voss, A. Cardiovascular and Cardiorespiratory Coupling Analyses: A Review. Phil. Trans. R. Soc. A. 2013, 371, 20120191. [Google Scholar] [CrossRef] [Green Version]
- Giannakakis, G.; Grigoriadis, D.; Giannakaki, K.; Simantiraki, O.; Roniotis, A.; Tsiknakis, M. Review on Psychological Stress Detection Using Biosignals. IEEE Trans. Affect. Comput. 2022, 13, 440–460. [Google Scholar] [CrossRef]
- Iatsenko, D.; Bernjak, A.; Stankovski, T.; Shiogai, Y.; Owen-Lynch, P.J.; Clarkson, P.B.M.; McClintock, P.V.E.; Stefanovska, A. Evolution of Cardiorespiratory Interactions with Age. Phil. Trans. R. Soc. A. 2013, 371, 20110622. [Google Scholar] [CrossRef] [Green Version]
- Ponomarenko, V.I.; Karavaev, A.S.; Borovkova, E.I.; Hramkov, A.N.; Kiselev, A.R.; Prokhorov, M.D.; Penzel, T. Decrease of Coherence between the Respiration and Parasympathetic Control of the Heart Rate with Aging. Chaos 2021, 31, 73105. [Google Scholar] [CrossRef]
- Borovkova, E.I.; Hramkov, A.N.; Karavaev, A.S.; Ponomarenko, V.I.; Prokhorov, M.D.; Ishbulatov, Y.M.; Penzel, T. Directional Couplings Between Electroencephalogram and Interbeat Intervals Signals in Awake State and Different Stages of Sleep. In 2021 43rd Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC), Guadalajara, Mexico, 1–5 November 2021; IEEE: Mexico, 2021; pp. 5398–5402. [Google Scholar] [CrossRef]
- Karavaev, A.S.; Skazkina, V.V.; Borovkova, E.I.; Prokhorov, M.D.; Hramkov, A.N.; Ponomarenko, V.I.; Runnova, A.E.; Gridnev, V.I.; Kiselev, A.R.; Kuznetsov, N.V.; et al. Synchronization of the Processes of Autonomic Control of Blood Circulation in Humans Is Different in the Awake State and in Sleep Stages. Front. Neurosci. 2022, 15, 791510. [Google Scholar] [CrossRef]
- Borovkova, E.I.; Prokhorov, M.D.; Kiselev, A.R.; Hramkov, A.N.; Mironov, S.A.; Agaltsov, M.V.; Ponomarenko, V.I.; Karavaev, A.S.; Drapkina, O.M.; Penzel, T. Directional Couplings between the Respiration and Parasympathetic Control of the Heart Rate during Sleep and Wakefulness in Healthy Subjects at Different Ages. Front. Netw. Physiol. 2022, 2, 942700. [Google Scholar] [CrossRef]
- Karavaev, A.S.; Prokhorov, M.D.; Ponomarenko, V.I.; Kiselev, A.R.; Gridnev, V.I.; Ruban, E.I.; Bezruchko, B.P. Synchronization of Low-Frequency Oscillations in the Human Cardiovascular System. Chaos 2009, 19, 33112. [Google Scholar] [CrossRef] [Green Version]
- Bockeria, O.L.; Shvartz, V.A.; Akhobekov, A.A.; Glushko, L.A.; Le, T.G.; Kiselev, A.R.; Prokhorov, M.D.; Bockeria, L.A. Statin Therapy in the Prevention of Atrial Fibrillation in the Early Postoperative Period after Coronary Artery Bypass Grafting: A Meta-Analysis. Cor Vasa 2017, 59, e266–e271. [Google Scholar] [CrossRef] [Green Version]
- Kiselev, A.R.; Gridnev, V.I.; Prokhorov, M.D.; Karavaev, A.S.; Posnenkova, O.M.; Ponomarenko, V.I.; Bezruchko, B.P.; Shvartz, V.A. Evaluation of 5-Year Risk of Cardiovascular Events in Patients after Acute Myocardial Infarction Using Synchronization of 0.1-Hz Rhythms in Cardiovascular System: 0.1-Hz Rhythms and 5-Year Risk. Ann. Noninvasive Electrocardiol. 2012, 17, 204–213. [Google Scholar] [CrossRef]
- Borovkova, E.I.; Hramkov, A.N.; Dubinkina, E.S.; Ponomarenko, V.I.; Bezruchko, B.P.; Ishbulatov, Y.M.; Kurbako, A.V.; Karavaev, A.S.; Prokhorov, M.D. Biomarkers of the Psychophysiological State during the Cognitive Tasks Estimated from the Signals of the Brain, Cardiovascular and Respiratory Systems. Eur. Phys. J. Spec. Top. 2023, 232, 625–633. [Google Scholar] [CrossRef]
- Kalauzi, A.; Kesic, S.; Saponjic, J. Cortico-pontine theta synchronization phase shift following monoaminergic lesion in rat. J. Physiol. Pharmacol. 2009, 60, 79–84. [Google Scholar]
- Kesic, S.; Kalauzi, A.; Radulovacki, M.; Carley, D.W.; Saponjic, J. Coupling changes in cortical and pontine sigma and theta frequency oscillations following monoaminergic lesions in rat. Sleep Breath. 2010, 15, 35–47. [Google Scholar] [CrossRef]
- Xia, L.; Malik, A.S.; Subhani, A.R. A Physiological Signal-Based Method for Early Mental-Stress Detection. In Cyber-Enabled Intelligence; Ning, H., Chen, L., Ullah, A., Luo, X., Eds.; Taylor & Francis: Abingdon, UK, 2019; pp. 259–289. [Google Scholar] [CrossRef]
- Subhani, A.R.; Malik, A.S.; Kamil, N.; Saad, M.N.M. Difference in Brain Dynamics during Arithmetic Task Performed in Stress and Control Conditions. In Proceedings of the 2016 IEEE EMBS Conference on Biomedical Engineering and Sciences (IECBES), Kuala Lumpur, Malaysia, 4–8 December 2016; pp. 695–698. [Google Scholar] [CrossRef]
- Subhani, A.R.; Mumtaz, W.; Saad, M.N.M.; Kamel, N.; Malik, A.S. Machine Learning Framework for the Detection of Mental Stress at Multiple Levels. IEEE Access 2017, 5, 13545–13556. [Google Scholar] [CrossRef]
- Al-Shargie, F. Early Detection of Mental Stress Using Advanced Neuroimaging and Artificial Intelligence. arXiv 2019, arXiv:1903.08511. [Google Scholar]
- Alonso, J.F.; Romero, S.; Ballester, M.R.; Antonijoan, R.M.; Mañanas, M.A. Stress Assessment Based on EEG Univariate Features and Functional Connectivity Measures. Physiol. Meas. 2015, 36, 1351–1365. [Google Scholar] [CrossRef]
- Umar Saeed, S.M.; Anwar, S.M.; Majid, M.; Awais, M.; Alnowami, M. Selection of Neural Oscillatory Features for Human Stress Classification with Single Channel EEG Headset. BioMed Res. Int. 2018, 2018, 1049257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Shargie, F.; Tang, T.B.; Kiguchi, M. Assessment of Mental Stress Effects on Prefrontal Cortical Activities Using Canonical Correlation Analysis: An FNIRS-EEG Study. Biomed. Opt. Express 2017, 8, 2583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darzi, A.; Azami, H.; Khosrowabadi, R. Brain functional connectivity changes in long-term mental stress. J. Neurodev. Cogn. 2019, 1, 16–41. [Google Scholar] [CrossRef]
- Zanetti, M.; Faes, L.; De Cecco, M.; Fornaser, A.; Valente, M.; Guandalini, G.; Nollo, G. Assessment of Mental Stress Through the Analysis of Physiological Signals Acquired From Wearable Devices. In Ambient Assisted Living; Leone, A., Caroppo, A., Rescio, G., Diraco, G., Siciliano, P., Eds.; Lecture Notes in Electrical Engineering; Springer International Publishing: Cham, Switzerland, 2019; Volume 544, pp. 243–256. [Google Scholar] [CrossRef]
- Subhani, A.R.; Mumtaz, W.; Kamil, N.; Saad, N.M.; Nandagopal, N.; Malik, A.S. MRMR Based Feature Selection for the Classification of Stress Using EEG. In Proceedings of the 2017 Eleventh International Conference on Sensing Technology (ICST), Sydney, NSW, Australia, 4–6 December 2017; IEEE: Sydney, NSW, Australia, 2017; pp. 1–4. [Google Scholar] [CrossRef]
- Al-Shargie, F.M.; Hassanin, O.; Tariq, U.; Al-Nashash, H. EEG-Based Semantic Vigilance Level Classification Using Directed Connectivity Patterns and Graph Theory Analysis. IEEE Access 2020, 8, 115941–115956. [Google Scholar] [CrossRef]
- Al-Shargie, F.; Tariq, U.; Hassanin, O.; Mir, H.; Babiloni, F.; Al-Nashash, H. Brain Connectivity Analysis Under Semantic Vigilance and Enhanced Mental States. Brain Sci. 2019, 9, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosrowabadi, R. Stress and Perception of Emotional Stimuli: Long-Term Stress Rewiring the Brain. BCN 2018, 9, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Zhang, J. Estimating the Cortex and Autonomic Nervous Activity during a Mental Arithmetic Task. Biomed. Signal Process. Control. 2012, 7, 303–308. [Google Scholar] [CrossRef]
- Knyazev, G.G. EEG Delta Oscillations as a Correlate of Basic Homeostatic and Motivational Processes. Neurosci. Biobehav. Rev. 2012, 36, 677–695. [Google Scholar] [CrossRef]
- Lőrincz, M.L.; Geall, F.; Bao, Y.; Crunelli, V.; Hughes, S.W. ATP-Dependent Infra-Slow (<0.1 Hz) Oscillations in Thalamic Networks. PLoS ONE 2009, 4, e4447. [Google Scholar] [CrossRef] [Green Version]
- Karavaev, A.S.; Kiselev, A.R.; Runnova, A.E.; Zhuravlev, M.O.; Borovkova, E.I.; Prokhorov, M.D.; Ponomarenko, V.I.; Pchelintseva, S.V.; Efremova, T.Y.; Koronovskii, A.A.; et al. Synchronization of Infra-Slow Oscillations of Brain Potentials with Respiration. Chaos 2018, 28, 81102. [Google Scholar] [CrossRef]
- Ahn, J.W.; Ku, Y.; Kim, H.C. A Novel Wearable EEG and ECG Recording System for Stress Assessment. Sensors 2019, 19, 1991. [Google Scholar] [CrossRef] [Green Version]
- Colosio, M.; Shestakova, A.; Nikulin, V.V.; Blagovechtchenski, E.; Klucharev, V. Neural Mechanisms of Cognitive Dissonance (Revised): An EEG Study. J. Neurosci. 2017, 37, 5074–5083. [Google Scholar] [CrossRef] [Green Version]
- Nakao, T.; Miyagi, M.; Hiramoto, R.; Wolff, A.; Gomez-Pilar, J.; Miyatani, M.; Northoff, G. From Neuronal to Psychological Noise—Long-Range Temporal Correlations in EEG Intrinsic Activity Reduce Noise in Internally-Guided Decision Making. NeuroImage 2019, 201, 116015. [Google Scholar] [CrossRef]
- Sugimura, K.; Iwasa, Y.; Kobayashi, R.; Honda, T.; Hashimoto, J.; Kashihara, S.; Zhu, J.; Yamamoto, K.; Kawahara, T.; Anno, M.; et al. Association between Long-Range Temporal Correlations in Intrinsic EEG Activity and Subjective Sense of Identity. Sci. Rep. 2021, 11, 422. [Google Scholar] [CrossRef]
- Monto, S.; Palva, S.; Voipio, J.; Palva, J.M. Very Slow EEG Fluctuations Predict the Dynamics of Stimulus Detection and Oscillation Amplitudes in Humans. J. Neurosci. 2008, 28, 8268–8272. [Google Scholar] [CrossRef] [Green Version]
- Marshall, L.; Mölle, M.; Fehm, H.L.; Born, J. Changes in Direct Current (DC) Potentials and Infra-Slow EEG Oscillations at the Onset of the Luteinizing Hormone (LH) Pulse: DC Shifts, Infra-Slow Oscillations and LH. Eur. J. Neurosci. 2000, 12, 3935–3943. [Google Scholar] [CrossRef]
- Lázár, Z.I.; Dijk, D.-J.; Lázár, A.S. Infraslow Oscillations in Human Sleep Spindle Activity. J. Neurosci. Methods 2019, 316, 22–34. [Google Scholar] [CrossRef]
- Vanhatalo, S.; Palva, J.M.; Holmes, M.D.; Miller, J.W.; Voipio, J.; Kaila, K. Infraslow Oscillations Modulate Excitability and Interictal Epileptic Activity in the Human Cortex during Sleep. Proc. Natl. Acad. Sci. USA 2004, 101, 5053–5057. [Google Scholar] [CrossRef]
- Stroop, J.R. Studies of Interference in Serial Verbal Reactions. J. Exp. Psychol. 1935, 18, 643–662. [Google Scholar] [CrossRef]
- Schneider, G.M.; Jacobs, D.W.; Gevirtz, R.N.; O’Connor, D.T. Cardiovascular Haemodynamic Response to Repeated Mental Stress in Normotensive Subjects at Genetic Risk of Hypertension: Evidence of Enhanced Reactivity, Blunted Adaptation, and Delayed Recovery. J Hum Hypertens 2003, 17, 829–840. [Google Scholar] [CrossRef] [Green Version]
- Medicom MTD: Electroencephalographic studies “Encephalan-EEG”. Available online: Medicom-mtd.com (accessed on 23 June 2023).
- Ifeachor, E.C.; Jervis, B.W. Digital Signal Processing: A Practical Approach; Pearson Education: London, UK, 2002. [Google Scholar]
- Mormann, F.; Lehnertz, K.; David, P.E.; Elger, C. Mean Phase Coherence as a Measure for Phase Synchronization and Its Application to the EEG of Epilepsy Patients. Phys. D Nonlinear Phenom. 2000, 144, 358–369. [Google Scholar] [CrossRef]
- Gabor, D. Theory of Communication. Part 1: The Analysis of Information. J. Inst. Electr. Eng. Part III 1946, 93, 429–441. [Google Scholar] [CrossRef] [Green Version]
- Smirnov, D.A.; Bezruchko, B.P. Estimation of Interaction Strength and Direction from Short and Noisy Time Series. Phys. Rev. E 2003, 68, 46209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, H.B.; Whitney, D.R. On a Test of Whether One of Two Random Variables Is Stochastically Larger than the Other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Knott, V.; Mahoney, C.; Kennedy, S.; Evans, K. EEG Power, Frequency, Asymmetry and Coherence in Male Depression. Psychiatry Res. Neuroimag. 2001, 106, 123–140. [Google Scholar] [CrossRef]
- Tran, Y.; Thuraisingham, R.A.; Wijesuriya, N.; Nguyen, H.T.; Craig, A. Detecting Neural Changes during Stress and Fatigue Effectively: A Comparison of Spectral Analysis and Sample Entropy. In Proceedings of the 2007 3rd International IEEE/EMBS Conference on Neural Engineering, Kohala Coast, HI, USA, 2–5 May 2007; IEEE: Kohala Coast, HI, USA, 2007; pp. 350–353. [Google Scholar] [CrossRef]
- Al-Shargie, F.; Kiguchi, M.; Badruddin, N.; Dass, S.C.; Hani, A.F.M.; Tang, T.B. Mental Stress Assessment Using Simultaneous Measurement of EEG and FNIRS. Biomed. Opt. Express 2016, 7, 3882. [Google Scholar] [CrossRef] [Green Version]
- Al-shargie, F.M.; Tang, T.B.; Badruddin, N.; Kiguchi, M. Mental Stress Quantification Using EEG Signals. In International Conference for Innovation in Biomedical Engineering and Life Sciences; Ibrahim, F., Usman, J., Mohktar, M.S., Ahmad, M.Y., Eds.; IFMBE Proceedings; Springer: Singapore, 2016; Volume 56, pp. 15–19. [Google Scholar] [CrossRef]
- Demerdzieva, A. Eeg Characteristics of Generalized Anxiety Disorder in Childhood. Acta Inform. Medica 2011, 19, 9–15. [Google Scholar]
- Giannakakis, G.; Grigoriadis, D.; Tsiknakis, M. Detection of Stress/Anxiety State from EEG Features during Video Watching. In Proceedings of the 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Milan, Italy, 25-29 August 2015; pp. 6034–6037. [Google Scholar] [CrossRef]
- Minguillon, J.; Lopez-Gordo, M.A.; Pelayo, F. Stress Assessment by Prefrontal Relative Gamma. Front. Comput. Neurosci. 2016, 10. [Google Scholar] [CrossRef] [Green Version]
- Seo, S.; Gil, Y.; Lee, J. The Relation between Affective Style of Stressor on EEG Asymmetry and Stress Scale during Multimodal Task. In Proceedings of the 2008 Third International Conference on Convergence and Hybrid Information Technology, Busan, Republic of Korea, 11–13 November 2008; Volume 1, pp. 461–466. [Google Scholar] [CrossRef]
- Hayashi, T.; Okamoto, E.; Nishimura, H.; Mizuno-Matsumoto, Y.; Ishii, R.; Ukai, S. Beta activities in EEG associated with emotional stress. Int. J. Intell. Comput. Med. Sci. Image Process. 2009, 3, 57–68. [Google Scholar] [CrossRef]
- Lewis, R.S.; Weekes, N.Y.; Wang, T.H. The Effect of a Naturalistic Stressor on Frontal EEG Asymmetry, Stress, and Health. Biol. Psychol. 2007, 75, 239–247. [Google Scholar] [CrossRef]
- Benedek, M.; Schickel, R.J.; Jauk, E.; Fink, A.; Neubauer, A.C. Alpha Power Increases in Right Parietal Cortex Reflects Focused Internal Attention. Neuropsychologia 2014, 56, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Sauseng, P.; Klimesch, W.; Stadler, W.; Schabus, M.; Doppelmayr, M.; Hanslmayr, S.; Gruber, W.R.; Birbaumer, N. A Shift of Visual Spatial Attention Is Selectively Associated with Human EEG Alpha Activity. Eur. J. Neurosci. 2005, 22, 2917–2926. [Google Scholar] [CrossRef]
- Thatcher, R.W.; Krause, P.J.; Hrybyk, M. Cortico-Cortical Associations and EEG Coherence: A Two-Compartmental Model. Electroencephalogr. Clin. Neurophysiol. 1986, 64, 123–143. [Google Scholar] [CrossRef]
- Smets, E.M.; Garssen, B.; Bonke, B.; De Haes, J.C. The Multidimensional Fatigue Inventory (MFI) psychometric qualities of an instrument to assess fatigue. J. Psychosom. Res. 1995, 39, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://humansystems.arc.nasa.gov/groups/TLX/ (accessed on 15 December 2020).
- Kalauzi, A.; Bojić, T.; Rakić, L. Extracting complexity waveforms from one-dimensional signals. Nonlinear Biomed. Phys. 2009, 3, 8. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coefficients | SCWT | MAT |
|---|---|---|
| Intrahemispheric | ||
| Δδ1-coherence | −23 (−40; −6) | −19 (−35; −4) |
| Δδ2-coherence | −21 (−33; −8) | −10 (−20; 1) |
| Δδ-coherence | −7 (−17; 2) | −4 (−11; 2) |
| Δθ-coherence | −2 (−10; 2) | 2 (−3; 7) |
| Interhemispheric | ||
| Δδ1-coherence | −6 (−30; 32) | −9 (−32; 27) |
| Δδ2-coherence | 7 (−26; 57) | 29 (0; 81) |
| Δδ-coherence | 26 (8; 55) | 30 (11; 70) |
| Δθ-coherence | 6 (−3; 16) | 13 (4; 26) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prokhorov, M.D.; Borovkova, E.I.; Hramkov, A.N.; Dubinkina, E.S.; Ponomarenko, V.I.; Ishbulatov, Y.M.; Kurbako, A.V.; Karavaev, A.S. Changes in the Power and Coupling of Infra-Slow Oscillations in the Signals of EEG Leads during Stress-Inducing Cognitive Tasks. Appl. Sci. 2023, 13, 8390. https://doi.org/10.3390/app13148390
Prokhorov MD, Borovkova EI, Hramkov AN, Dubinkina ES, Ponomarenko VI, Ishbulatov YM, Kurbako AV, Karavaev AS. Changes in the Power and Coupling of Infra-Slow Oscillations in the Signals of EEG Leads during Stress-Inducing Cognitive Tasks. Applied Sciences. 2023; 13(14):8390. https://doi.org/10.3390/app13148390
Chicago/Turabian StyleProkhorov, Mikhail D., Ekaterina I. Borovkova, Aleksey N. Hramkov, Elizaveta S. Dubinkina, Vladimir I. Ponomarenko, Yurii M. Ishbulatov, Alexander V. Kurbako, and Anatoly S. Karavaev. 2023. "Changes in the Power and Coupling of Infra-Slow Oscillations in the Signals of EEG Leads during Stress-Inducing Cognitive Tasks" Applied Sciences 13, no. 14: 8390. https://doi.org/10.3390/app13148390