
Antidiabetic and Hypolipidemic Properties of Newly Isolated Wild Lacticaseibacillus paracasei Strains in Mature Adipocytes
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of L. paracasei Strains
2.2. Preparation of L. paracasei Cell-Free Supernatants
2.3. Cell Differentiation and Design of the Experiment
- Two controls—treated with 10% v/v MRS (MRS_C/24 h and MRS_C/48 h), added to the MM for 24 or 48 h, respectively.
- Eight experimental groups—M2.1/24 h, C8/24 h, C15/24 h, P4/24 h, M2.1/48 h, C8/48 h, C15/48 h, and P4/48 h; treated with L. paracasei CFSs of the respective strain, also as a 10% v/v of MM for 24 or 48 h, respectively.
- Two negative controls—composed of growth-arrested preadipocytes kept in BM throughout the whole experiment and served to exclude the percentage of spontaneous adipogenesis, basal lipolysis, and direct glucose uptake caused only by the high concentration of glucose in the medium.
2.4. Cell Viability Assay (MTT Assay)
2.5. Oil Red O-Staining and Neutral Lipid Measurement
2.6. Glucose Concentration in Supernatants and Glucose Consumption Calculation
2.7. Adipolysis Assay
2.8. Gene Expression Analysis with Real-Time PCR
2.9. Statistical Analysis
3. Results
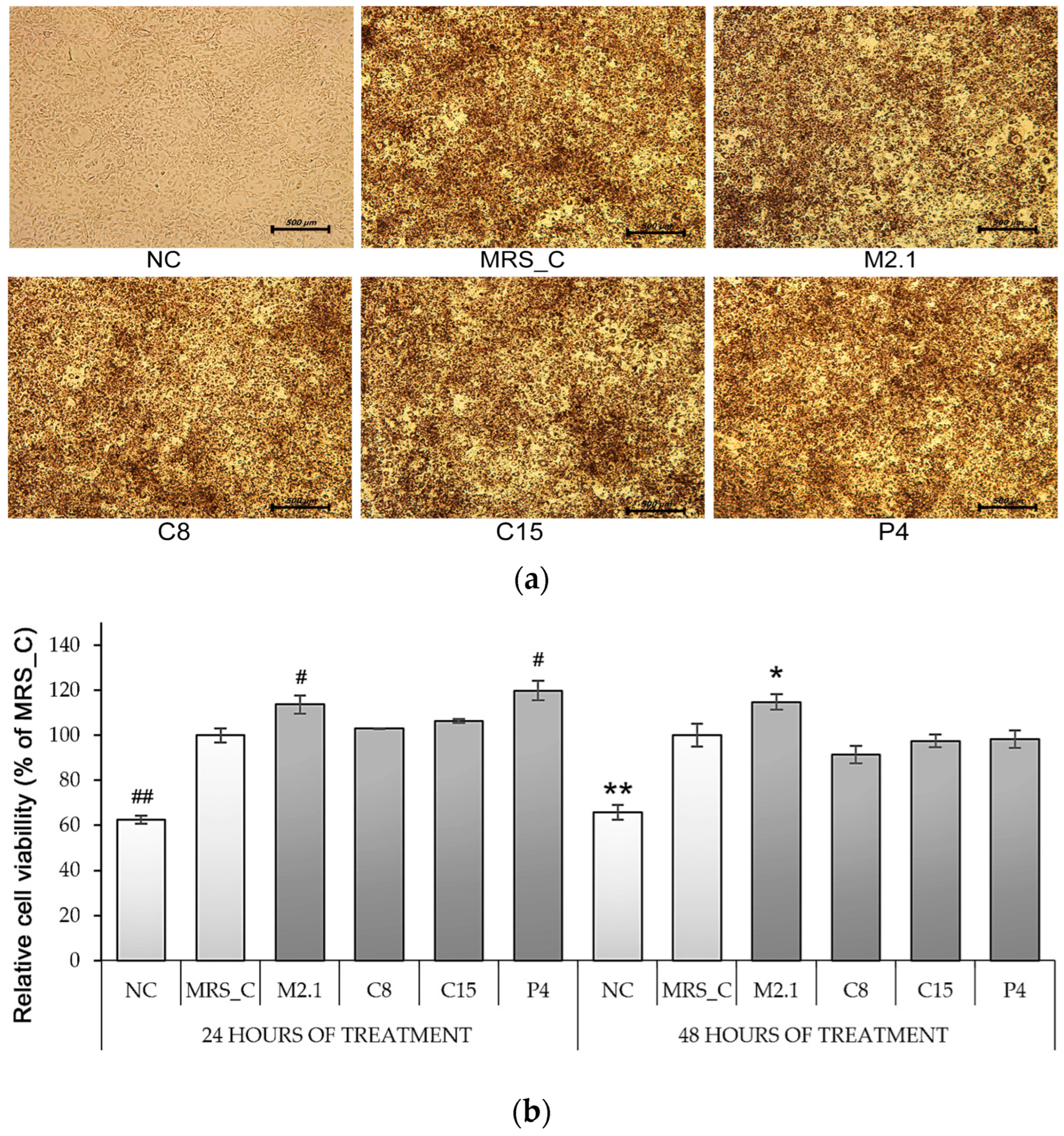
3.1. Effect of L. paracasei CFSs on Cell Viability in 3T3-L1 Adipocytes
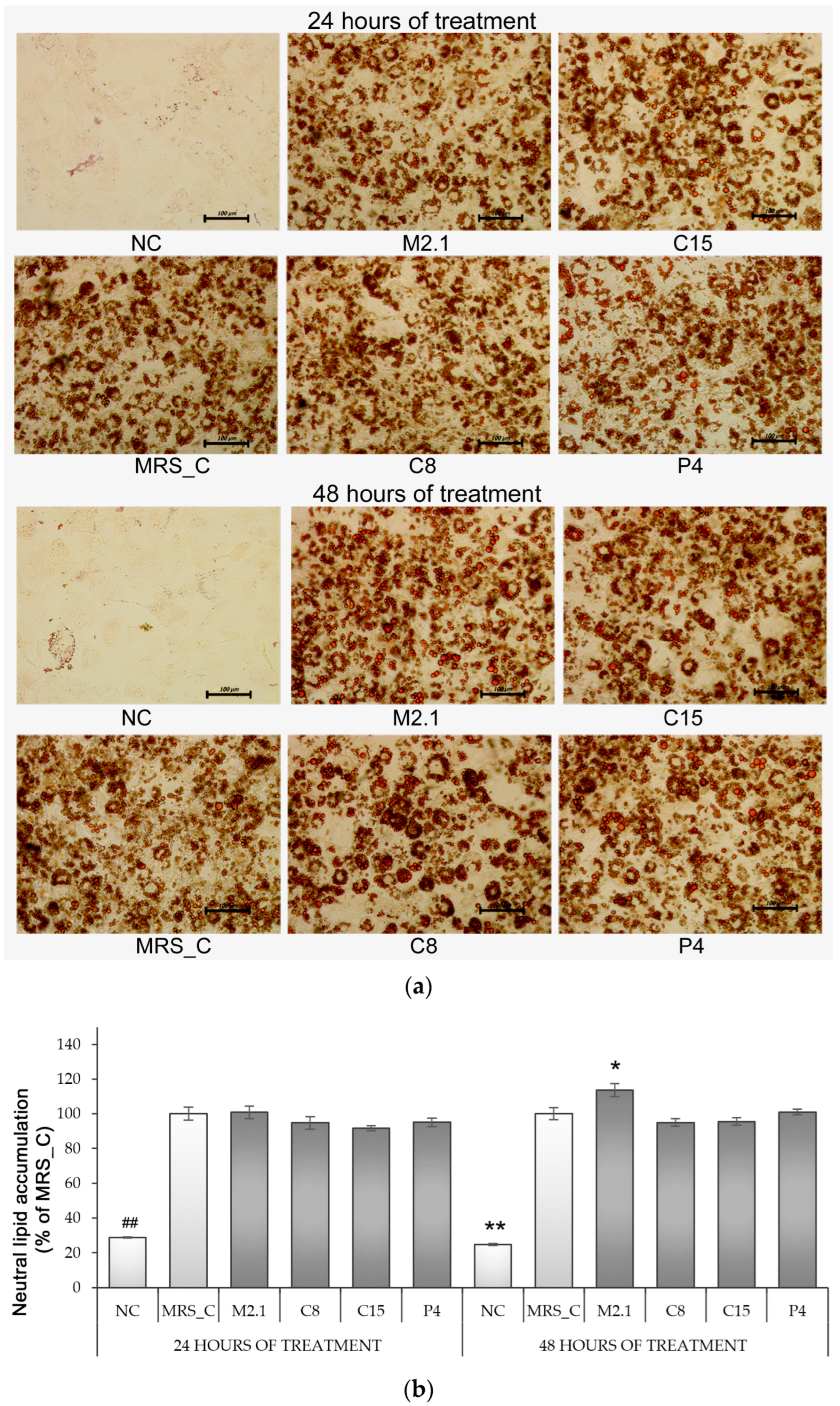
3.2. Effect of L. paracasei CFSs on Neutral Lipid Accumulation in 3T3-L1 Adipocytes
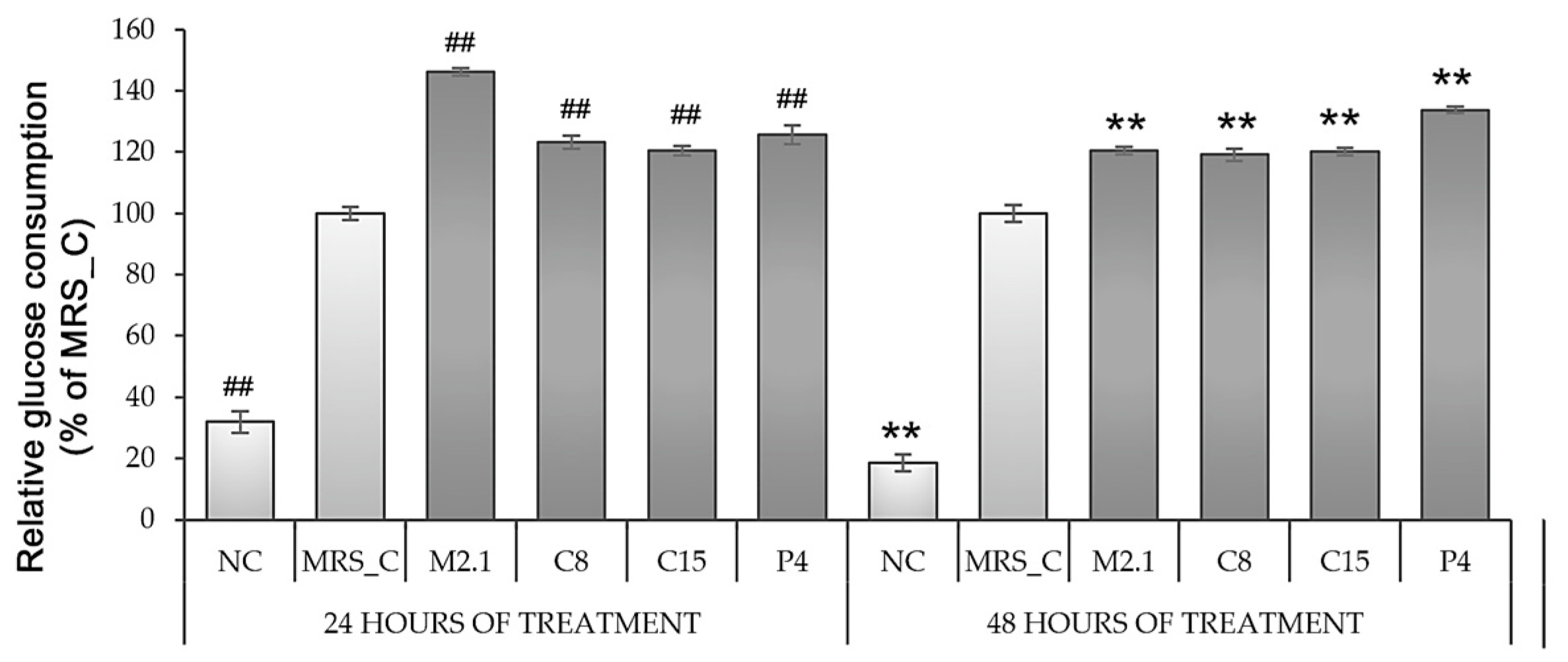
3.3. Effect of L. paracasei CFSs on Glucose Consumption in 3T3-L1 Adipocytes
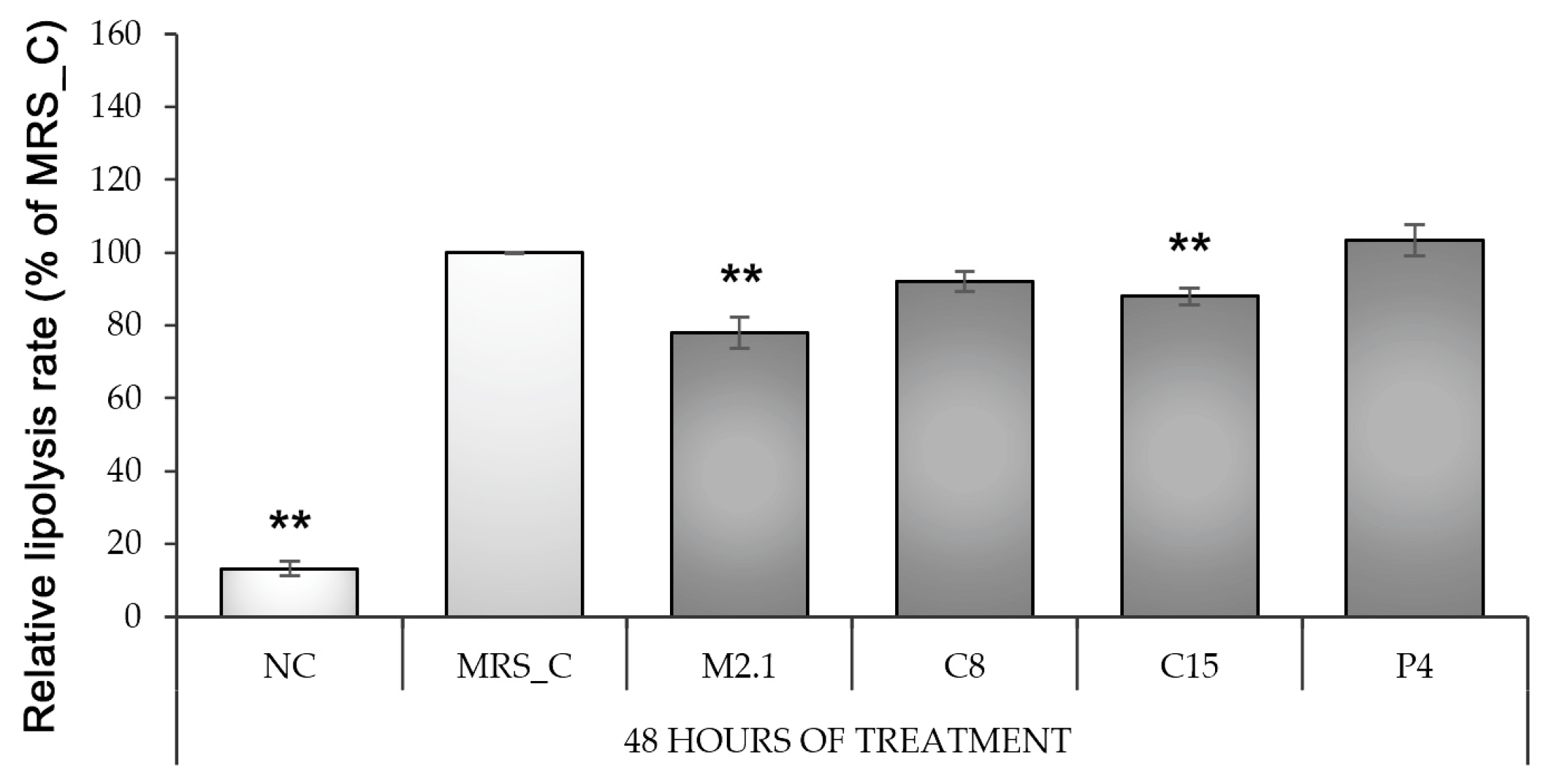
3.4. Effect of L paracasei CFSs on Lipolysis Rate in 3T3-L1 Adipocytes
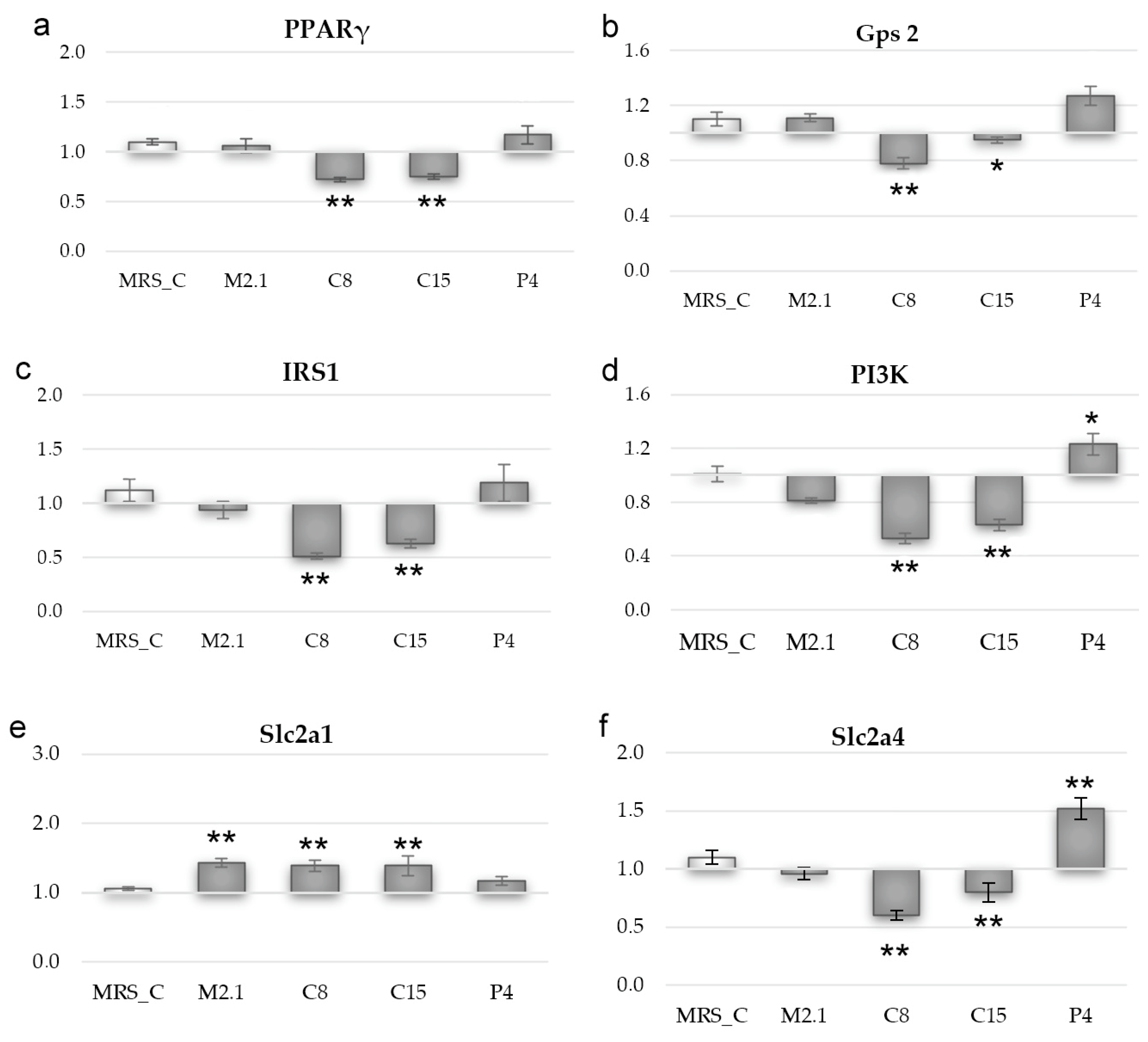
3.5. Effect of L. paracasei CFSs on Relative mRNA Expression in 3T3-L1 Adipocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, J.O.; Wyatt, H.R.; Peters, J.C. Energy Balance and Obesity. Circulation 2012, 126, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Virtue, S.; Vidal-Puig, A. Adipose Tissue Expandability, Lipotoxicity and the Metabolic Syndrome—An Allostatic Perspective. Biochim. Biophys. Acta 2010, 1801, 338–349. [Google Scholar] [CrossRef]
- Rutkowski, J.M.; Stern, J.H.; Scherer, P.E. The Cell Biology of Fat Expansion. J. Cell Biol. 2015, 208, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Cederquist, C.T.; Lentucci, C.; Martinez-Calejman, C.; Hayashi, V.; Orofino, J.; Guertin, D.; Fried, S.K.; Lee, M.J.; Cardamone, M.D.; Perissi, V. Systemic Insulin Sensitivity Is Regulated by GPS2 Inhibition of AKT Ubiquitination and Activation in Adipose Tissue. Mol. Metab. 2016, 6, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Jung, S.R.; Lee, S.Y.; Lee, N.K.; Paik, H.D.; Lim, S.I. Lactobacillus Plantarum Strain Ln4 Attenuates Diet-Induced Obesity, Insulin Resistance, and Changes in Hepatic MRNA Levels Associated with Glucose and Lipid Metabolism. Nutrients 2018, 10, 643. [Google Scholar] [CrossRef]
- Siebenhofer, A.; Winterholer, S.; Jeitler, K.; Horvath, K.; Berghold, A.; Krenn, C.; Semlitsch, T. Long-term Effects of Weight-reducing Drugs in People with Hypertension. Cochrane Database Syst. Rev. 2021, 2021, CD007654. [Google Scholar] [CrossRef]
- Cerdó, T.; García-Santos, J.A.; G Bermúdez, M.; Campoy, C. The Role of Probiotics and Prebiotics in the Prevention and Treatment of Obesity. Nutrients 2019, 11, 635. [Google Scholar] [CrossRef]
- Zsálig, D.; Berta, A.; Tóth, V.; Szabó, Z.; Simon, K.; Figler, M.; Pusztafalvi, H.; Polyák, É. A Review of the Relationship between Gut Microbiome and Obesity. Appl. Sci. 2023, 13, 610. [Google Scholar] [CrossRef]
- Geng, J.; Ni, Q.; Sun, W.; Li, L.; Feng, X. The Links between Gut Microbiota and Obesity and Obesity Related Diseases. Biomed. Pharmacother. 2022, 147, 112678. [Google Scholar] [CrossRef]
- Han, X.; Lee, A.; Huang, S.; Gao, J.; Spence, J.R.; Owyang, C. Lactobacillus Rhamnosus GG Prevents Epithelial Barrier Dysfunction Induced by Interferon-Gamma and Fecal Supernatants from Irritable Bowel Syndrome Patients in Human Intestinal Enteroids and Colonoids. Gut Microbes 2019, 10, 59–76. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and Prebiotics in Intestinal Health and Disease: From Biology to the Clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Raheem, A.; Liang, L.; Zhang, G.; Cui, S. Modulatory Effects of Probiotics During Pathogenic Infections with Emphasis on Immune Regulation. Front. Immunol. 2021, 12, 616713. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhou, L.; Liu, D.; Ge, L.; Li, Y. Probiotic Effect on Helicobacter pylori Attachment and Inhibition of Inflammation in Human Gastric Epithelial Cells. Exp. Ther. Med. 2019, 18, 1551–1562. [Google Scholar] [CrossRef]
- Oh, N.; Lee, J.; Kim, H.; Kwon, M.; Seo, J.; Roh, S. Comparison of Cell-Free Extracts from Three Newly Identified Lactobacillus Plantarum Strains on the Inhibitory Effect of Adipogenic Differentiation and Insulin Resistance in 3T3-L1 Adipocytes. BioMed Res. Int. 2021, 2021, 6676502. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Bron, P.A.; Marco, M.L.; Van Pijkeren, J.P.; O’Connell Motherway, M.; Hill, C.; Pot, B.; Roos, S.; Klaenhammer, T. Identification of Probiotic Effector Molecules: Present State and Future Perspectives. Curr. Opin. Biotechnol. 2018, 49, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.; Zhao, G.; Li, J.; Wang, R.; Ren, F.; Li, Y.; Wang, X. Bifidobacterium Animalis Subsp. Lactis A6 Enhances Fatty Acid-Oxidation of Adipose Tissue to Ameliorate the Development of Obesity in Mice. Nutrients 2022, 14, 598. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, B.; Hu, J.; Nie, S.; Xiong, T.; Xie, M. Intervention of Five Strains of Lactobacillus on Obesity in Mice Induced by High-Fat Diet. J. Funct. Foods 2020, 72, 104078. [Google Scholar] [CrossRef]
- Kim, H.; Lim, J.J.; Shin, H.Y.; Suh, H.J.; Choi, H.S. Lactobacillus Plantarum K8-Based Paraprobiotics Suppress Lipid Accumulation during Adipogenesis by the Regulation of JAK/STAT and AMPK Signaling Pathways. J. Funct. Foods 2021, 87, 104824. [Google Scholar] [CrossRef]
- Castañeda-Márquez, A.C.; Díaz-Benítez, C.E.; Bahena-Roman, M.; Campuzano-Benítez, G.E.; Galván-Portillo, M.; Campuzano-Rincón, J.C.; Lagunas-Martínez, A.; Bermudez-Morales, V.H.; Orbe-Orihuela, Y.C.; Peralta-Romero, J.; et al. Lactobacillus paracasei as a Protective Factor of Obesity Induced by an Unhealthy Diet in Children. Obes. Res. Clin. Pract. 2020, 14, 271–278. [Google Scholar] [CrossRef]
- Guha, D.; Mukherjee, R.; Aich, P. Effects of Two Potential Probiotic Lactobacillus Bacteria on Adipogenesis in Vitro. Life Sci. 2021, 278, 119538. [Google Scholar] [CrossRef]
- Han, K.J.; Lee, N.K.; Yu, H.S.; Park, H.; Paik, H.D. Anti-Adipogenic Effects of the Probiotic Lactiplantibacillus Plantarum KU15117 on 3T3-L1 Adipocytes. Probiotics Antimicrob. Proteins 2022, 14, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.J.; Dong, H.J.; Jeong, H.U.; Ryu, D.W.; Song, S.M.; Kim, Y.R.; Jung, H.H.; Kim, T.H.; Kim, Y.H. Lactobacillus Plantarum LMT1-48 Exerts Anti-Obesity Effect in High-Fat Diet-Induced Obese Mice by Regulating Expression of Lipogenic Genes. Sci. Rep. 2020, 10, 869. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Park, M.H.; Kim, S.H. Selection and Characterization of Probiotic Bacteria Exhibiting Antiadipogenic Potential in 3T3-L1 Preadipocytes. Probiotics Antimicrob. Proteins 2022, 14, 72–86. [Google Scholar] [CrossRef]
- Wu, C.-S.; Lin, C.-C.; Hsieh, F.-C.; Wu, T.-Y.; Fang, A.-H. Antiobesity Effect of Lacticaseibacillus Paracasei LM-141 on High-Fat Diet-Induced Rats through Alleviation of Inflammation and Insulin Resistance. Evid. Based Complement. Alternat. Med. 2023, 2023, 1011591. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Park, M.H.; Kim, B.K.; Kim, S.H. Antiobesity Effect of Novel Probiotic Strains in a Mouse Model of High-Fat Diet–Induced Obesity. Probiotics Antimicrob. Proteins 2021, 13, 1054–1067. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Zheng, H.; Liu, W.-H.; Cheng, R.; Lan, H.; Sun, T.; Zhao, W.; Li, J.; Shen, X.; Li, H.; et al. Lacticaseibacillus Paracasei K56 Attenuates High-Fat Diet-Induced Obesity by Modulating the Gut Microbiota in Mice. Probiotics Antimicrob. Proteins 2022. [CrossRef]
- Gu, Y.; Chen, H.; Li, X.; Li, D.; Sun, Y.; Yang, L.; Ma, Y.; Chan, E.C.Y. Lactobacillus Paracasei IMC 502 Ameliorates Type 2 Diabetes by Mediating Gut Microbiota– SCFA –Hormone/Inflammation Pathway in Mice. J. Sci. Food Agric. 2023, 103, 2949–2959. [Google Scholar] [CrossRef]
- Stefanovic, E.; McAuliffe, O. Comparative Genomic and Metabolic Analysis of Three Lactobacillus paracasei Cheese Isolates Reveals Considerable Genomic Differences in Strains from the Same Niche. BMC Genom. 2018, 19, 869. [Google Scholar] [CrossRef]
- Petrova, P.; Arsov, A.; Tsvetanova, F.; Parvanova-Mancheva, T.; Vasileva, E.; Tsigoriyna, L.; Petrov, K. The Complex Role of Lactic Acid Bacteria in Food Detoxification. Nutrients 2022, 14, 2038. [Google Scholar] [CrossRef]
- Krastanov, A.; Georgiev, M.; Slavchev, A.; Blazheva, D.; Goranov, B.; Ibrahim, S.A. Design and Volatile Compound Profiling of Starter Cultures for Yogurt Preparation. Foods 2023, 12, 379. [Google Scholar] [CrossRef]
- Ivanov, I.; Petrov, K.; Lozanov, V.; Hristov, I.; Wu, Z.; Liu, Z.; Petrova, P. Bioactive Compounds Produced by the Accompanying Microflora in Bulgarian Yoghurt. Processes 2021, 9, 114. [Google Scholar] [CrossRef]
- Dinev, T.; Rusenova, N.; Velichkova, K.; Beev, G. Antimicrobial Potential of Eleven Lacticaseibacillus Paracasei Strains Isolated from Mountain Anthills. Bulg. J. Agric. Sci. 2022, 28, 949–955. [Google Scholar]
- Vachkova, E.; Petrova, V.; Grigorova, N.; Ivanova, Z.; Beev, G. Evaluation of the Anticancer and Probiotic Potential of Autochthonous (Wild) Lacticaseibacillus paracasei Strains from New Ecological Niches as a Possible Additive for Functional Dairy Foods. Foods 2023, 12, 185. [Google Scholar] [CrossRef] [PubMed]
- Ayichew, T.; Belete, A.; Alebachew, T.; Tsehaye, H.; Berhanu, H.; Minwuyelet, A. Bacterial Probiotics Their Importances and Limitations: A Review. J. Nutr. Health Sci. 2017, 4, 1. [Google Scholar] [CrossRef]
- Lee, J.; Park, S.; Oh, N.; Park, J.; Kwon, M.; Seo, J.; Roh, S. Oral Intake of Lactobacillus Plantarum L-14 Extract Alleviates TLR2- and AMPK-Mediated Obesity-Associated Disorders in High-Fat-Diet-Induced Obese C57BL/6J Mice. Cell Prolif. 2021, 54, e13039. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.J.; Rupérez, A.I.; Gomez-Llorente, C.; Gil, A.; Aguilera, C.M. Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review. Int. J. Mol. Sci. 2016, 17, 1040. [Google Scholar] [CrossRef]
- Beev, G.; Michaylova, M.; Dinev, T.; Naydenova, N.; Tzanova, M.; Urshev, Z. ARDRA Analysis on Biodiversity of Lactobacilli Isolated from Bulgarian Raw Buffalo Milk. Acta Microbiol. Bulg. 2021, 37, 22–26. [Google Scholar]
- Roy, D.; Ward, P.; Vincent, D.; Mondou, F. Molecular Identification of Potentially Probiotic Lactobacilli. Curr. Microbiol. 2000, 40, 40–46. [Google Scholar] [CrossRef]
- Walter, J.; Tannock, G.W.; Tilsala-Timisjarvi, A.; Rodtong, S.; Loach, D.M.; Munro, K.; Alatossava, T. Detection and Identification of Gastrointestinal Lactobacillus Species by Using Denaturing Gradient Gel Electrophoresis and Species-Specific PCR Primers. Appl. Environ. Microbiol. 2000, 66, 297–303. [Google Scholar] [CrossRef]
- Ward, L.J.H.; Timmins, M.J. Differentiation of Lactobacillus casei, Lactobacillus paracasei and Lactobacillus rhamnosus by Polymerase Chain Reaction. Lett. Appl. Microbiol. 1999, 29, 90–92. [Google Scholar] [CrossRef]
- Melo, T.A.; Dos Santos, T.F.; De Almeida, M.E.; Junior, L.A.G.F.; Andrade, E.F.; Rezende, R.P.; Marques, L.M.; Romano, C.C. Inhibition of Staphylococcus aureus Biofilm by Lactobacillus Isolated from Fine Cocoa. BMC Microbiol. 2016, 16, 250. [Google Scholar] [CrossRef]
- De Marco, S.; Sichetti, M.; Muradyan, D.; Piccioni, M.; Traina, G.; Pagiotti, R.; Pietrella, D. Probiotic Cell-Free Supernatants Exhibited Anti-Inflammatory and Antioxidant Activity on Human Gut Epithelial Cells and Macrophages Stimulated with LPS. Evid. Based Complement. Alternat. Med. 2018, 105, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Tu, Y.; Xia, H.; Jie, G.; Chen, X.; He, P. Suppression of Free-Radicals and Protection against H2O2-Induced Oxidative Damage in HPF-1 Cell by Oxidized Phenolic Compounds Present in Black Tea. Food Chem. 2007, 105, 1349–1356. [Google Scholar] [CrossRef]
- Park, Y.J.; Liang, J.F.; Ko, K.S.; Kim, S.W.; Yang, V.C. Low Molecular Weight Protamine as an Efficient and Nontoxic Gene Carrier: In Vitro Study. J. Gene Med. 2003, 5, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.T.; Fu, J.; Wang, Y.K.; Desai, R.A.; Chen, C.S. Assaying Stem Cell Mechanobiology on Microfabricated Elastomeric Substrates with Geometrically Modulated Rigidity. Nat. Protoc. 2011, 6, 187–213. [Google Scholar] [CrossRef] [PubMed]
- Diaz, P.A.; Gómez Camargo, D.E.; Ondo-Méndez, A.; Gómez-Alegría, C.J. A Colorimetric Bioassay for Quantitation of Both Basal and Insulin-Induced Glucose Consumption in 3T3-L1 Adipose Cells. Heliyon 2020, 6, e03422. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. QBase Relative Quantification Framework and Software for Management and Automated Analysis of Real-Time Quantitative PCR Data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef]
- Zhai, W.; Xu, C.; Ling, Y.; Liu, S.; Deng, J.; Qi, Y.; Londos, C.; Xu, G. Increased Lipolysis in Adipose Tissues Is Associated with Elevation of Systemic Free Fatty Acids and Insulin Resistance in Perilipin Null Mice. Horm. Metab. Res. Horm. Stoffwechs. Horm. Metab. 2010, 42, 247–253. [Google Scholar] [CrossRef]
- Tumova, J.; Andel, M.; Trnka, J. Excess of Free Fatty Acids as a Cause of Metabolic Dysfunction in Skeletal Muscle. Physiol. Res. 2016, 65, 193–207. [Google Scholar] [CrossRef]
- Taghizad, F.; Kazerani, H.R.; Dehghani, H.; Asoodeh, A.; Yaghubi, D. A Novel Approach towards Obesity: The Use of a Bacterial Product, Gassericin A, in 3T3-L1 Cells. Obes. Res. Clin. Pract. 2021, 15, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Liu, G.; Guo, J.; Su, Z.Q. The PI3K/AKT Pathway in Obesity and Type 2 Diabetes. Int. J. Biol. Sci. 2018, 14, 1483. [Google Scholar] [CrossRef] [PubMed]
- Kahn, B.B.; Charron, M.J.; Lodish, H.F.; Cushman, S.W.; Flier, J.S. Differential Regulation of Two Glucose Transporters in Adipose Cells from Diabetic and Insulin-Treated Diabetic Rats. J. Clin. Investig. 1989, 84, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Chadt, A.; Al-Hasani, H. Glucose Transporters in Adipose Tissue, Liver, and Skeletal Muscle in Metabolic Health and Disease. Pflüg. Arch.-Eur. J. Physiol. 2020, 472, 1273–1298. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Mao, S.; Chen, S.; Zhang, W.; Liu, C. Ppars-Orchestrated Metabolic Homeostasis in the Adipose Tissue. Int. J. Mol. Sci. 2021, 22, 8974. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Takakuwa, R.; Marchand, S.; Dentz, E.; Bornert, J.M.; Messaddeq, N.; Wendling, O.; Mark, M.; Desvergne, B.; Wahli, W.; et al. Peroxisome Proliferator-Activated Receptor Gamma Is Required in Mature White and Brown Adipocytes for Their Survival in the Mouse. Proc. Natl. Acad. Sci. USA 2004, 101, 4543–4547. [Google Scholar] [CrossRef]
- Barilla, S.; Liang, N.; Mileti, E.; Ballaire, R.; Lhomme, M.; Ponnaiah, M.; Lemoine, S.; Soprani, A.; Gautier, J.F.; Amri, E.Z.; et al. Loss of G Protein Pathway Suppressor 2 in Human Adipocytes Triggers Lipid Remodeling by Upregulating ATP Binding Cassette Subfamily G Member. Mol. Metab. 2020, 42, 101066. [Google Scholar] [CrossRef]
- Phua, W.W.T.; Wong, M.X.Y.; Liao, Z.; Tan, N.S. An APPARent Functional Consequence in Skeletal Muscle Physiology via Peroxisome Proliferator-Activated Receptors. Int. J. Mol. Sci. 2018, 19, 1425. [Google Scholar] [CrossRef]
- Bond, S.T.; Moody, S.C.; Liu, Y.; Civelek, M.; Villanueva, C.J.; Gregorevic, P.; Kingwell, B.A.; Hevener, A.L.; Lusis, A.J.; Henstridge, D.C.; et al. The E3 Ligase MARCH5 Is a PPARγ Target Gene That Regulates Mitochondria and Metabolism in Adipocytes. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E293–E304. [Google Scholar] [CrossRef]
- Drareni, K.; Ballaire, R.; Barilla, S.; Mathew, M.J.; Toubal, A.; Fan, R.; Liang, N.; Chollet, C.; Huang, Z.; Kondili, M.; et al. GPS2 Deficiency Triggers Maladaptive White Adipose Tissue Expansion in Obesity via HIF1A Activation. Cell Rep. 2018, 24, 2957–2971.e6. [Google Scholar] [CrossRef]
- English, J.; Orofino, J.; Cederquist, C.T.; Paul, I.; Li, H.; Auwerx, J.; Emili, A.; Belkina, A.; Cardamone, D.; Perissi, V. GPS2-Mediated Regulation of the Adipocyte Secretome Modulates Adipose Tissue Remodeling at the Onset of Diet-Induced Obesity. Mol. Metab. 2023, 69, 101682. [Google Scholar] [CrossRef] [PubMed]
- Serena, B. Role of GPS2 in the Regulation of Adipocyte Fate and Function: A Multi-Omics Approach; Karolinska Institutet: Solna, Sweden, 2020; ISBN 978-91-7831-960-2. [Google Scholar]
- Jackson, R.M.; Griesel, B.A.; Gurley, J.M.; Szweda, L.I.; Olson, A.L. Glucose Availability Controls Adipogenesis in Mouse 3T3-L1 Adipocytes via Up-Regulation of Nicotinamide Metabolism. J. Biol. Chem. 2017, 292, 18556–18564. [Google Scholar] [CrossRef] [PubMed]
- Toubal, A.; Clément, K.; Fan, R.; Ancel, P.; Pelloux, V.; Rouault, C.; Veyrie, N.; Hartemann, A.; Treuter, E.; Venteclef, N. SMRT-GPS2 Corepressor Pathway Dysregulation Coincides with Obesity-Linked Adipocyte Inflammation. J. Clin. Investig. 2012, 123, 362–379. [Google Scholar] [CrossRef]
- Cheng, H.S.; Yip, Y.S.; Lim, E.K.Y.; Wahli, W.; Tan, N.S. PPARs and Tumor Microenvironment: The Emerging Roles of the Metabolic Master Regulators in Tumor Stromal–Epithelial Crosstalk and Carcinogenesis. Cancers 2021, 13, 2153. [Google Scholar] [CrossRef] [PubMed]
- Hartley, A.; Ahmad, I. The Role of PPARγ in Prostate Cancer Development and Progression. Br. J. Cancer 2023, 128, 940–945. [Google Scholar] [CrossRef]
- Eghtedari, A.R.; Vaezi, M.A.; Safizadeh, B.; Ghasempour, G.; Babaheidarian, P.; Salimi, V.; Tavakoli-Yaraki, M. Evaluation of the Expression Pattern and Diagnostic Value of PPARγ in Malignant and Benign Primary Bone Tumors. BMC Musculoskelet. Disord. 2022, 23, 746. [Google Scholar] [CrossRef]
- Szablewski, L. Glucose Transporters as Markers of Diagnosis and Prognosis in Cancer Diseases. Oncol. Rev. 2022, 16, 561. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Full Name | Forward Primer | Reverse Primer | Product Size (bp) |
|---|---|---|---|---|
| PPARγ NM_001127330.2 | Peroxisome proliferator-activated receptor gamma, transcript variant 2 | AGGGCGATCTTGACAGGAAA | CGAAACTGGCACCCTTGAAA | 164 |
| Gps2 NM_001357906.2 | G protein pathway suppressor 2 | ACAAGTGCTTACGACCCGG | GGAAATGCTGATGGGGCTCT | 183 |
| Slc2a1 NM_011400.3 | Solute carrier family 2 (facilitated glucose transporter), member 1 | GGAATCGTCGTTGGCATCCT | TCTTCAGCACACTCTTGGCC | 202 |
| IRS-1 NM_010570.4 | Insulin receptor substrate 1 | TTAGGCAGCAATGAGGGCAA | CGTGAGGTCCTGGTTGTGAA | 212 |
| PI3K NM_001077495.2 | Phosphoinositide-3-kinase regulatory subunit 1 | ATACTTGATGTGGCTGACGC | GTCTCGCTTCCCTCGCAATA | 193 |
| Slc2a4 NM_009204.2 | Solute carrier family 2 (facilitated glucose transporter), member 4 | CGTTGGTCTCGGTGCTCTTA | AGCTCTGCCACAATGAACCA | 220 |
| 36b4 NM_007475.5 | Ribosomal protein, large, P0 | TTATAACCCTGAAGTGCTCGAC | CGCTTGTACCCATTGATGATG | 147 |
| Actb NM_007393.5 | β-actin | CCTCTATGCCAACACAGTGC | GTACTCCTGCTTGCTGATCC | 211 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grigorova, N.; Ivanova, Z.; Vachkova, E.; Petrova, V.; Beev, G. Antidiabetic and Hypolipidemic Properties of Newly Isolated Wild Lacticaseibacillus paracasei Strains in Mature Adipocytes. Appl. Sci. 2023, 13, 6489. https://doi.org/10.3390/app13116489
Grigorova N, Ivanova Z, Vachkova E, Petrova V, Beev G. Antidiabetic and Hypolipidemic Properties of Newly Isolated Wild Lacticaseibacillus paracasei Strains in Mature Adipocytes. Applied Sciences. 2023; 13(11):6489. https://doi.org/10.3390/app13116489
Chicago/Turabian StyleGrigorova, Natalia, Zhenya Ivanova, Ekaterina Vachkova, Valeria Petrova, and Georgi Beev. 2023. "Antidiabetic and Hypolipidemic Properties of Newly Isolated Wild Lacticaseibacillus paracasei Strains in Mature Adipocytes" Applied Sciences 13, no. 11: 6489. https://doi.org/10.3390/app13116489