

Neuroprotective Effect of the Mixture of Gastrodiae elata and Glycyrrhizae uralensis In Vitro
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Preparation of GE and GU Extracts
2.3. Cell Culture
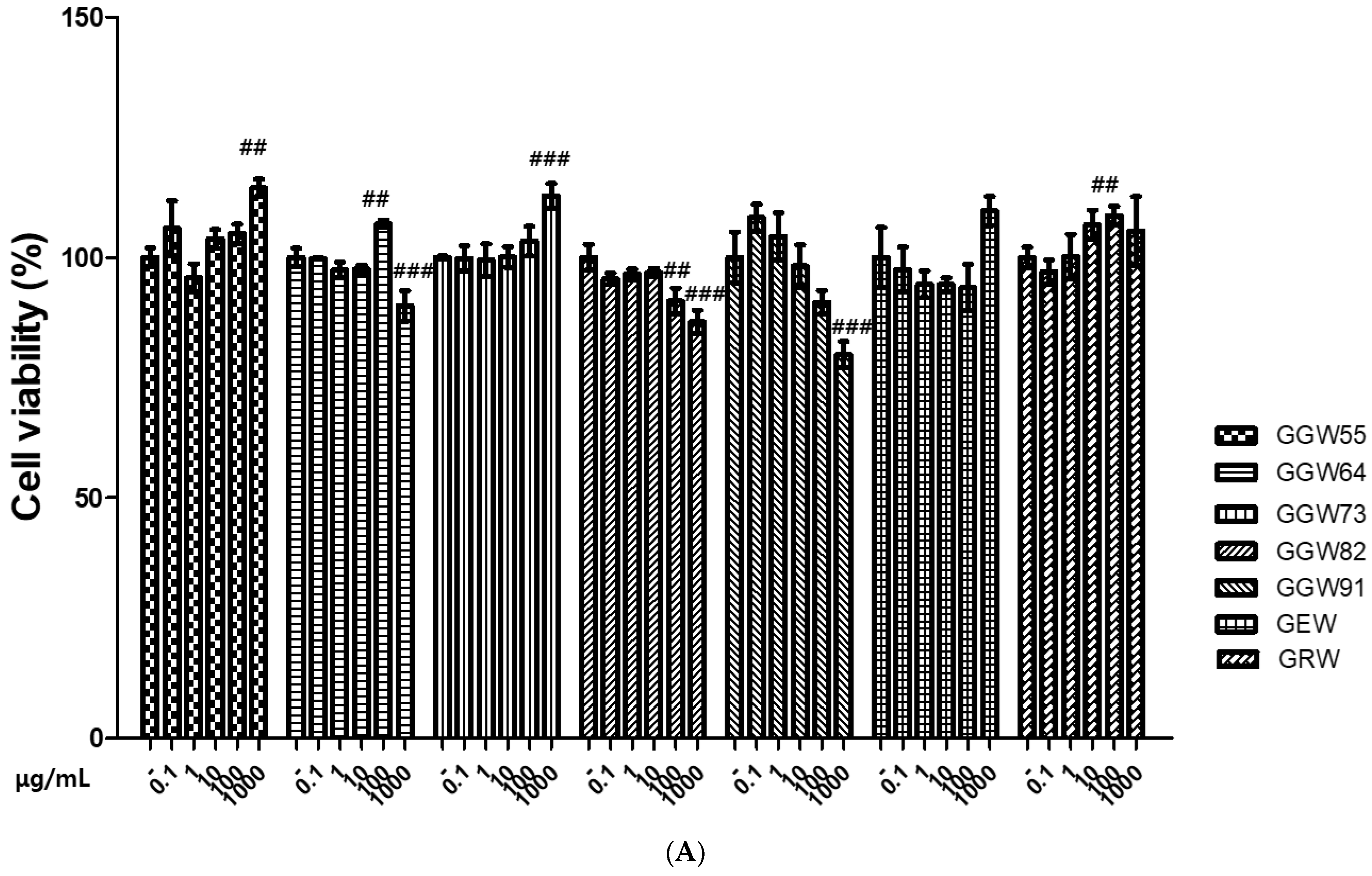
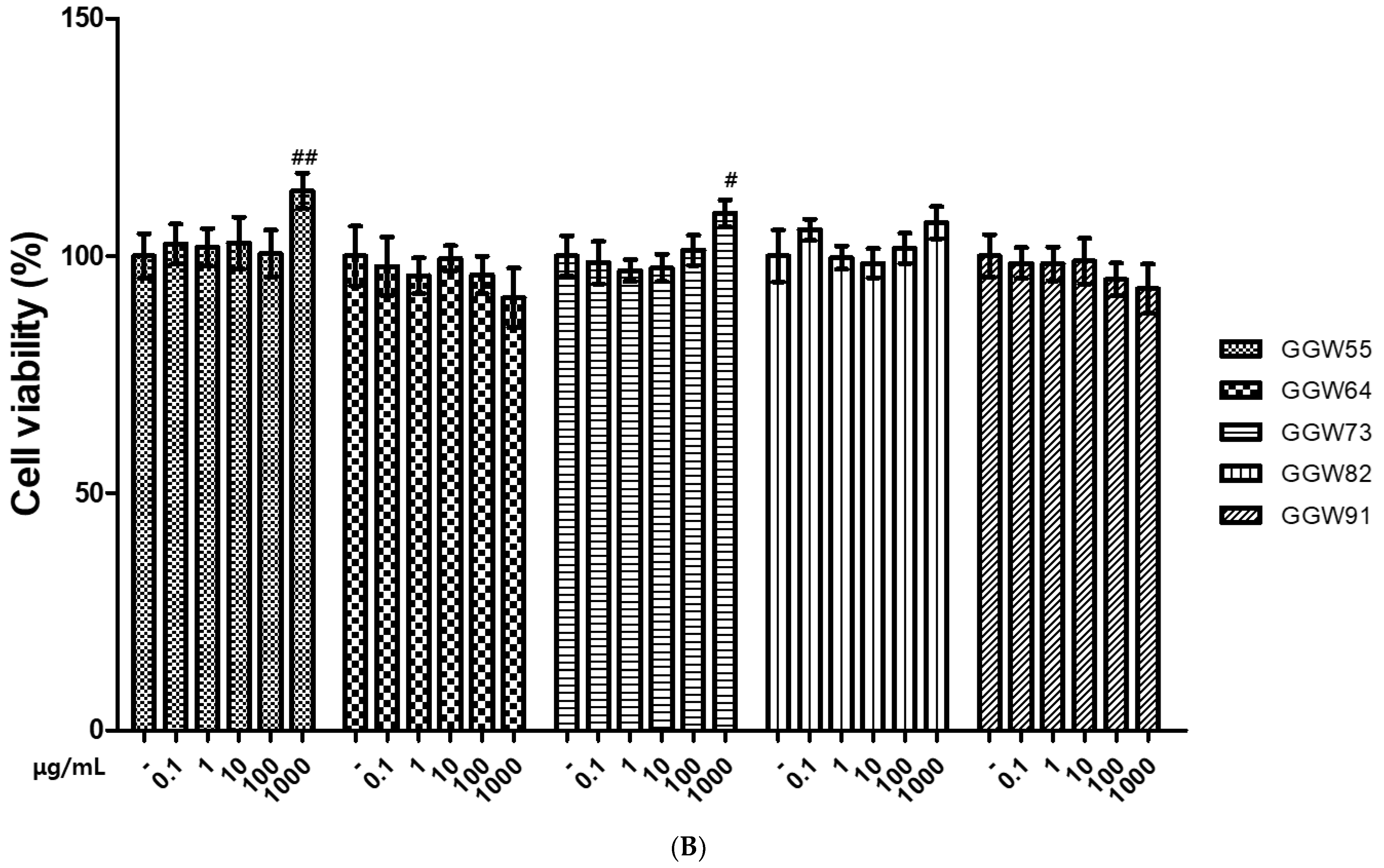
2.4. Cell Viability
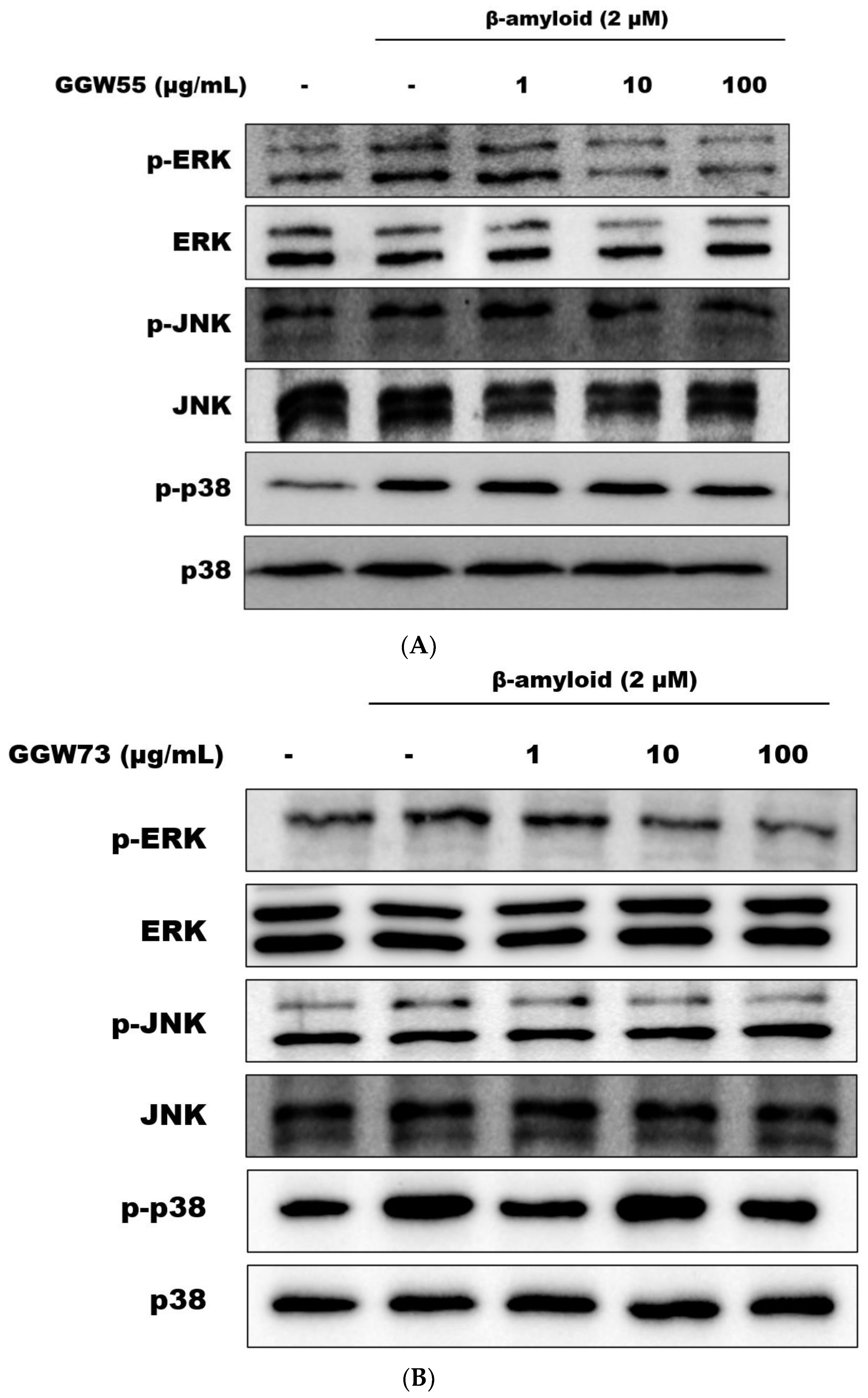
2.5. Western Blot Analysis
2.6. Reactive Oxygen Species (ROS) Analysis
2.7. Quantitative Real Time Polymerase Chain Reaction (qPCR)
2.8. Statistical Analysis
3. Results
3.1. The Cytotoxicity of GGWs on Neuronal Cells
3.2. The Protective Effect of GGW on Aβ-Treated Neuronal Cells
3.3. The Regulatory Effect of GGW on Aβ-Stimulated ACHE Pathway on SH-SY-5Y Cells
3.4. The Regulatory Effect of GGW on Aβ-Stimulatedmitogen Activated Protein Kinases MAPKs) Expression on Neuronal Cells
3.5. The Attenuation Effect of GGW on Aβ-Stimulated Apoptotic Factors on SK-N-SH Cells
3.6. The Regulatory Effect of GGW on H2O2-Stimulated ROS on SK-N-SH Cell and RAW264.7 Cell
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pen, A.E.; Jensen, U.B. Current status of treating neurodegenerative disease with induced pluripotent stem cells. Acta Neurol. Scand. 2016, 135, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Department of Economic and Social Affairs, Population Division. World Population Ageing, United Nations 2019; United Nations: New York, NY, USA, 2019; p. 5. [Google Scholar]
- Gavrilova, S.I.; Alvarez, A. Cerebrolysin in the Therapy of Mild Cognitive Impairment and Dementia Due to Alzheimer’s Disease: 30 Years of Clinical Use. Med. Res. Rev. 2021, 41, 2775–2803. [Google Scholar] [CrossRef] [PubMed]
- Wimo, A.; Jönsson, L.; Bond, J.; Prince, M.; Winblad, B.; International, A.D. Worldwide economic impact of dementia 2010. Alzheimers Dement. 2013, 9, 1–11.e3. [Google Scholar] [CrossRef] [PubMed]
- Vassar, R.; Kandalepas, P.C. The β-secretase enzyme BACE1 as a therapeutic target for Alzheimer ’s disease. Alzheimers Res. Ther. 2011, 3, 20. [Google Scholar] [CrossRef] [PubMed]
- Lauterbach, E.C.; Victoroff, J.; Coburn, K.L.; Shillcutt, S.D.; Doonan, S.M.; Mendez, M.F. Psychopharmacological neuroprotection in neurodegenerative disease: Assessing the preclinical data. J. Neuropsychiatry Clin. Neurosci. 2010, 22, 8–18. [Google Scholar] [CrossRef]
- Sperling, R.A.; Dickerson, B.C.; Pihlajamaki, M.; Vannini, P.; LaViolette, P.S.; Vitolo, O.V.; Hedden, T.; Becker, J.A.; Rentz, D.M.; Selkoe, D.J.; et al. Functional alterations in memory networks in early Alzheimer’s disease. Neuromol. Med. 2010, 12, 27–43. [Google Scholar] [CrossRef] [Green Version]
- Schütt, T.; Helboe, L.; Pedersen, L.Ø.; Waldemar, G.; Berendt, M.; Pedersen, J.T. Dogs with cognitive dysfunction as a spontaneous model for early Alzheimer’s disease: A translational study of neuropathological and inflammatory markers. J. Alzheimers Dis. 2016, 52, 433–449. [Google Scholar] [CrossRef]
- Chen, J.H.; Ke, K.F.; Lu, J.H.; Qiu, Y.H.; Peng, Y.P. Protection of TGF-β 1 against neroinflammation and neurodegeneration in Aβ1-42-induced Alzheimer’s disease model rats. PLoS ONE 2015, 10, e0116549. [Google Scholar]
- Mattson, M.P. Pathways towards and away from Alzheimer ’s disease. Nature 2004, 430, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Findeis, M.A. The role of amyloid beta peptide 42 in Alzheimer ’s disease. Pharmacol. Ther. 2007, 116, 266–286. [Google Scholar] [CrossRef]
- Panza, F.; Seripa, D.; Solfrizzi, V.; Imbimbo, B.P.; Lozupone, M.; Leo, A.; Sardone, R.; Gagliardi, G.; Lofano, L.; Creanza, B.C.; et al. Emerging drugs to reduce abnormal β-amyloid protein in Alzheimer ’s disease patients. Expert Opin. Emerg. Drugs 2016, 21, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Weon, J.B.; Yun, B.R.; Lee, J.; Eom, M.R.; Ko, H.J.; Lee, H.Y.; Park, D.S.; Chung, H.C.; Chung, J.Y.; Ma, C.J. Cognitive-Enhancing Effect of Steamed and Fermented Codonopsis lanceolata: A Behavioral and Biochemical Study. Evid.-Based Complement. Altern. Med. 2014, 2014, 319436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangialasche, F.; Solomon, A.; Winblad, B.; Mecocci, P.; Kivipelto, M. Alzheimer ’s disease: Clinical trials and drug development. Lancet Neurol. 2010, 9, 702–716. [Google Scholar] [CrossRef]
- Hassel, B.; Solyga, V.; Lossius, A. High-affinity choline uptakeandacetylcholine-metabolizing enzymes in CNS white matter. A quantitative study. Neurochem. Int. 2008, 53, 193–198. [Google Scholar] [CrossRef]
- Lee, D.W.; Huh, Y.S.; Kim, K.W. Evidence-based treatment of Alzheimer’s disease. J. Korean Med. Assoc. 2009, 52, 417–425. [Google Scholar] [CrossRef] [Green Version]
- LaFerla, F.M.; Green, K.N.; Oddo, S. Intracellular amyloid-beta in Alzheimer’s disease. Nat. Rev. Neurosci. 2007, 8, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, W.; Tong, Y.; He, G.; Song, W. Control of APP processing and Abeta generation level by BACE1 enzymatic activity and transcription. FASEB J. 2006, 20, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.M.; Lee, B.G.; Park, S.H.; Oh, H.G.; Kang, Y.G.; Kim, O.J.; Kwon, L.S.; Kim, Y.P.; Choi, M.H.; Jeong, Y.S.; et al. Prolonged oral administration of Gastrodia elata extract improves spatial learning and memory of scopolamine-treated rats. Lab. Anim. Res. 2015, 31, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Guan, H.; Cui, C.; Tian, S.; Yang, D.; Wang, X.; Zhang, S.; Wang, L.; Jiang, H. Gastrodin inhibits cell proliferation in vascular smooth muscle cells and attenuates neointima formation in vivo. Int. J. Mol. Med. 2012, 30, 1034–1040. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.L.; Tang, N.Y.; Chiang, S.Y.; Hsieh, C.T.; Lin, J.G. Anticonvulsive and free radical scavenging actions of two herbs, Uncaria rhynchophylla (MIQ) Jack and Gastrodia elata Bl., in kainic acid-treated rats. Life Sci. 1999, 65, 2071–2082. [Google Scholar] [CrossRef]
- Ha, J.H.; Lee, D.U.; Lee, J.T.; Kim, J.S.; Yong, C.S.; Kim, J.A.; Ha, J.S.; Huh, K. 4-Hydroxybenzaldehyde from Gastrodia elata B1. is active in the antioxidation and GABAergic neuromodulation of the rat brain. J. Ethnopharmacol. 2000, 73, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Mishra, M.; Huang, J.; Lee, Y.Y.; Chua, D.S.; Lin, X.; Hu, J.M.; Heese, K. Gastrodia elata modulates amyloid precursor protein cleavage and cognitive functions in mice. Biosci. Trends 2011, 5, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Gao, J.; Peng, M.; Meng, H.; Ma, H.; Cai, P.; Xu, Y.; Zhao, Q.; Si, G. A review on central nervous system effects of gastrodin. Front. Pharmacol. 2018, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Xing, G.H.; Hong, B.; Li, X.M.; Zou, Y.; Zhang, X.J.; Dong, M.X. Gastrodin prevents motor deficits and oxidative stress in the MPTP mouse model of parkinson’s disease: Involvement of ERK1/2-Nrf2 signaling pathway. Life Sci. 2014, 114, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.C.; Lin, Y.H.; Chang, T.M.; Huang, W.Y. Identification of two licorice species, Glycyrrhiza uralensis and Glycyrrhiza glabra, based on separation and identification of their bioactive components. Food Chem. 2012, 132, 2188–2193. [Google Scholar] [CrossRef]
- Mochida, K.; Sakurai, T.; Seki, H.; Yoshida, T.; Takahagi, K.; Sawai, S.; Uchiyama, H.; Muranaka, T.; Saito, K. Draft genome assembly and annotation of Glycyrrhiza uralensis, a medicinal legume. Plant J. 2017, 89, 181–194. [Google Scholar] [CrossRef]
- Asl, M.N.; Hosseinzadeh, H. Review of pharmacological effects of Glycyrrhiza sp. and its bioactive compounds. Phytother. Res. 2008, 22, 709–724. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Nassiri-Asl, M. Pharmacological effects of Glycyrrhiza spp. and its bioactive constituents: Update and review. Phytother. Res. 2015, 29, 1868–1886. [Google Scholar] [CrossRef]
- Nomura, T.; Fukai, T. Phenolic constituents of licorice (Glycyrrhiza species). Fortschr. Chem. Org. Naturst. 1998, 73, 1–158. [Google Scholar]
- Lee, H.K.; Yang, E.J.; Kim, J.Y.; Song, K.S.; Seong, Y.H. Inhibitory effects of Glycyrrhizae radix and its active component, isoliquiritigenin, on Aβ(25-35)-induced neurotoxicity in cultured rat cortical neurons. Arch. Pharm. Res. 2012, 35, 897–904. [Google Scholar] [CrossRef]
- Ahn, J.; Um, M.; Choi, W.; Kim, S.; Ha, T. Protective effects of Glycyrrhiza uralensis Fisch. on the cognitive deficits caused by beta-amyloid peptide 25-35 in young mice. Biogerontology 2006, 7, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Jin, Y.; Kim, I.D.; Lee, J.K. Glycyrrhizin attenuates kainic Acid- induced neuronal cell death in the mouse hippocampus. Exp. Neurobiol. 2013, 22, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.K.; Chun, H.S. Isoliquiritigenin isolated from licorice glycyrrhiza uralensis prevents 6-hydroxydopamine-induced apoptosis in dopaminergic neurons. Biosci. Biotechnol. Biochem. 2012, 76, 536–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parle, M.; Dhingra, D.; Kulkarni, S.K. Memory-strengthening activity of Glycyrrhiza glabra in exteroceptive and interoceptive behavioral models. J. Med. Food 2004, 7, 462–466. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, Y.; Yao, C.; He, Q.; Chen, F.; Yu, H.; Lu, G.; Jiang, N.; Liu, X. The effects of fresh Gastrodia elata Blume on the cognitive deficits induced by chronic restraint stress. Front. Pharmacol. 2022, 13, 890330. [Google Scholar] [CrossRef]
- Zhang, J.S.; Zhou, S.F.; Wang, Q.; Guo, J.N.; Liang, H.M.; Deng, J.B.; He, W.Y. Gastrodin suppresses BACE1 expression under oxidative stress condition via inhibition of the PKR/eIF2alpha pathway in Alzheimer’s disease. Neuroscience 2016, 325, 1–9. [Google Scholar] [CrossRef]
- Maurer, S.V.; Williams, C.L. The cholinergic system modulates memory and hippocampal plasticity via its interactions with non-neuronal cells. Front. Immunol. 2017, 8, 1489. [Google Scholar] [CrossRef] [Green Version]
- Lionetto, M.G.; Caricato, R.; Calisi, A.; Giordano, M.E.; Schettino, T. Acetylcholinesterase as a Biomarker in Environmental and Occupational Medicine: New Insights and Future Perspectives. Biomed. Res. Int. 2013, 2013, 321213. [Google Scholar] [CrossRef] [Green Version]
- Das, K.P.; Barone, S. Neuronal Differentiation in PC12 Cells Is Inhibited by Chlorpyrifos and Its Metabolites: Is Acetylcholinesterase Inhibition the Site of Action? Toxicol. Appl. Pharmacol. 1999, 160, 217–230. [Google Scholar] [CrossRef]
- Lee, J.M.; Ji, E.S.; Kim, T.W.; Kim, C.J.; Shin, M.S.; Lim, B.V.; Chung, Y.R.; Cho, Y.S. Treadmill exercise improves memory function by inhibiting hippocampal apoptosis in pilocarpine-induced epileptic rats. J. Exerc. Rehabil. 2018, 14, 713–723. [Google Scholar] [CrossRef]
- Bekinschtein, P.; Cammarota, M.; Katche, C.; Slipczuk, L.; Rossato, J.I.; Goldin, A.; Izquierdo, I.; Medina, J.H. BDNF is essential to promote persistence of long-term memory storage. Proc. Natl. Acad. Sci. USA 2008, 105, 2711–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnydrig, S.; Korner, L.; Landweer, S.; Ernst, B.; Walker, G.; Otten, U.; Kunz, D. Peripheral lipopolysaccharide administration transiently affects expression of brain-derived neurotrophic factor, corticotropin and proopiomelanocortin in mouse brain. Neurosci. Lett. 2007, 429, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Vassar, R. Targeting the β secretase BACE1 for Alzheimer ’s disease therapy. Lancet Neurol. 2014, 13, 319–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feld, M.; Dimant, B.; Delorenzi, A.; Coso, O.; Romano, A. Phosphorylation of extra-nuclear ERK/MAPK is required for long-term memory consolidation in the crab Chasmagnathus. Behav. Brain Res. 2005, 158, 251–261. [Google Scholar] [CrossRef]
- Isaacs, A.M.; Riva-Cambrin, J.; Yavin, D.; Hockley, A.; Pringsheim, T.M.; Jette, N.; Lethebe, B.C.; Lowerison, M.; Dronyk, J.; Hamilton, M.G. Age-specific Global Epidemiology of Hydrocephalus: Systematic Review, Metanalysis and Global Birth Surveillance. PLoS ONE 2018, 13, e0204926. [Google Scholar] [CrossRef]
- Gholampour, S.; Yamini, B.; Drossler, J.; Frim, D. A new definition for intracranial compliance to evaluate adult hydrocephalus after shunting. Front. Bioeng. Biotechnol. 2022, 10, 900644. [Google Scholar] [CrossRef]
- Peng, S.; Zhang, Y.; Zhang, J.; Wang, H.; Ren, B. ERK in learning and memory: A review of recent research. Int. J. Mol. Sci. 2010, 11, 222–232. [Google Scholar] [CrossRef] [Green Version]
- Tabas, I.; Ron, D. Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat. Cell Biol. 2011, 13, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Hu, C.; Huang, J.; Liu, W.; Lai, W.; Leng, F.; Tang, Q.; Liu, Y.; Wang, Q.; Zhou, M.; et al. ROCK1 Induces Dopaminergic Nerve Cell Apoptosis via the Activation of Drp1- Mediated Aberrant Mitochondrial Fission in Parkinson’s Disease. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Zhang, C.; Qu, S.; Dong, S.; Ma, Q.; Hao, Y.; Liu, Z.; Wang, S.; Zhao, H.; Shi, Y. Chinese Herbal Extracts Exert Neuroprotective Effect in Alzheimer’s Disease Mouse Through the Dopaminergic Synapse/Apoptosis Signaling Pathway. Front. Pharmacol. 2022, 13, 817213. [Google Scholar] [CrossRef]
- Smith, M.A.; Rottkamp, C.A.; Nunomura, A.; Raina, A.K.; Perry, G. Oxidative stress in Alzheimer’s disease. Biochim. Biophys. Acta 2000, 1502, 139–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.D.; Carney, J.M.; Starke-Reed, P.E.; Oliver, C.N.; Stadtman, E.R.; Floyd, R.A.; Markesbery, W.R. Excess brain protein oxidation and enzyme dysfunction in normal aging and in Alzheimer disease. Proc. Natl. Acad. Sci. USA 1991, 88, 10540–10543. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, S.; Oh, Y.; Kim, Y.-M.; Chin, Y.-W.; Cho, J. Inhibition of Oxidative Neurotoxicity and Scopolamine-Induced Memory Impairment by γ-Mangostin: In Vitro and In Vivo Evidence. Oxidative Med. Cell. Longev. 2019, 2019, 3640753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelat, P.B.; Chalimoniuk, M.; Wang, J.H.; Strosznajder, J.B.; Lee, J.C.; Sun, A.Y.; Simonyi, A.; Sun, G.Y. Amyloid beta peptide and NMDA induce ROS from NADPH oxidase and AA release from cytosolic phospholipase A2 in cortical neurons. J. Neurochem. 2008, 106, 45–55. [Google Scholar] [CrossRef]
- Ling, J.; Yu, Q.; Yuan, X.; Liu, W.; Guo, Y.; Duan, Y.; Li, L. Edaravone Improves Intermittent Hypoxia-Induced Cognitive Impairment and Hippocampal Damage in Rats. Biol. Pharm. Bull. 2020, 43, 1196–1201. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| h GAPDH | 5′-TCTAGACGGCAGGTCAGGTCCACC-3′ | 5′-CCACCCATGGCAAATTCCATGGCA-3′ |
| h AChE | 5′-ACAGGTCTGAGCAGCGATCCTGCTTGCT-3′ | 5′-TACGCCTACGTCTTGAACACCGTGCTTC-3 |
| h iNOS | 5′-GCTCTACACCTCCAATGTGACC-3′ | 5′-CTGCCGAGATTTGAGCCTCATG-3′ |
| h Beclin-1 | 5′-CCATGCAGGTGAGCTTCGT-3′ | 5′-GAATCTGCGAGAGACACCATC-3′ |
| h Bcl-2 | 5′-ATGTGTGTGGAGAGCGTCAA-3′ | 5′-GCCGGTTCAGGTACTCAGTC-3′ |
| h BACE-1 | 5′- TCTGTCGGAGGGAGCATGAT-3′ | 5′- GCAAACGAAGGTTGGTGGT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, S.-H.; Park, S.-B.; Yu, D.-Y.; Cho, J.-Y.; Lee, W.-W.; Park, M.-R.; Lee, J.-W.; Jeon, Y.-D. Neuroprotective Effect of the Mixture of Gastrodiae elata and Glycyrrhizae uralensis In Vitro. Appl. Sci. 2023, 13, 190. https://doi.org/10.3390/app13010190
Hwang S-H, Park S-B, Yu D-Y, Cho J-Y, Lee W-W, Park M-R, Lee J-W, Jeon Y-D. Neuroprotective Effect of the Mixture of Gastrodiae elata and Glycyrrhizae uralensis In Vitro. Applied Sciences. 2023; 13(1):190. https://doi.org/10.3390/app13010190
Chicago/Turabian StyleHwang, Su-Ha, Su-Bin Park, Da-Young Yu, Jae-Yeon Cho, Won-Woo Lee, Mi-Ran Park, Jang-Won Lee, and Yong-Deok Jeon. 2023. "Neuroprotective Effect of the Mixture of Gastrodiae elata and Glycyrrhizae uralensis In Vitro" Applied Sciences 13, no. 1: 190. https://doi.org/10.3390/app13010190