
Morphological, Histological and Ultrastructural Changes in Hordeum vulgare (L.) Roots That Have Been Exposed to Negatively Charged Gold Nanoparticles


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterisation of Gold Nanoparticles
2.2. Plant Material
2.3. Culture and Treatment
2.4. Light Microscopy
2.5. Transmission Electron Microscopy (TEM)
3. Results
3.1. Morphological Changes after Treatment with (-) AuNP
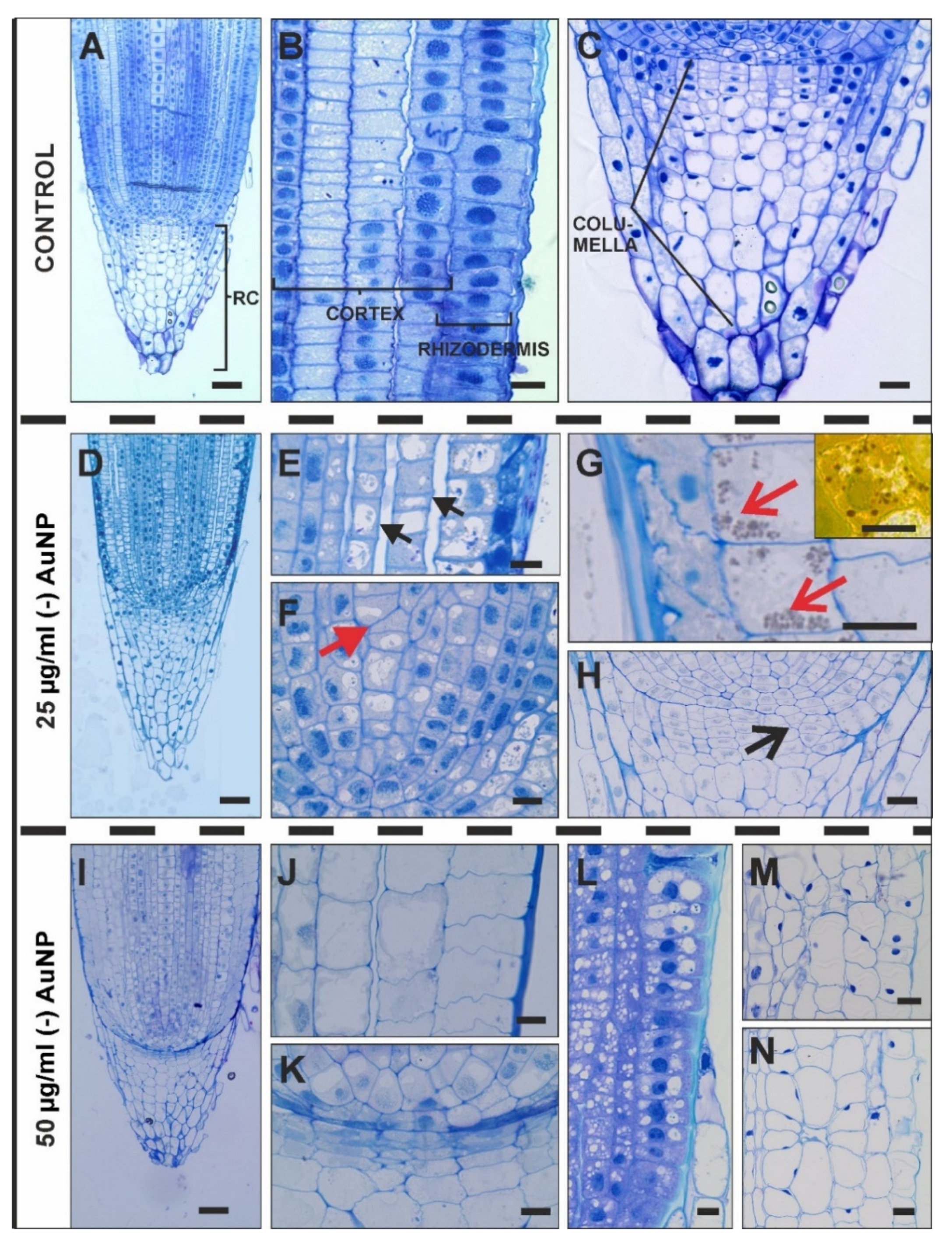
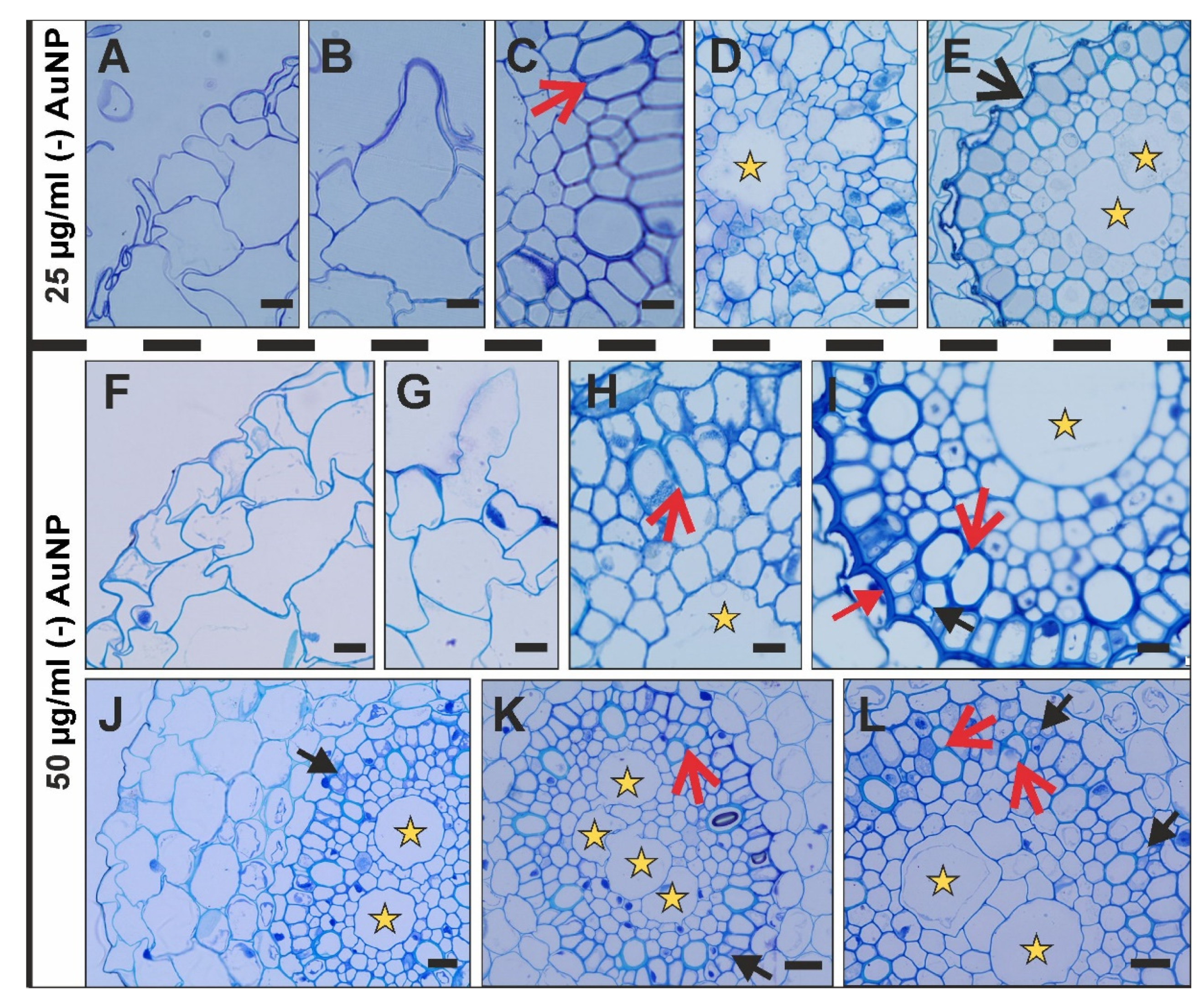
3.2. Histological Changes after Treatment with (-) AuNP
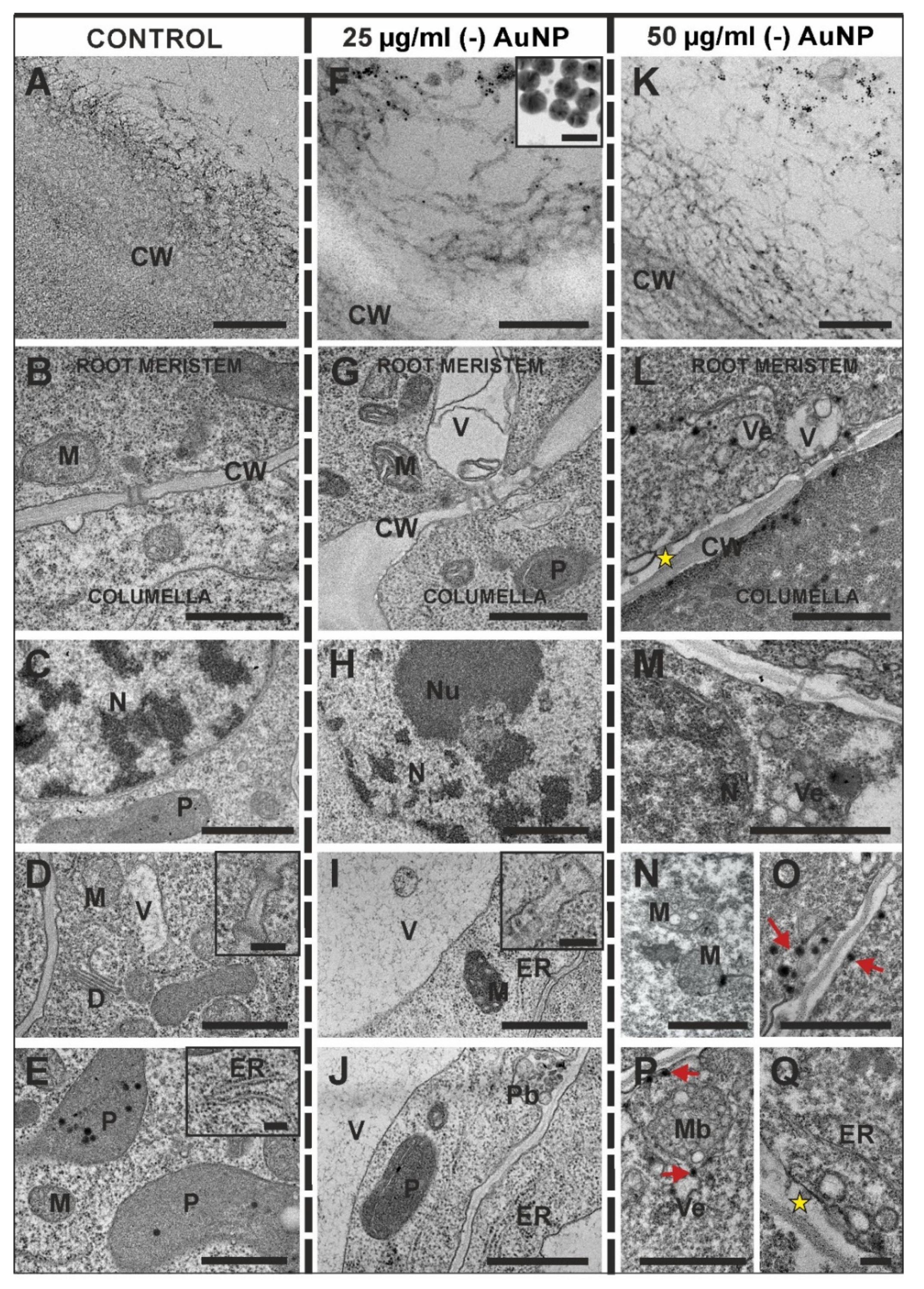
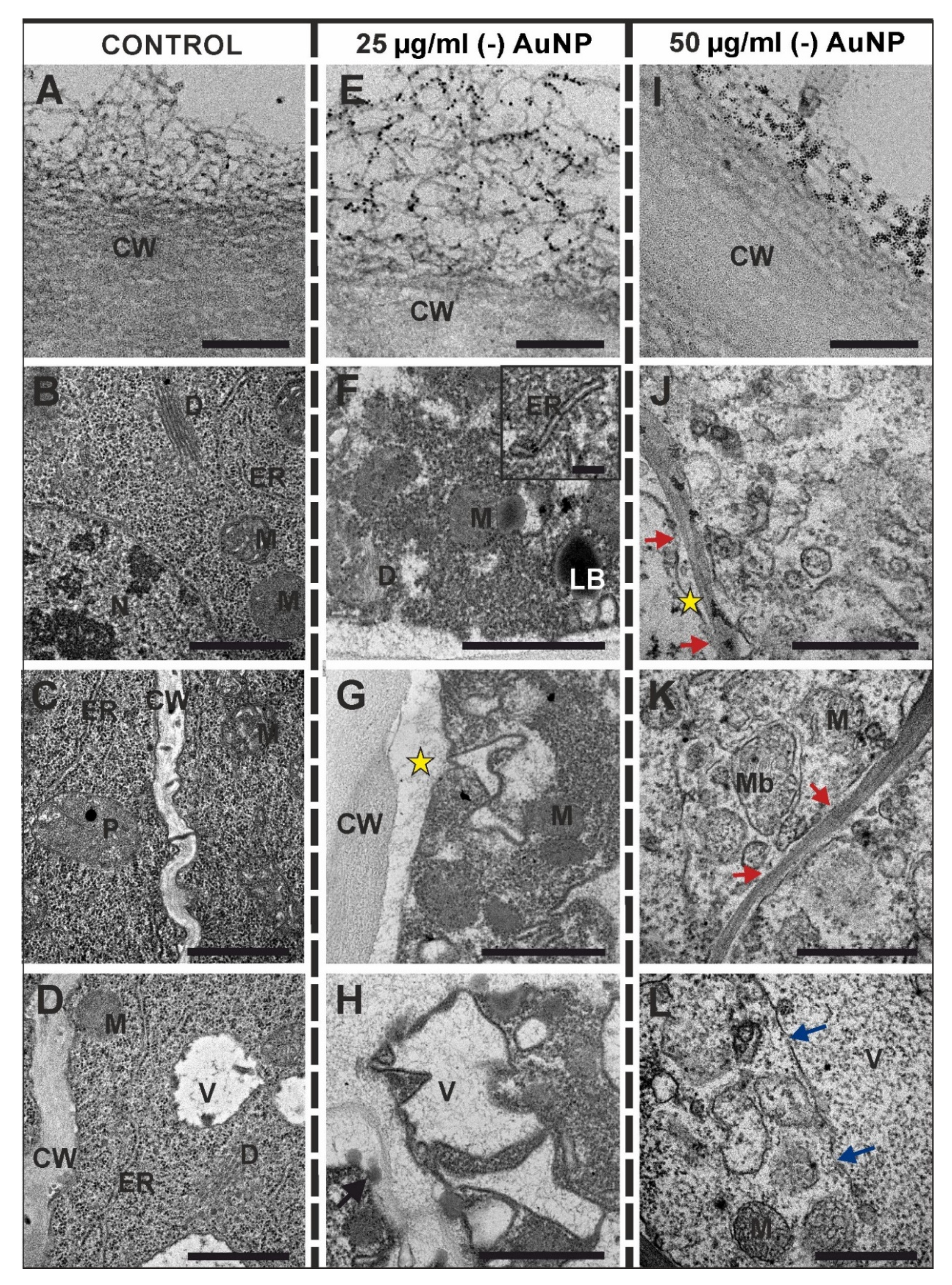
3.3. Ultrastructural Changes after Treatment with (-) AuNP
4. Discussion
4.1. Morphological Malformations of the Roots
4.2. Histological Changes
4.3. Ultrastructural Distortion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Rudramurthy, G.R.; Swamy, M.K. Potential applications of engineered nanoparticles in medicine and biology: An update. J. Biol. Inorg. Chem. 2018, 23, 1185–1204. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Xie, H. Nanoparticles in Daily Life: Applications, Toxicity and Regulations. J. Environ. Pathol. Toxol. Oncol. 2018, 37, 209–230. [Google Scholar] [CrossRef] [PubMed]
- Pestovsky, Y.S.; Martinez-Antonio, A. The Use of Nanoparticles and Nanoformulations in Agriculture. J. Nanosci. Nanotechnol. 2017, 17, 8699–8730. [Google Scholar] [CrossRef]
- Dhiman, S.; Yadav, A.; Debnath, N.; Das, S. Application of Core/Shell Nanoparticles in Smart Farming: A Paradigm Shift for Making the Agriculture Sector More Sustainable. J. Agric. Food Chem. 2021, 69, 3267–3283. [Google Scholar] [CrossRef] [PubMed]
- Usman, M.; Farooq, M.; Wakeel, A.; Nawaz, A.; Cheema, S.A.; Rehman, H.U.; Ashraf, I.; Sanaullah, M. Nanotechnology in agriculture: Current status, challenges and future opportunities. Sci. Total Environ. 2020, 721, 137778. [Google Scholar] [CrossRef]
- Song, U.; Jun, H.; Waldman, B.; Roh, J.; Kim, Y.; Yi, J.; Lee, E.J. Functional analyses of nanoparticle toxicity: A comparative study of the effects of TiO2 and Ag on tomatoes (Lycopersicon esculentum). Ecotoxicol. Environ. Saf. 2013, 93, 60–67. [Google Scholar] [CrossRef]
- Pakrashi, S.; Jain, N.; Dalai, S.; Jayakumar, J.; Chandrasekaran, P.T. In Vivo Genotoxicity Assessment of Titanium Dioxide Nanoparticles by Allium cepa Root Tip Assay at High Exposure Concentrations. PLoS ONE 2014, 9, e87789. [Google Scholar] [CrossRef]
- Ma, R.; Levard, C.; Judy, J.D.; Unrine, J.M.; Durenkamp, M.; Martin, B.; Jefferson, B.; Lowry, G.V. Fate of Zinc Oxide and Silver Nanoparticles in a Pilot Wastewater Treatment Plant and in Processed Biosolids. Environ. Sci. Technol. 2014, 48, 104–112. [Google Scholar] [CrossRef]
- Sharma, R.; Garg, R.; Kumari, A. A Review on Biogenic Synthesis, Applications and Toxicity Aspects of Zinc Oxide Nanoparticles. EXCLI J. 2020, 19, 1325–1340. [Google Scholar]
- Liu, W.T.; Zeb, A.R.; Lian, J.P.; Wu, J.N.; Xiong, H.X.; Tang, J.C.; Zheng, S.N. Interactions of metal-based nanoparticles (MBNPs) and metal-oxide nanoparticles (MONPs) with crop plants: A critical review of research progress and prospects. Environ. Rev. 2020, 28, 294–310. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.; Suskova, S.; Mandzhieva, S.; Tsitsuashvili, V.; Chapligin, V.; Fedorenko, A. Effects of Copper Nanoparticles (CuO NPs) on Crop Plants: A Mini Review. Bionanoscience 2018, 8, 36–42. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. Impacts of Silver Nanoparticles on Plants: A Focus on the Phytotoxicity and Underlying Mechanism. Int. J. Mol. Sci. 2019, 20, 1003. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Xianyu, Y. When nano meets plants: A review on the interplay between nanoparticles and plants. Nano Today 2021, 38, 101143. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; Singh, U.; Adisa, I.O.; Bindraban, P.S.; Elmer, W.H.; Gardea-Torresdey, J.L.; White, J.C. Effects of Manganese Nanoparticle Exposure on Nutrient Acquisition in Wheat (Triticum aestivum L.). Agronomy 2018, 8, 158. [Google Scholar] [CrossRef] [Green Version]
- Zulfiqar, F.; Navarro, M.; Ashraf, M.; Akram, N.A.; Munne-Bosch, S. Nanofertilizer use for sustainable agriculture: Advantages and limitations. Plant Sci. 2019, 289, 110270. [Google Scholar] [CrossRef]
- Dykman, L.; Khlebtsov, N. Gold nanoparticles in biomedical applications: Recent advances and perspectives. Chem. Soc. Rev. 2012, 41, 2256–2282. [Google Scholar] [CrossRef]
- Thompson, D.T. Using gold nanoparticles for catalysis. Nano Today 2007, 2, 40–43. [Google Scholar] [CrossRef]
- Cai, W.B.; Gao, T.; Hong, H.; Sun, J.T. Applications of gold nanoparticles in cancer nanotechnology. Nanotechnol. Sci. Appl. 2008, 1, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Yeh, Y.C.; Creran, B.; Rotello, V.M. Gold nanoparticles: Preparation, properties, and applications in bionanotechnology. Nanoscale 2012, 4, 1871–1880. [Google Scholar] [CrossRef]
- Lafleur, J.P.; Senkbeil, S.; Jensen, T.G.; Kutter, J.P. Gold nanoparticle-based optical microfluidic sensors for analysis of environmental pollutants. Lab Chip 2012, 12, 4651–4656. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Guleria, P.; Kumar, V.; Yadav, S.K. Gold nanoparticle exposure induces growth and yield enhancement in Arabidopsis thaliana. Sci. Total Environ. 2013, 461, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.; Belozerova, I. Influence of Metal Nanoparticles on the Soil Microbial Community and Germination of Lettuce Seeds. Water Air Soil Pollut. 2009, 197, 143–148. [Google Scholar] [CrossRef]
- Arora, S.; Sharma, P.; Kumar, S.; Nayan, R.; Khanna, P.K.; Zaidi, M.G.H. Gold-nanoparticle induced enhancement in growth and seed yield of Brassica juncea. Plant Growth Regul. 2012, 66, 303–310. [Google Scholar] [CrossRef]
- Zhu, Z.J.; Wang, H.H.; Yan, B.; Zheng, H.; Jiang, Y.; Miranda, O.R.; Rotello, V.M.; Xing, B.S.; Vachet, R.W. Effect of Surface Charge on the Uptake and Distribution of Gold Nanoparticles in Four Plant Species. Environ. Sci. Technol. 2012, 46, 12391–12398. [Google Scholar] [CrossRef] [PubMed]
- Milewska-Hendel, A.; Zubko, M.; Stroz, D.; Kurczynska, E.U. Effect of Nanoparticles Surface Charge on the Arabidopsis thaliana (L.) Roots Development and Their Movement into the Root Cells and Protoplasts. Int. J. Mol. Sci. 2019, 20, 1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milewska-Hendel, A.; Witek, W.; Rypien, A.; Zubko, M.; Baranski, R.; Stroz, D.; Kurczynska, E.U. The development of a hairless phenotype in barley roots treated with gold nanoparticles is accompanied by changes in the symplasmic communication. Sci. Rep. 2019, 9, 4724. [Google Scholar] [CrossRef]
- Sabo-Attwood, T.; Unrine, J.M.; Stone, J.W.; Murphy, C.J.; Ghoshroy, S.; Blom, D.; Bertsch, P.M.; Newman, L.A. Uptake, distribution and toxicity of gold nanoparticles in tobacco (Nicotiana xanthi) seedlings. Nanotoxicology 2012, 6, 353–360. [Google Scholar] [CrossRef]
- Pokhrel, L.R.; Dubey, B. Evaluation of developmental responses of two crop plants exposed to silver and zinc oxide nanoparticles. Sci. Total Environ. 2013, 452, 321–332. [Google Scholar] [CrossRef]
- Siddiqi, K.S.; Husen, A. Plant Response to Engineered Metal Oxide Nanoparticles. Nanoscale Res. Lett. 2017, 12, 92. [Google Scholar] [CrossRef] [Green Version]
- Milewska-Hendel, A.; Zubko, M.; Karcz, J.; Stroz, D.; Kurczynska, E. Fate of neutral-charged gold nanoparticles in the roots of the Hordeum vulgare L. cultivar Karat. Sci. Rep. 2017, 7, 3014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurczyńska, E.; Godel-Jędrychowska, K.; Sala, K.; Milewska-Hendel, A. Nanoparticles—Plant Interaction: What We Know, Where We Are? Appl. Sci. 2021, 11, 5473. [Google Scholar] [CrossRef]
- Braszewska-Zalewska, A.J.; Wolny, E.A.; Smialek, L.; Hasterok, R. Tissue-Specific Epigenetic Modifications in Root Apical Meristem Cells of Hordeum vulgare. PLoS ONE 2013, 8, e69204. [Google Scholar] [CrossRef]
- Yin, L.Y.; Cheng, Y.W.; Espinasse, B.; Colman, B.P.; Auffan, M.; Wiesner, M.; Rose, J.; Liu, J.; Bernhardt, E.S. More than the Ions: The Effects of Silver Nanoparticles on Lolium multiflorum. Environ. Sci. Technol. 2011, 45, 2360–2367. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sanchez, S.; Bernales, I.; Cristobal, S. Early response to nanoparticles in the Arabidopsis transcriptome compromises plant defence and root-hair development through salicylic acid signalling. BMC Genom. 2015, 16, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, M.J.; Qu, Q.; Peijnenburg, W.J.G.M.; Li, X.X.; Zhang, M.; Zhang, Z.Y.; Lu, T.; Pan, X.L.; Qian, H.F. Phytotoxic effects of silver nanoparticles and silver ions to Arabidopsis thaliana as revealed by analysis of molecular responses and of metabolic pathways. Sci. Total Environ. 2018, 644, 1070–1079. [Google Scholar] [CrossRef]
- Lee, S.; Chung, H.; Kim, S.; Lee, I. The Genotoxic Effect of ZnO and CuO Nanoparticles on Early Growth of Buckwheat, Fagopyrum esculentum. Water Air Soil Pollut. 2013, 224, 1668. [Google Scholar] [CrossRef]
- Adams, J.; Wright, M.; Wagner, H.; Valiente, J.; Britt, D.; Anderson, A. Cu from dissolution of CuO nanoparticles signals changes in root morphology. Plant Physiol. Biochem. 2017, 110, 108–117. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ebbs, S.E.; Chenc, Y.; Ma, X. Trans-generational impact of cerium oxide nanoparticles on tomato plants. Metallomics 2013, 5, 753–759. [Google Scholar] [CrossRef]
- Burklew, C.E.; Ashlock, J.; Winfrey, W.B.; Zhang, B.H. Effects of Aluminum Oxide Nanoparticles on the Growth, Development, and microRNA Expression of Tobacco (Nicotiana tabacum). PLoS ONE 2012, 7, e34783. [Google Scholar]
- Desoukey, S.F.; Taha, Z.; El-Shabrawi, H.M.; Sabh, A.Z. Impact of zinc oxide nanoparticles on Asparagus officinalis plant. Plant Arch. 2020, 20, 9323–9338. [Google Scholar]
- Venkatachalam, P.; Priyanka, N.; Manikandan, K.; Ganeshbabu, I.; Indiraarulselvi, P.; Geetha, N.; Muralikrishna, K.; Bhattacharya, R.C.; Tiwari, M.; Sharma, N.; et al. Enhanced plant growth promoting role of phycomolecules coated zinc oxide nanoparticles with P supplementation in cotton (Gossypium hirsutum L.). Plant Physiol. Biochem. 2017, 110, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Sotoodehnia-Korani, S.; Iranbakhsh, A.; Ebadi, M.; Majd, A.; Ardebili, Z.O. Selenium nanoparticles induced variations in growth, morphology, anatomy, biochemistry, gene expression, and epigenetic DNA methylation in Capsicum annuum; an in vitro study. Environ. Pollut. 2020, 265, 114727. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.H.; Xing, B.S. Root uptake and phytotoxicity of ZnO nanoparticles. Environ. Sci. Technol. 2008, 42, 5580–5585. [Google Scholar] [CrossRef]
- Sridhar, B.B.M.; Diehl, S.V.; Han, F.X.; Monts, D.L.; Su, Y. Anatomical changes due to uptake and accumulation of Zn and Cd in Indian mustard (Brassica juncea). Environ. Exp. Bot. 2005, 54, 131–141. [Google Scholar] [CrossRef]
- Kouhi, S.M.M.; Lahouti, M.; Ganjeali, A.; Entezari, M.H. Comparative Effects of ZnO Nanoparticles, ZnO Bulk Particles, and Zn2+ on Brassica napus After Long-Term Exposure: Changes in Growth, Biochemical Compounds, Antioxidant Enzyme Activities, and Zn Bioaccumulation. Water Air Soil Pollut. 2015, 226, 364. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.; Fedorenko, A.; Mandzhieva, S.; Sushkova, S.; Lysenko, V.; Duplii, N.; Azarov, A.; Chokheli, V. Destructive Effect of Copper Oxide Nanoparticles on Ultrastructure of Chloroplast, Plastoglobules and Starch Grains in Spring Barley (Hordeum sativum). Int. J. Agric. Biol. 2019, 21, 171–174. [Google Scholar]
- Maeda, K.; Kunieda, T.; Tamura, K.; Hatano, K.; Hara-Nishimura, I.; Shimada, T. Identification of Periplasmic Root-Cap Mucilage in Developing Columella Cells of Arabidopsis thaliana. Plant Cell Physiol. 2019, 60, 1296–1303. [Google Scholar] [CrossRef]
- Kumar, N.; Iyer-Pascuzzi, A.S. Shedding the Last Layer: Mechanisms of Root Cap Cell Release. Plants 2020, 9, 308. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.S.; Mittra, B.; Sharma, S.; Das, T.K.; Babu, C.R. Detection of root mucilage using an anti-fucose antibody. Ann. Bot. 2002, 89, 293–299. [Google Scholar]
- Fox, D.I.; Pichler, T.; Yeh, D.H.; Alcantar, N.A. Removing Heavy Metals in Water: The Interaction of Cactus Mucilage and Arsenate (As (V)). Sci. Technol. 2012, 46, 4553–4559. [Google Scholar] [CrossRef] [PubMed]
- Castiglione, M.R.; Giorgetti, L.; Bellani, L.; Muccifora, S.; Bottega, S.; Spano, C. Root responses to different types of TiO2 nanoparticles and bulk counterpart in plant model system Vicia faba L. Exp. Bot. 2016, 130, 11–21. [Google Scholar] [CrossRef]
- Fedorenko, A.G.; Minkina, T.M.; Chernikova, N.P.; Fedorenko, G.M.; Mandzhieva, S.S.; Rajput, V.D.; Burachevskaya, M.V.; Chaplygin, V.A.; Bauer, T.V.; Sushkova, S.N.; et al. The toxic effect of CuO of different dispersion degrees on the structure and ultrastructure of spring barley cells (Hordeum sativum distichum). Environ. Geochem. Health 2021, 43, 1673–1687. [Google Scholar] [CrossRef] [PubMed]
- Muccifora, S.; Castillo-Michel, H.; Barbieri, F.; Bellani, L.; Castiglione, M.R.; Spano, C.; del Real, A.E.P.; Giorgetti, L.; Tassi, E.L. Synchrotron Radiation Spectroscopy and Transmission Electron Microscopy Techniques to Evaluate TiO2 NPs Incorporation, Speciation, and Impact on Root Cells Ultrastructure of Pisum sativum L. Plants. Nanomaterials 2021, 11, 921. [Google Scholar] [CrossRef] [PubMed]
- Sheteiwy, M.S.; Dong, Q.; An, J.Y.; Song, W.J.; Guan, Y.J.; He, F.; Huang, Y.T.; Hu, J. Regulation of ZnO nanoparticles-induced physiological and molecular changes by seed priming with humic acid in Oryza sativa seedlings. Plant Growth Regul. 2017, 83, 27–41. [Google Scholar] [CrossRef]
- Alkhatib, R.; Alkhatib, B.; Abdo, N.; AL-Eitan, L.; Creamer, R. Physio-biochemical and ultrastructural impact of (Fe3O4) nanoparticles on tobacco. BMC Plant Biol. 2019, 19, 253. [Google Scholar] [CrossRef] [Green Version]
- Kouhi, S.M.M.; Lahouti, M.; Ganjeali, A.; Entezari, M.H. Long-term exposure of rapeseed (Brassica napus L.) to ZnO nanoparticles: Anatomical and ultrastructural responses. Environ. Sci. Pollut. Res. 2015, 22, 10733–10743. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milewska-Hendel, A.; Gepfert, W.; Zubko, M.; Kurczyńska, E. Morphological, Histological and Ultrastructural Changes in Hordeum vulgare (L.) Roots That Have Been Exposed to Negatively Charged Gold Nanoparticles. Appl. Sci. 2022, 12, 3265. https://doi.org/10.3390/app12073265
Milewska-Hendel A, Gepfert W, Zubko M, Kurczyńska E. Morphological, Histological and Ultrastructural Changes in Hordeum vulgare (L.) Roots That Have Been Exposed to Negatively Charged Gold Nanoparticles. Applied Sciences. 2022; 12(7):3265. https://doi.org/10.3390/app12073265
Chicago/Turabian StyleMilewska-Hendel, Anna, Weronika Gepfert, Maciej Zubko, and Ewa Kurczyńska. 2022. "Morphological, Histological and Ultrastructural Changes in Hordeum vulgare (L.) Roots That Have Been Exposed to Negatively Charged Gold Nanoparticles" Applied Sciences 12, no. 7: 3265. https://doi.org/10.3390/app12073265