
Loranthus regularis Ameliorates Neurodegenerative Factors in the Diabetic Rat Retina

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Induction of Diabetes
2.3. Preparation of L. regularis Extract
2.4. Experimental Design
2.5. Retina Isolation
2.6. Determination of Retinal BDNF by ELISA
2.7. Western Blotting
2.8. Glutathione (GSH) and TBAR Assay
2.9. Caspase-3 Assay
2.10. Statistical Analysis
3. Results
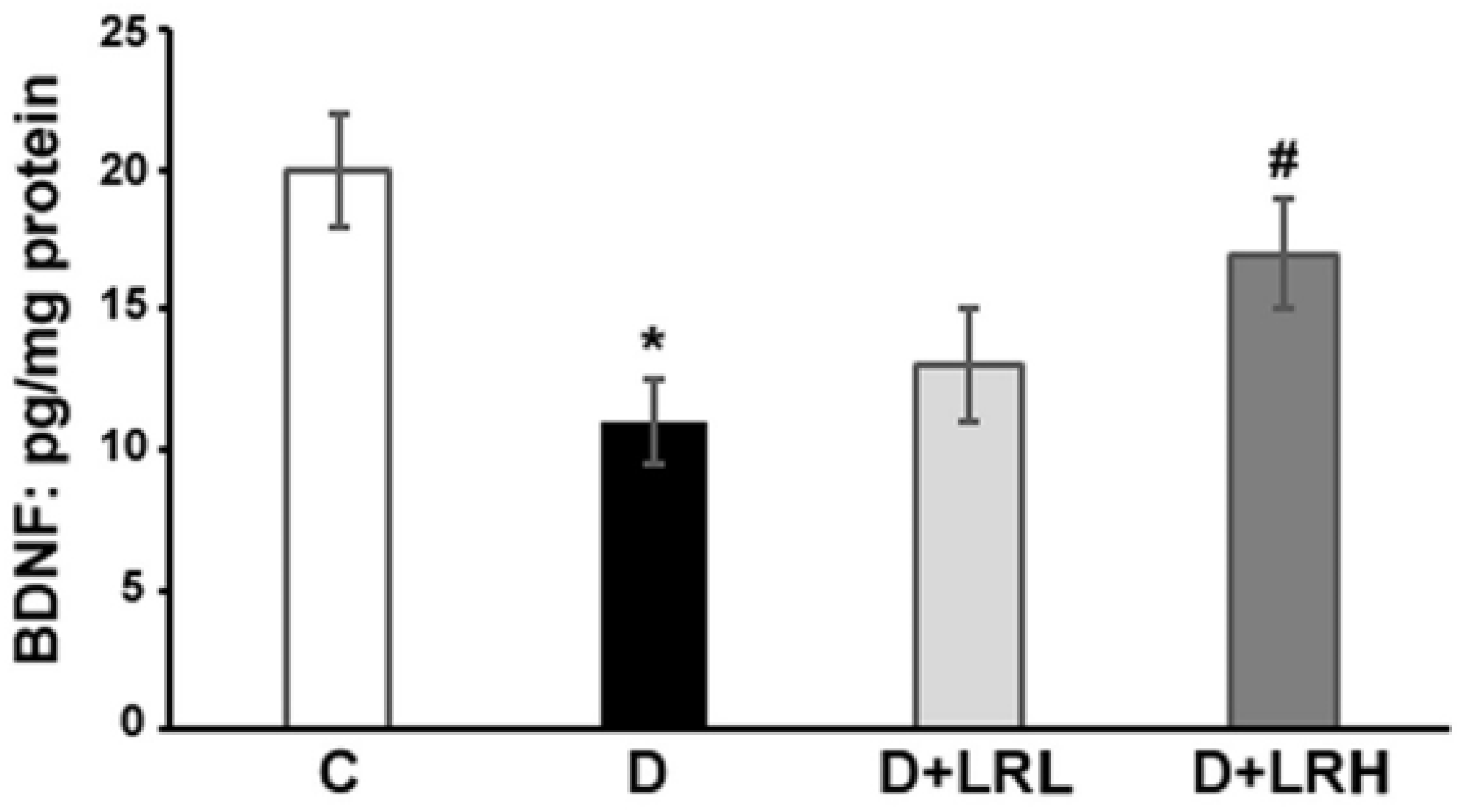
3.1. Effects of L. regularis on Retinal BDNF Levels
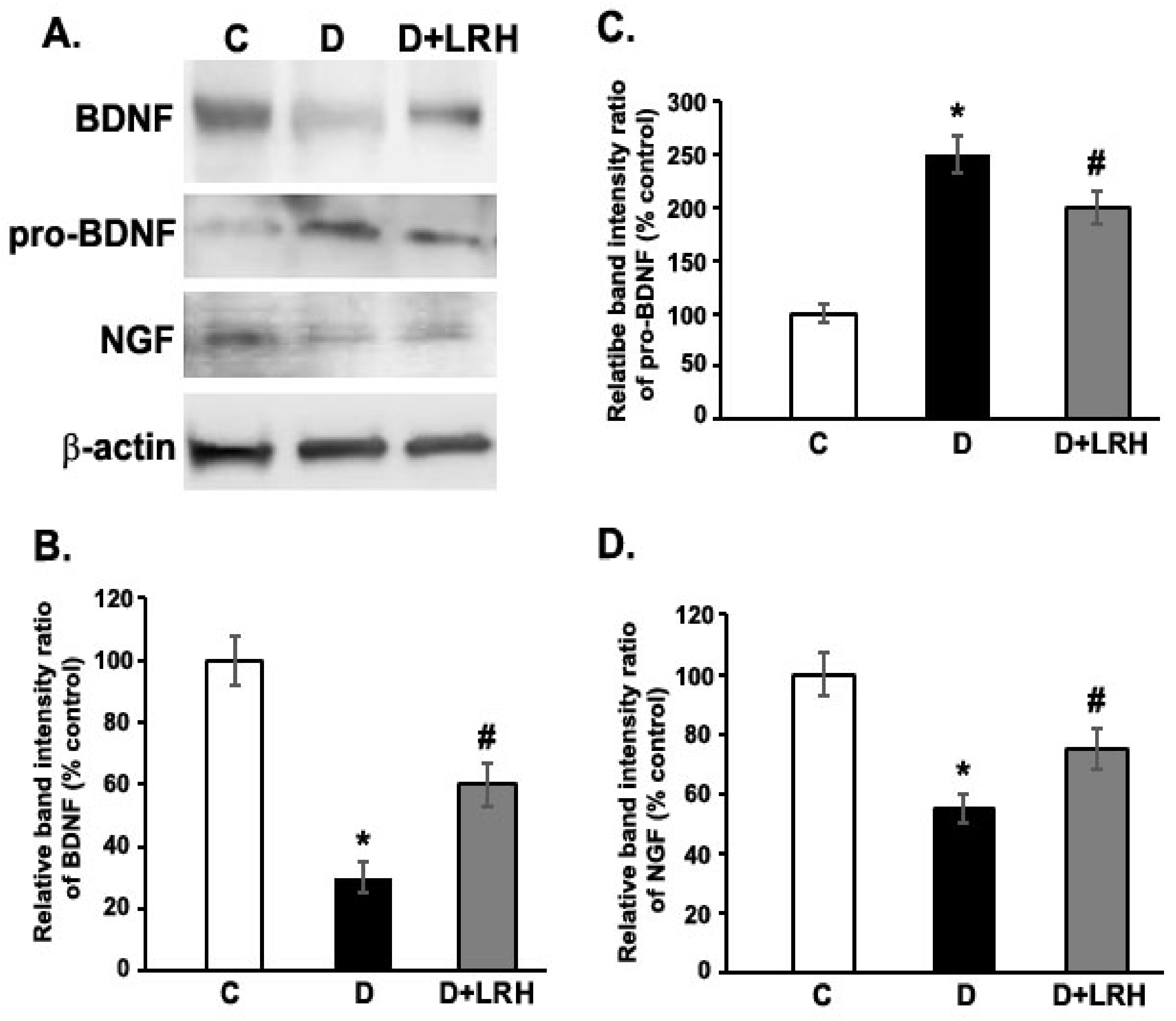
3.2. Effects of L. regularis on Protein Expression of BDNF, proBDNF, and NGF in the Retina of Diabetic Rat
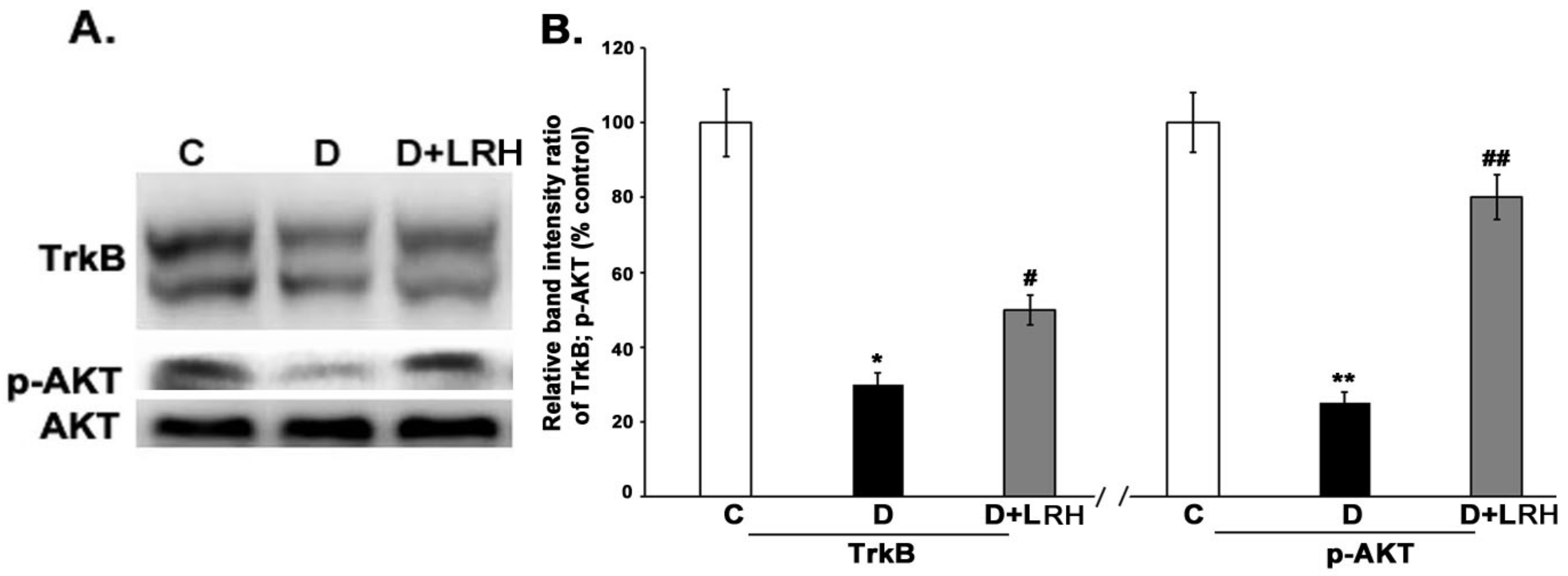
3.3. L. regularis Upregulates TrkB and Phospho-Akt Protein Expression in the Diabetic Retina
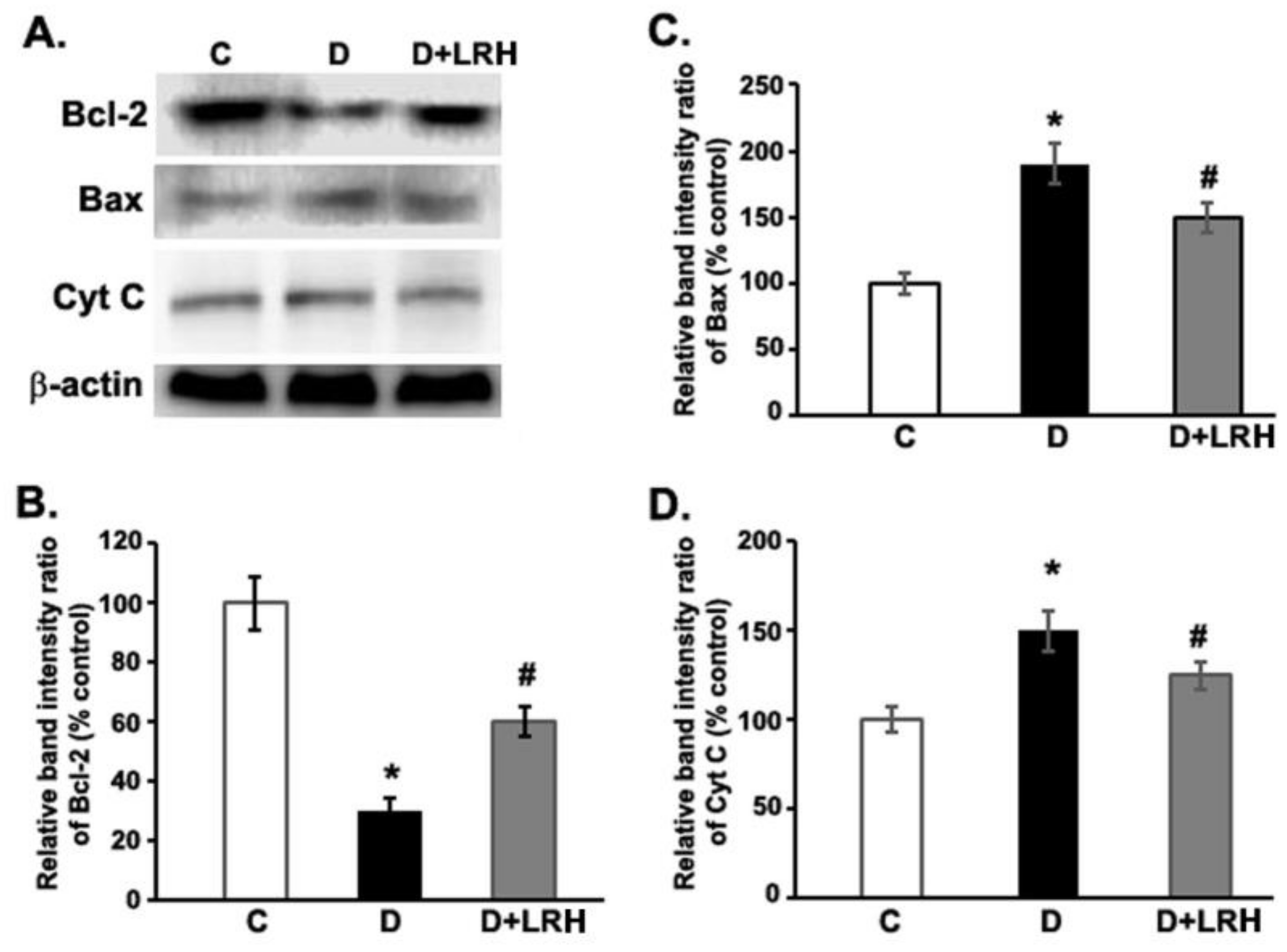
3.4. Effects of L. regularis Treatments on Retinal Bcl-2, Bax, and Cytochrome C Protein Expression in Diabetic Rats
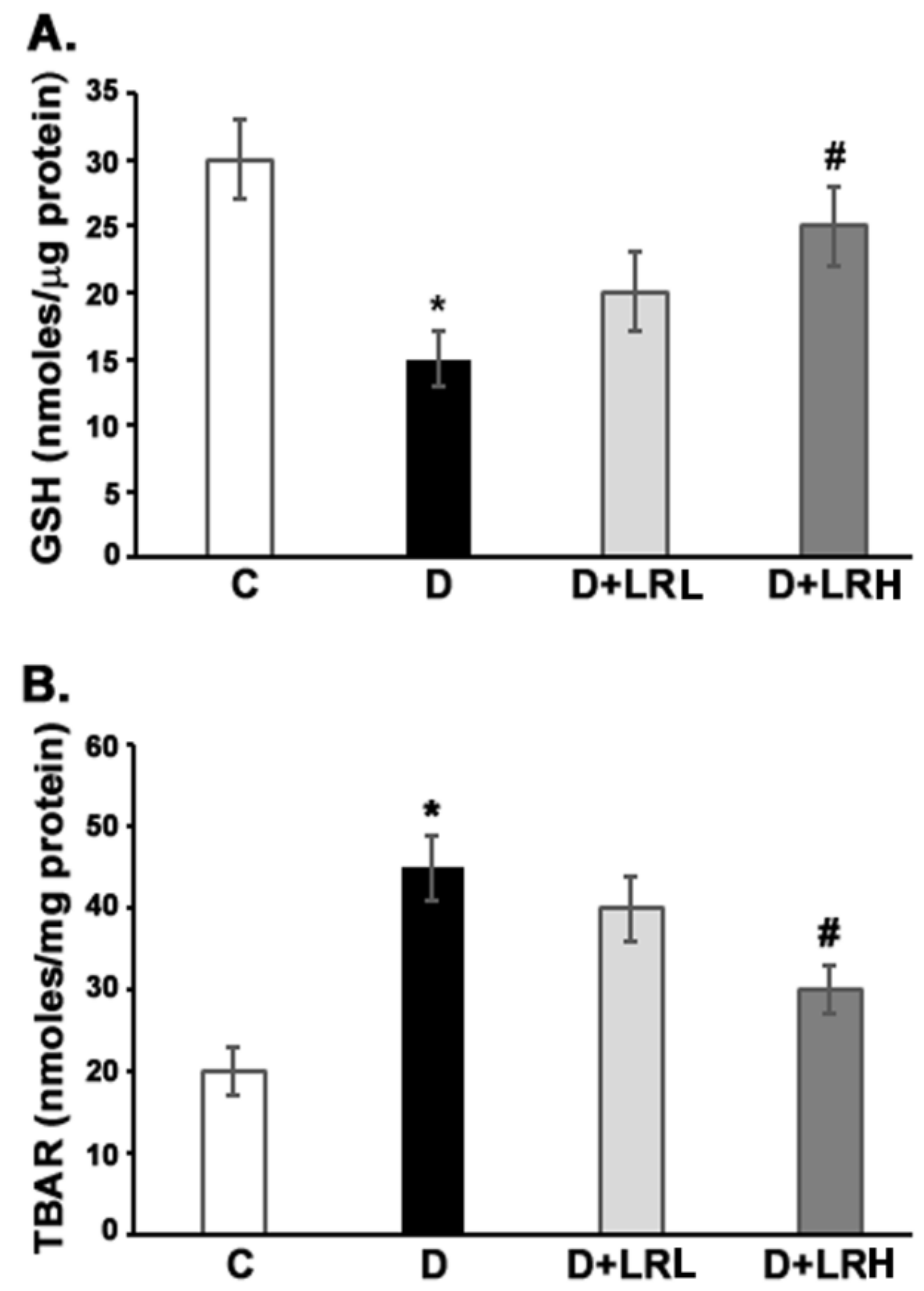
3.5. Effect of L. regularis on Glutathione and TBARs Level
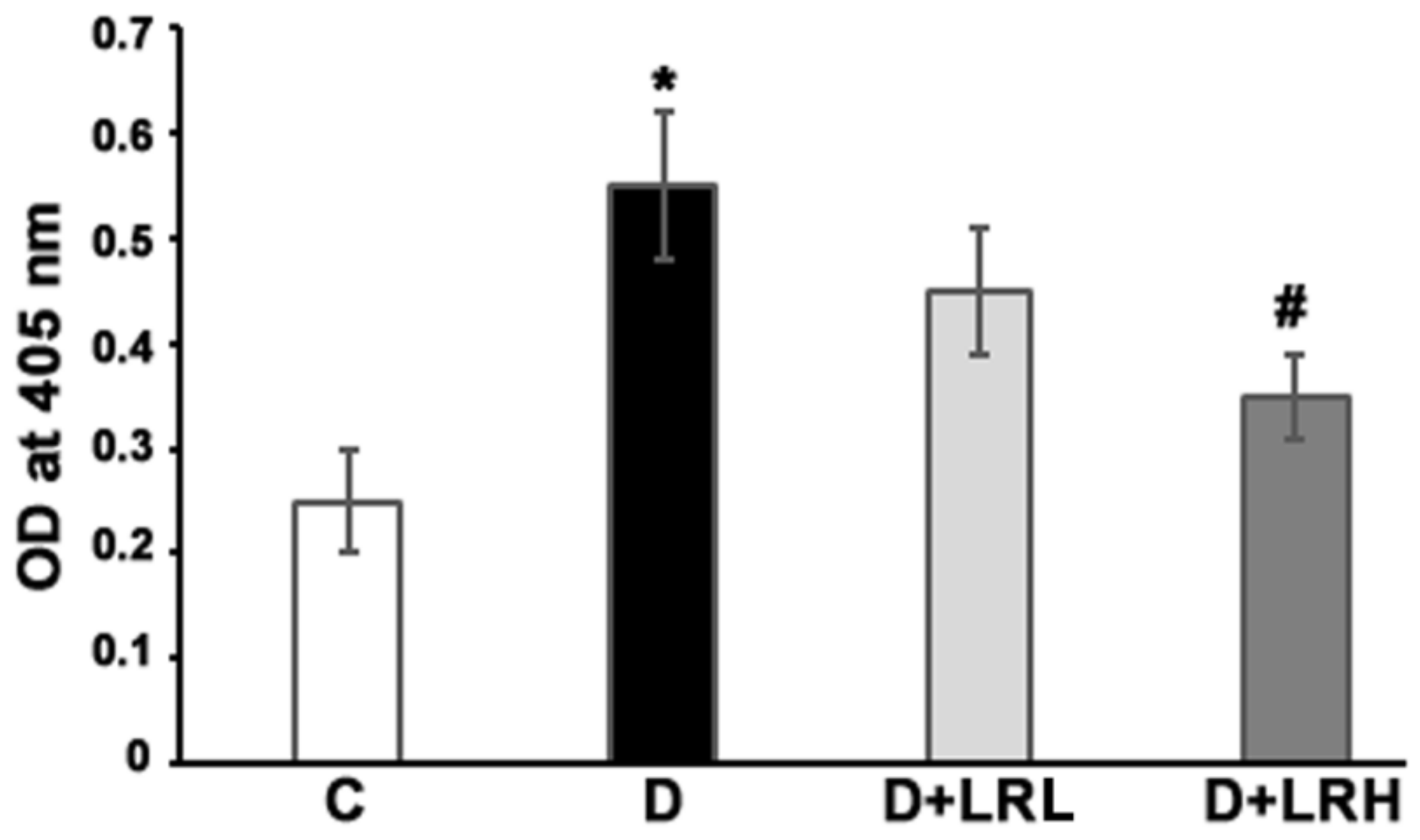
3.6. Caspase-3 Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teo, Z.L.; Tham, Y.-C.; Yu, M.; Chee, M.L.; Rim, T.H.; Cheung, N.; Bikbov, M.M.; Wang, Y.X.; Tang, Y.; Lu, Y.; et al. Global Prevalence of Diabetic Retinopathy and Projection of Burden through 2045: Systematic Review and Meta-analysis. Ophthalmology 2021, 128, 1580–1591. [Google Scholar] [CrossRef]
- Barber, A.J.; Lieth, E.; Khin, S.A.; Antonetti, D.A.; Buchanan, A.G.; Gardner, T.W. Neural apoptosis in the retina during experimental and human diabetes. Early-onset and effect of insulin. J. Clin. Investig. 1998, 102, 783–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bearse, M.A., Jr.; Han, Y.; Schneck, M.E.; Barez, S.; Jacobsen, C.; Adams, A.J. Local multifocal oscillatory potential abnormalities in diabetes and early diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3259–3265. [Google Scholar] [CrossRef] [PubMed]
- Moran, E.P.; Wang, Z.; Chen, J.; Sapieha, P.; Smith, L.E.H.; Ma, J.-X. Neurovascular cross talk in diabetic retinopathy: Pathophysiological roles and therapeutic implications. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H738–H749. [Google Scholar] [CrossRef] [Green Version]
- Ola, M.S.; Nawaz, M.I.; Siddiquei, M.M.; Al-Amro, S.; Abu El-Asrar, A.M. Recent advances in understanding the biochemical and molecular mechanism of diabetic retinopathy. J. Diabetes Complicat. 2012, 26, 56–64. [Google Scholar] [CrossRef]
- Ola, M.S.; Nawaz, M.I.; Khan, H.A.; Alhomida, A.S. Neurodegeneration and neuroprotection in diabetic retinopathy. Int. J. Mol. Sci. 2013, 14, 2559–2572. [Google Scholar] [CrossRef] [Green Version]
- Ola, M.S.; Alhomida, A.S. Neurodegeneration in the diabetic retina and its potential drug targets. Curr. Neuropharmacol. 2014, 12, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Gardner, T.W.; Davila, J.R. The neurovascular unit and the pathophysiologic basis of diabetic retinopathy. Graefe’s Arch. Clin. Exp. Ophthalmol. 2017, 255, 1–6. [Google Scholar] [CrossRef]
- Ola, M.S.; Aleisa, A.M.; Al-Rejaie, S.S.; Abuohashish, H.M.; Parmar, M.Y.; Alhomida, A.S.; Ahmed, M.M. Flavonoid, morin inhibits oxidative stress, inflammation and enhances neurotrophic support in the brain of streptozotocin-induced diabetic rats. Neurol. Sci. 2014, 35, 1003–1008. [Google Scholar] [CrossRef]
- Hackam, A.S. Regulation of neurotrophin expression and activity in the retina. Adv. Exp. Med. Biol. 2008, 613, 343–349. [Google Scholar] [PubMed]
- Cohen-Cory, S.; Kidane, A.H.; Shirkey, N.J.; Marshak, S. Brain derived neurotrophic factor and the development of structural neuronal connectivity. Dev. Neurobiol. 2010, 70, 271–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagahara, A.H.; Tuszynski, M.H. Potential therapeutic uses of BDNF in neurological and psychiatric disorders. Nat. Rev. Drug Discov. 2011, 10, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Tessier-Lavigne, M. Common mechanisms of nerve and blood vessel wiring. Nature 2005, 436, 193–200. [Google Scholar] [CrossRef]
- Suchting, S.; Bicknell, R.; Eichmann, A. Neuronal clues to vascular guidance. Exp. Cell Res. 2006, 312, 668–675. [Google Scholar] [CrossRef]
- Rohrer, B.; Korenbrot, J.I.; LaVail, M.M.; Reichardt, L.; Xu, B. Role of neurotrophin receptor TrkB in the maturation of rod photoreceptors and establishment of synaptic transmission to the inner retina. J. Neurosci. 1999, 19, 8919–8930. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.Q.; Qi, D.S.; Yu, H.L.; Liu, J.; Yang, L.H.; Wu, X.X. Quercetin attenuates cell apoptosis in focal cerebral ischemia rat brain via activation of BDNF-TrkB-PI3K/Akt signaling pathway. Neurochem. Res. 2012, 37, 2777–2786. [Google Scholar] [CrossRef]
- Ola, M.S.; Nawaz, M.I.; El-Asrar, A.A.; Abouammoh, M.; Alhomida, A.S. Reduced levels of brain derived neurotrophic factor (BDNF) in the serum of diabetic retinopathy patients and in the retina of diabetic rats. Cell. Mol. Neurobiol. 2013, 33, 359–367. [Google Scholar] [CrossRef]
- Ola, M.S.; Ahmed, M.M.; Ahmad, R.; Abuohashish, H.M.; Al-Rejaie, S.S.; Alhomida, A.S. Neuroprotective effects of rutin in streptozotocin-induced diabetic rat retina. J. Mol. Neurosci. 2015, 56, 440–448. [Google Scholar] [CrossRef]
- Krabbe, K.S.; Nielsen, A.R.; Krogh-Madsen, R.; Plomgaard, P.; Rasmussen, P.; Erikstrup, C.; Fischer, C.P.; Lindegaard, B.; Petersen, A.M.; Taudorf, S.; et al. Brain derived neurotrophic factor (BDNF) and type 2 diabetes. Diabetologia 2007, 50, 431–438. [Google Scholar] [CrossRef]
- Podesta, F.; Romeo, G.; Liu, W.H.; Krajewski, S.; Reed, J.C.; Gerhardinger, C.; Lorenzi, M. Bax is increased in the retina of diabetic subjects and is associated with pericyte apoptosis in vivo and in vitro. Am. J. Pathol. 2000, 156, 1025–1032. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.; Gupta, S.K.; Srinivasan, B.P.; Nag, T.C.; Srivastava, S.; Saxena, R.; Jha, K.A. Hesperetin rescues retinal oxidative stress, neuroinflammation and apoptosis in diabetic rats. Microvasc. Res. 2013, 87, 65–74. [Google Scholar] [CrossRef]
- Ola, M.S.; Ahmed, M.M.; Shams, S.; Al-Rejaie, S.S. Neuroprotective effects of quercetin in diabetic rat retina. Saudi J. Biol. Sci. 2017, 24, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Obatomi, D.K.; Bikomo, E.O.; Temple, V.J. Anti-diabetic properties of the African mistletoe in streptozotocin-induced diabetic rats. J. Ethnopharmacol. 1994, 43, 13–17. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Kim, Y.S.; Choi, S.U.; Ryu, S.Y. Isolation of flavonol rhamnosides from Loranthus tanakae and cytotoxic effect of them on human tumor cell lines. Arch. Pharm. Res. 2004, 27, 44–47. [Google Scholar] [CrossRef]
- Adesina, S.K.; Illoh, H.C.; Johnny, I.I.; Jacobs, I.E. African mistletoes (Loranthaceae); ethnopharmacology, chemistry and medicinal values: An update. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Mothana, R.A.; Kriegisch, S.; Harms, M.; Wende, K.; Lindequist, U. Assessment of selected Yemeni medicinal plants for their in vitro antimicrobial, anticancer, and antioxidant activities. Pharm. Biol. 2011, 49, 200–210. [Google Scholar] [CrossRef]
- Mothana, R.A.; Al-Said, M.S.; Al-Rehaily, A.J.; Thabet, T.M.; Awad, N.A.; Lalk, M.; Lindequist, U. Anti-inflammatory, antinociceptive, antipyretic and antioxidant activities and phenolic constituents from Loranthus regularis Steud. ex Sprague. Food Chem. 2012, 130, 344–349. [Google Scholar] [CrossRef]
- Alanazi, A.Z.; Mohany, M.; Alasmari, F.; Mothana, R.A.A.; Alshehri, A.O.A.; Alhazzani, K.; Ahmed, M.M.; Al-Rejaie, S.S. Amelioration of Diabetes-Induced Nephropathy by Loranthus regularis: Implication of Oxidative Stress, Inflammation and Hyperlipidaemia. Appl. Sci. 2021, 11, 4548. [Google Scholar] [CrossRef]
- Alanazi, A.Z.; Mohany, M.; Alasmari, F.; Mothana, R.A.A.; Alshehri, A.O.A.; Alhazzani, K.; Ahmed, M.M.; Al-Rejaie, S.S. Protective Role of Loranthus regularis against Liver Dysfunction, Inflammation, and Oxidative Stress in Streptozotocin Diabetic Rat Model. Evid.-Based Complement. Altern. Med. 2020, 2020, 5027986. [Google Scholar] [CrossRef]
- Mothana, R.A.; Al-Musayeib, N.M.; Al-Ajmi, M.F.; Cos, P.; Maes, L. Evaluation of the in vitro antiplasmodial, antileishmanial, and antitrypanosomal activity of medicinal plants used in saudi and yemeni traditional medicine. Evid. Based-Complement. Altern. Med. 2014, 2014, 905639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navaratna, D.; Guo, S.Z.; Hayakawa, K.; Wang, X.; Gerhardinger, C.; Lo, E.H. Decreased cerebrovascular brain derived neurotrophic factor-mediated neuroprotection in the diabetic brain. Diabetes 2011, 60, 1789–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, M.; Ozawa, Y.; Kurihara, T.; Kubota, S.; Yuki, K.; Noda, K.; Kobayashi, S.; Ishida, S.; Tsubota, K. Neurodegenerative influence of oxidative stress in the retina of a murine model of diabetes. Diabetologia 2010, 53, 971–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Nicoló, S.; Tarani, L.; Ceccanti, M.; Maldini, M.; Natella, F.; Vania, A.; Chaldakov, G.N.; Fiore, M. Effects of olive polyphenols administration on nerve growth factor and brain derived neurotrophic factor in the mouse brain. Nutrition 2013, 29, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Xia, X.B.; Xiong, S.Q. BDNF regulates GLAST and glutamine synthetase in mouse retinal Muller cells. J. Cell. Physiol. 2012, 227, 596–603. [Google Scholar] [CrossRef]
- Wang, Q.; Pfister, F.; Dorn-Beineke, A.; vom Hagen, F.; Lin, J.; Feng, Y.; Hammes, H.P. Low-dose erythropoietin inhibits oxidative stress and early vascular changes in the experimental diabetic retina. Diabetologia 2010, 53, 1227–1238. [Google Scholar] [CrossRef] [Green Version]
- Al-Gayyar, M.M.; Matragoon, S.; Pillai, B.A.; Ali, T.K.; Abdelsaid, M.A.; El-Remessy, A.B. Epicatechin blocks pro-nerve growth factor (proNGF)-mediated retinal neurodegeneration via inhibition of p75 neurotrophin receptor expression in a rat model of diabetes. Diabetologia 2011, 54, 669–680. [Google Scholar] [CrossRef] [Green Version]
- Silva, K.C.; Rosales, M.A.; Hamassaki, D.E.; Saito, K.C.; Faria, A.M.; Ribeiro, P.A.; Faria, J.B.; Faria, J.M. Green tea is neuroprotective in diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1325–1336. [Google Scholar] [CrossRef] [Green Version]
- Mothana, R.A.A.; Al-Said, M.S.; Al-Rehaily, A.J.; Thabet, T.M.; Awad, N.A.; Lalk, M.; Lindequist, U. Anti-inflammatory, antinociceptive, antipyretic and antioxidant activities and phenolic constituents from Loranthus regularis growing in Saudi Arabia. Planta Med. 2013, 79, P66. [Google Scholar] [CrossRef]
- Achi, N.; Ohaeri, O.C.; Ijeh, I.I.; Eleazu, C. Modulation of the lipid profile and insulin levels of streptozotocin induced diabetic rats by ethanol extract of Cnidoscolus aconitifolius leaves and some fractions: Effect on the oral glucose tolerance of normoglycemic rats. Biomed. Pharmacother. 2017, 86, 562–569. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ola, M.S.; Alanazi, A.Z.; Malik, A.; Malik, A.; Ahmed, M.; Al-Rejaie, S.S.; Alhomida, A.S. Loranthus regularis Ameliorates Neurodegenerative Factors in the Diabetic Rat Retina. Appl. Sci. 2022, 12, 2875. https://doi.org/10.3390/app12062875
Ola MS, Alanazi AZ, Malik A, Malik A, Ahmed M, Al-Rejaie SS, Alhomida AS. Loranthus regularis Ameliorates Neurodegenerative Factors in the Diabetic Rat Retina. Applied Sciences. 2022; 12(6):2875. https://doi.org/10.3390/app12062875
Chicago/Turabian StyleOla, Mohammad Shamsul, Ahmed Z. Alanazi, Ajamaluddin Malik, Abdul Malik, Mohammed Ahmed, Salim S. Al-Rejaie, and Abdullah S. Alhomida. 2022. "Loranthus regularis Ameliorates Neurodegenerative Factors in the Diabetic Rat Retina" Applied Sciences 12, no. 6: 2875. https://doi.org/10.3390/app12062875