
Mesenchymal Stromal Cells (MSCs) Isolated from Various Tissues of the Human Arthritic Knee Joint Possess Similar Multipotent Differentiation Potential

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Cultivation of Cells
2.2. Flow Cytometry Analysis of Surface Antigens
2.3. Cell Culture and Differentiation
2.4. Histological Staining and Immunohistochemistry
2.5. RNA Isolation and Semi-Quantitative RT-PCR
2.6. Statistical Analysis
3. Results
3.1. Isolation of Cells from Respective Native Tissues
3.2. Surface Markers of Isolated MSCs
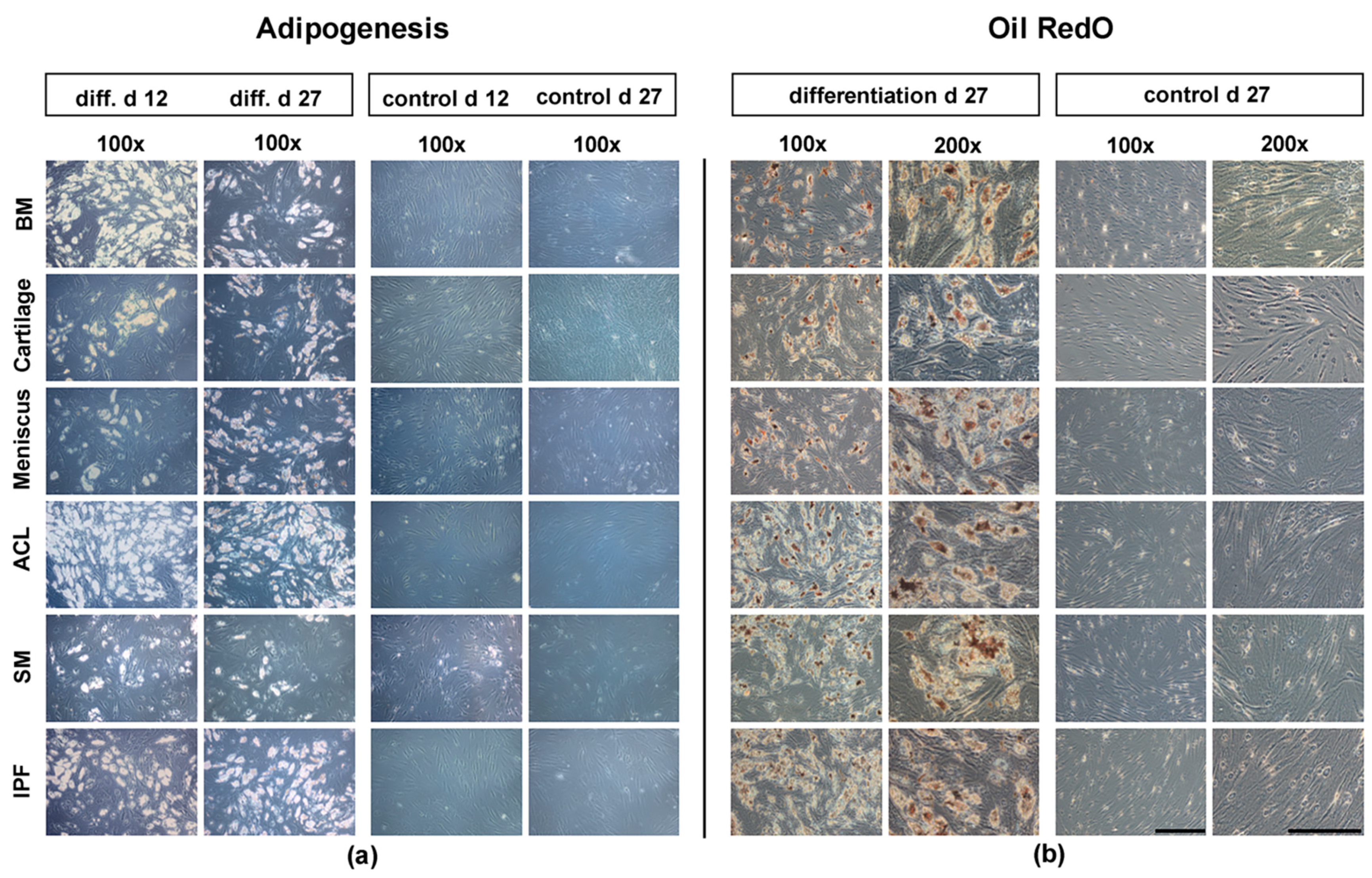
3.3. Histological Analysis of Adipogenesis
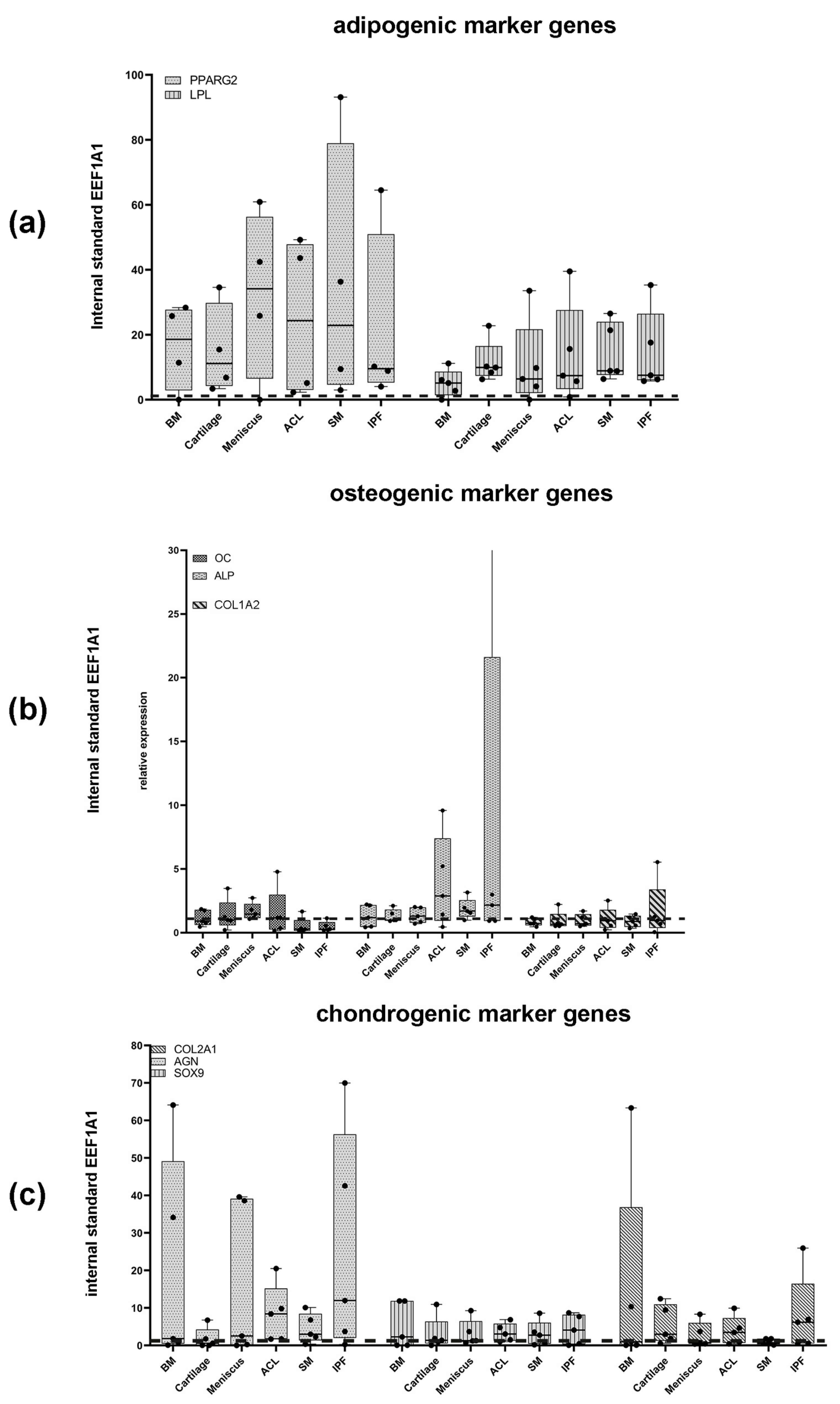
3.4. Expression of Adipogenic Marker Genes after 27 d
3.5. Histological and Immunohistochemical Analysis of Osteogenesis
3.6. Expression of Osteogenic Marker Genes after 27 d
3.7. Histological and Immunohistochemical Analysis of Chondrogenesis
3.8. Expression of Chondrogenic Marker Genes after 21 d
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sharma, P.; Kumar, P.; Sharma, R.; Bhatt, V.D.; Dhot, P.S. Tissue Engineering; Current Status & Futuristic Scope. J. Med. Life 2019, 12, 225–229. [Google Scholar] [PubMed]
- Bauge, C.; Boumediene, K. Use of Adult Stem Cells for Cartilage Tissue Engineering: Current Status and Future Developments. Stem Cells Int. 2015, 2015, 438026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzobo, K.; Thomford, N.; Senthebane, D.; Shipanga, H.; Rowe, A.; Dandara, C.; Pillay, M.; Motaung, K.S. Advances in Regenerative Medicine and Tissue Engineering: Innovation and Transformation of Medicine. Stem Cells Int. 2018, 2018, 2495848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jessop, Z.; Al-Sabah, A.; Francis, W.R.; Whitaker, I.S. Transforming healthcare through regenerative medicine. BMC Med. 2016, 14, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berebichez-Fridman, R.; Montero-Olvera, P.R. Sources and Clinical Applications of Mesenchymal Stem Cells: State-of-the-art review. Sultan Qaboos Univ. Med. J. 2018, 18, e264–e277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, M.J. Keeping kidneys. Bull. World Health Organ. 2012, 90, 718–719. [Google Scholar]
- Żylińska, B.; Silmanowicz, P.; Sobczyńska-Rak, A.; Jarosz, Ł.; Szponder, T. Treatment of Articular Cartilage Defects: Focus on Tissue Engineering. In Vivo 2018, 32, 1289–1300. [Google Scholar] [CrossRef] [Green Version]
- Health at a Glance 2019 OECD Indicators: OECD Indicators; OECD Publishing: Berlin, Germany, 2019.
- Klug, A.; Gramlich, Y.; Rudert, M.; Drees, P.; Hoffmann, R.; Weißenberger, M.; Kutzner, K.P. The projected volume of primary and revision total knee arthroplasty will place an immense burden on future health care systems over the next 30 years. Knee Surg. Sports Traumatol. Arthrosc. 2020, 12, 3287–3298. [Google Scholar] [CrossRef]
- Karachalios, T.; Komnos, G.; Koutalos, A. Total hip arthroplasty: Survival and modes of failure. EFORT Open Rev. 2018, 3, 232–239. [Google Scholar] [CrossRef]
- Argenson, J.N.; Boisgard, S.; Parratte, S.; Descamps, S.; Bercovy, M.; Bonnevialle, P.; Briard, J.L.; Brilhault, J.; Chouteau, J.; Nizard, R.; et al. Survival analysis of total knee arthroplasty at a minimum 10 years’ follow-up: A multicenter French nationwide study including 846 cases. Orthop. Traumatol. Surg. Res. 2013, 99, 385–390. [Google Scholar] [CrossRef] [Green Version]
- Zhan, C.; Kaczmarek, R.; Loyo-Berrios, N.; Sangl, J.; Bright, R.A. Incidence and short-term outcomes of primary and revision hip replacement in the United States. J. Bone Joint Surg. Am. 2007, 89, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Lee, S.H.; Song, E.K.; Seon, J.K.; Lim, H.A.; Yang, H.Y. Causes and Clinical Outcomes of Revision Total Knee Arthroplasty. Knee Surg. Relat. Res. 2017, 29, 104–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, G.I. Tissue Engineering in Osteoarthritis: Current Status and Prospect of Mesenchymal Stem Cell Therapy. BioDrugs 2018, 32, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, E.M.; Blanc, K.L.; Dominici, M.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Deans, R.J.; Krause, D.S.; Keating, A. Clarification of the nomenclature for MSC: The International Society for Cellular Therapy position statement. Cytotherapy 2005, 7, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Li, X.; Zhang, Y.; Han, Y.; Chang, F.; Ding, J. Mesenchymal Stem Cells for Regenerative Medicine. Cells 2019, 8, 886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinert, A.F.; Ghivizzani, S.C.; Rethwilm, A.; Tuan, R.S.; Evans, C.H.; Nöth, U. Major biological obstacles for persistent cell-based regeneration of articular cartilage. Arthritis Res. Ther. 2007, 9, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, Y.; Sekiya, I.; Yagishita, K.; Muneta, T. Comparison of human stem cells derived from various mesenchymal tissues: Superiority of synovium as a cell source. Arthritis Rheum. 2005, 52, 2521–2529. [Google Scholar] [CrossRef] [PubMed]
- Wagenbrenner, M.; Heinz, T.; Horas, K.; Jakuscheit, A.; Arnholdt, J.; Herrmann, M.; Rudert, M.; Holzapfel, B.M.; Steinert, A.F.; Weißenberger, M. The human arthritic hip joint is a source of mesenchymal stromal cells (MSCs) with extensive multipotent differentiation potential. BMC Musculoskelet Disord. 2020, 21, 297. [Google Scholar] [CrossRef]
- Elahi, K.C.; Klein, G.; Avci-Adali, M.; Sievert, K.D.; MacNeil, S.; Aicher, W.K. Human Mesenchymal Stromal Cells from Different Sources Diverge in Their Expression of Cell Surface Proteins and Display Distinct Differentiation Patterns. Stem Cells Int. 2016, 2016, 5646384. [Google Scholar] [CrossRef] [Green Version]
- Fu, W.; Xie, X.; Li, Q.; Chen, G.; Zhang, C.; Tang, X.; Li, J. Isolation, Characterization, and Multipotent Differentiation of Mesenchymal Stem Cells Derived from Meniscal Debris. Stem Cells Int. 2016, 2016, 5093725. [Google Scholar] [CrossRef] [PubMed]
- Steinert, A.F.; Kunz, M.; Prager, P.; Barthel, T.; Jakob, F.; Nöth, U.; Murray, M.M.; Evans, C.H.; Porter, R.M. Mesenchymal stem cell characteristics of human anterior cruciate ligament outgrowth cells. Tissue Eng. Part A 2011, 17, 1375–1388. [Google Scholar] [CrossRef] [PubMed]
- Steinert, A.F.; Proffen, B.; Kunz, M.; Hendrich, C.; Ghivizzani, S.C.; Nöth, U.; Rethwilm, A.; Eulert, J.; Evans, C.H. Hypertrophy is induced during the in vitro chondrogenic differentiation of human mesenchymal stem cells by bone morphogenetic protein-2 and bone morphogenetic protein-4 gene transfer. Arthritis Res. Ther. 2009, 11, R148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichert, J.C.; Schmalzl, J.; Prager, P.; Gilbert, F.; McQuent, V.; Steinert, A.F.; Rudert, M.; Noth, U. Synergistic effect of Indian hedgehog and bone morphogenetic protein-2 gene transfer to increase the osteogenic potential of human mesenchymal stem cells. Stem Cell Res. Ther. 2013, 4, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nöth, U.; Osyczka, A.M.; Tuli, R.; Hickok, N.J.; Danielson, K.G.; Tuan, R.S. Multilineage mesenchymal differentiation potential of human trabecular bone-derived cells. J. Orthop. Res. 2002, 20, 1060–1069. [Google Scholar] [CrossRef]
- Steinert, A.F.K.M.; Prager, P.; Gobel, S.; Klein-Hitpass, L.; Ebert, R.; Noth, U.; Jakob, F.; Gohlke, F. Characterization of bursa subacromialis-derived mesenchymal stem cells. Stem Cell Res. Ther. 2015, 6, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zołocińska, A. The expression of marker genes during the differentiation of mesenchymal stromal cells. Adv. Clin. Exp. Med. 2018, 27, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.B.; Ha, C.W.; Rhim, J.H.; Lee, H.J. Stem Cell Therapy for Articular Cartilage Repair: Review of the Entity of Cell Populations Used and the Result of the Clinical Application of Each Entity. Am. J. Sports Med. 2018, 46, 2540–2552. [Google Scholar] [CrossRef]
- Berebichez-Fridman, R.; Gómez-García, R.; Granados-Montiel, J.; Berebichez-Fastlicht, E.; Olivos-Meza, A.; Granados, J.; Velasquillo, C.; Ibarra, C. The Holy Grail of Orthopedic Surgery: Mesenchymal Stem Cells-Their Current Uses and Potential Applications. Stem Cells Int. 2017, 2017, 2638305. [Google Scholar] [CrossRef] [PubMed]
- Mafi, R.; Hindocha, S.; Mafi, P.; Griffin, M.; Khan, W.S. Sources of adult mesenchymal stem cells applicable for musculoskeletal applications—A systematic review of the literature. Open Orthop. J. 2011, 5 (Suppl. 2), 242–248. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Charbord, P. Bone marrow mesenchymal stem cells: Historical overview and concepts. Hum. Gene. Ther. 2010, 21, 1045–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pretzel, D.; Linss, S.; Rochler, S.; Endres, M.; Kaps, C.; Alsalameh, S.; Kinne, R.W. Relative percentage and zonal distribution of mesenchymal progenitor cells in human osteoarthritic and normal cartilage. Arthritis Res. Ther. 2011, 13, R64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsalameh, S.; Amin, R.; Gemba, T.; Lotz, M. Identification of mesenchymal progenitor cells in normal and osteoarthritic human articular cartilage. Arthritis Rheum. 2004, 50, 1522–1532. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, A.; Uchida, S.; Utsunomiya, H.; Tsukamoto, M.; Nakashima, H.; Nakamura, E.; Pascual-Garrido, C.; Sekiya, I.; Sakai, A. Isolation and Characterization of Synovial Mesenchymal Stem Cell Derived from Hip Joints: A Comparative Analysis with a Matched Control Knee Group. Stem Cells Int. 2017, 2017, 9312329. [Google Scholar] [CrossRef] [PubMed]
- De Bari, C.; Dell’Accio, F.; Tylzanowski, P.; Luyten, F.P. Multipotent mesenchymal stem cells from adult human synovial membrane. Arthritis Rheum. 2001, 44, 1928–1942. [Google Scholar] [CrossRef]
- Li, N.; Gao, J.; Mi, L.; Zhang, G.; Zhang, L.; Zhang, N.; Huo, R.; Hu, J.; Xu, K. Synovial membrane mesenchymal stem cells: Past life, current situation, and application in bone and joint diseases. Stem Cell Res. Ther. 2020, 11, 381. [Google Scholar] [CrossRef] [PubMed]
- Macchi, V.; Stocco, E.; Stecco, C.; Belluzzi, E.; Favero, M.; Porzionato, A.; De Caro, R. The infrapatellar fat pad and the synovial membrane: An anatomo-functional unit. J. Anat. 2018, 233, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Huri, P.Y.; Hamsici, S.; Ergene, E.; Huri, G.; Doral, M.N. Infrapatellar Fat Pad-Derived Stem Cell-Based Regenerative Strategies in Orthopedic Surgery. Knee Surg. Relat. Res. 2018, 30, 179–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.F.; Chen, Y.T.; Yang, T.H.; Chen, L.L.; Chiou, S.H.; Tsai, T.H.; Tsai, C.C.; Chen, M.H.; Ma, H.L.; Hung, S.C. Isolation and characterization of mesenchymal stromal cells from human anterior cruciate ligament. Cytotherapy 2008, 10, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.T.; Yang, H.W.; Chen, T.H.; Lee, O.K. Isolation and characterization of multipotent stem cells from human cruciate ligaments. Cell Prolif. 2009, 42, 448–460. [Google Scholar] [CrossRef] [PubMed]
- Sheng, G. The developmental basis of mesenchymal stem/stromal cells (MSCs). BMC Dev. Biol. 2015, 15, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segawa, Y.; Muneta, T.; Makino, H.; Nimura, A.; Mochizuki, T.; Ju, Y.J.; Ezura, Y.; Umezawa, A.; Sekiya, I. Mesenchymal stem cells derived from synovium, meniscus, anterior cruciate ligament, and articular chondrocytes share similar gene expression profiles. J. Orthop. Res. 2009, 27, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.; Wright, K.; Roberts, S.; Kuiper, J.H.; Mangham, C.; Richardson, J.; Mennan, C. Characterisation of synovial fluid and infrapatellar fat pad derived mesenchymal stromal cells: The influence of tissue source and inflammatory stimulus. Sci. Rep. 2016, 6, 24295. [Google Scholar] [CrossRef] [Green Version]
- Hudetz, D.; Borić, I.; Rod, E.; Jeleč, Ž.; Kunovac, B.; Polašek, O.; Vrdoljak, T.; Plečko, M.; Skelin, A.; Polančec, D.; et al. Early results of intra-articular micro-fragmented lipoaspirate treatment in patients with late stages knee osteoarthritis: A prospective study. Croat. Med. J. 2019, 60, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, F.; Maglio, M.; Tschon, M.; Aldini, N.N.; Fini, M. Adipose-derived mesenchymal stem cells for cartilage tissue engineering: State-of-the-art in in vivo studies. J. Biomed. Mater. Res. A 2014, 102, 2448–2466. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Csaki, C.; Clutterbuck, A.L.; Rahmanzadeh, M.; Shakibaei, M. Mesenchymal stem cells in connective tissue engineering and regenerative medicine: Applications in cartilage repair and osteoarthritis therapy. Histol. Histopathol. 2009, 24, 347–366. [Google Scholar] [PubMed]
- Yang, Y.K. Aging of mesenchymal stem cells: Implication in regenerative medicine. Regen. Ther. 2018, 9, 120–122. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Number | Age (Years) | BMI (kg/m2) | Secondary Diagnosis | Osteoarthritis Grade (Kellgren & Lawrence) | Smoking |
|---|---|---|---|---|---|
| 1 | 61 | 23.62 | hypothyreodism | 3 | no |
| 2 | 55 | 29.07 | hypertension, breast cancer | 4 | no |
| 3 | 61 | 18.22 | - | 3 | yes |
| 4 | 62 | 28.05 | - | 3 | no |
| 5 | 59 | 26.44 | hypertension | 4 | yes |
| Nomenclature | Donor Tissue | Surgical Extraction |
|---|---|---|
| BM-MSCs | Bone marrow (BM) of the proximal tibia and the femoral condyles | The femoral condyles and the proximal tibia are removed with a bone saw during surgery to replace the damaged knee joint with the endoprosthesis. Bone marrow was extracted from the bone using a bone curette. |
| MSC-like cells | Arthritic hyaline cartilage of the tibia and the femoral condyles | Macroscopically eroded hyaline cartilage was scraped off the femoral condyles with a scalpel. |
| Inner and outer meniscus | The inner and outer meniscus are removed during the procedure and secured. | |
| Anterior cruciate ligament (ACL) | The ACL was removed during the procedure and secured. | |
| Synovial membrane (SM) | To access the knee-joint the synovial membrane was removed, and a sample was secured. | |
| Infrapatellar fat pad (IPF) | The infrapatellar fat pad is removed during surgery and was secured. |
| Gene | Primer Sequence (5′-3′) | Anneal. Temp. (°C) | Product Size (bp) | Cycles |
|---|---|---|---|---|
| Markers of adipogenesis | ||||
| LPL | Sense: GAGATTTCTCTGTATGGCACC Antisense: CTGCAAATGAGACACTTTCTC | 51.0 | 250 | 30 |
| PPARG2 | Sense: GCTGTTATGGGTGAAACTCTG Antisense: ATAAGGTGGAGATGCAGGCTC | 61.0 | 380 | 35 |
| Markers of chondrogenesis | ||||
| COL2A1 | Sense: TTTCCCAGGTCAAGATGGTC Antisense: CTTCAGCACCTGTCCACCA | 58.0 | 374 | 35 |
| AGN | Sense: TGAGGAGGGCTGGAACAAGTACC Antisense: GGAGGTGGTAATTGCAGGGAACA | 54.0 | 392 | 30 |
| SOX9 | Sense: ATCTGAAGAAGGAGAGCGAG Antisense: TCAGAAGTCTCCAGAGCTTG | 58.0 | 263 | 35 |
| Markers of osteogenesis | ||||
| COL1A2 | Sense: GGACACAATGGATTGCAAGG Antisense: TAACCACTGCTCCACTCTGG | 54.0 | 461 | 32 |
| OC | Sense: ATGAGAGCCCTCACACTCCTC Antisense: GCCGTAGAAGCGCCGATAGGC | 59.0 | 387 | 35 |
| ALP | Sense: TGGAGCTTCAGAAGCTCAACACCA Antisense: TCTCGTTGTCTGAGTACCAGTCC | 51.0 | 454 | 35 |
| Internal control | ||||
| EEF1A1 | Sense: AGGTGATTATCCTGAACCATCC Antisense: AAAGGTGGATAGTCTGAGAAGC | 54.0 | 234 | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagenbrenner, M.; Poker, K.; Heinz, T.; Herrmann, M.; Horas, K.; Ebert, R.; Mayer-Wagner, S.; Holzapfel, B.M.; Rudert, M.; Steinert, A.F.; et al. Mesenchymal Stromal Cells (MSCs) Isolated from Various Tissues of the Human Arthritic Knee Joint Possess Similar Multipotent Differentiation Potential. Appl. Sci. 2022, 12, 2239. https://doi.org/10.3390/app12042239
Wagenbrenner M, Poker K, Heinz T, Herrmann M, Horas K, Ebert R, Mayer-Wagner S, Holzapfel BM, Rudert M, Steinert AF, et al. Mesenchymal Stromal Cells (MSCs) Isolated from Various Tissues of the Human Arthritic Knee Joint Possess Similar Multipotent Differentiation Potential. Applied Sciences. 2022; 12(4):2239. https://doi.org/10.3390/app12042239
Chicago/Turabian StyleWagenbrenner, Mike, Konrad Poker, Tizian Heinz, Marietta Herrmann, Konstantin Horas, Regina Ebert, Susanne Mayer-Wagner, Boris M. Holzapfel, Maximilian Rudert, Andre F. Steinert, and et al. 2022. "Mesenchymal Stromal Cells (MSCs) Isolated from Various Tissues of the Human Arthritic Knee Joint Possess Similar Multipotent Differentiation Potential" Applied Sciences 12, no. 4: 2239. https://doi.org/10.3390/app12042239